Аллельный полиморфизм гена PIT-1 в стадах крупного рогатого скота Брянской области и его связь с молочной продуктивностью

Автор: Дроздов Евгений Владимирович, Заякин Владимир Васильевич, Нам Ирина Янгуковна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Генетика
Статья в выпуске: 5-3 т.13, 2011 года.
Бесплатный доступ
Изучен аллельный полиморфизм гипофизарного фактора транскрипции PIT-1 в трех группах КРС. Более редкий А-аллель обеспечивает достоверное повышение молочной продуктивности у коров черно-пестрой породы.
Гипофизарный фактор транскрипции, аллельный полиморфизм, молочная продуктивность
Короткий адрес: https://sciup.org/148200455
IDR: 148200455 | УДК: 575.113.2+575.17:577.213/217:636.2
Allelic polymorphism of factor PIT-1 in cattle of Bryansk region and its relation with dairy productivity
Allelic polymorphism of factor pit-1 has been investigated in tree cattle groups. Rare A-allele give essential rise of dairy productivity.
Текст научной статьи Аллельный полиморфизм гена PIT-1 в стадах крупного рогатого скота Брянской области и его связь с молочной продуктивностью
Возрождение животноводства, в частности молочного, является одной из важнейших задач сельского хозяйства России. Большую роль при этом играет целенаправленное формирование стад с высоким генетическим потенциалом молочной продуктивности. Первоначально для этих целей использовали морфологические признаки. Развитие молекулярной биологии и биотехнологии обеспечило создание новых тест-систем, позволяющих выявлять генетические маркеры непосредственно на уровне ДНК.
Молочная продуктивность относится к сложным количественным признакам, которые контролируются большим количеством генетических локусов (QTL). К таким локусам относятся гены белков молока, вторую группу составляют гены гормонов, в частности, пролактина и соматотропина, определяющие развитие животных, подготовку к лактации и стимулирующие саму лактацию. Для этих генов показана возможная связь с молочной продуктивностью и качеством молока. Мы изучили аллельный полиморфизм генов каппа-казеина и бета-лактоглобулина, а также пролактина и соматотропина, в некоторых стадах КРС Брянской обл. и подтвердили наличие корреляций молочной продуктивности с определенными аллелями генов лактоглобулина и этих гормонов [1,2].
Гипофизарный фактор транскрипции (PIT-1) занимает особое место в детерминации молочной продуктивности и может рассматриваться как третья, самая высокая ступень в регуляции этого процесса. На ранних этапах эмбриогенеза он направляет дифференциацию клеток гипофиза, определяет развитие зон ответственных за синтез соматотропина, пролактина и тиреотропного гормона и участвует в регуляции экспрессии их генов [3]. Его аллельные варианты могут влиять на молочную продуктивность [4,5], мутации в гене фактора PIT-1 приводят к гипоплазии гипофиза, синдрому врожденного гипотиреоза у мышей, а
также к отсутствие соматотропина [6]. Ген фактора PIT-1 КРС расположен в центромерной зоне первой хромосомы между локусами TGLA57 и RM95 [7].
Белок PIT-1 состоит из 291 аминокислоты, узнает мотив ATGNATA(A/T)(A/T) и является членом POU-домена, в который входит группа транскрипционных регуляторов, имеющих важную роль в дифференциации и пролиферации клеток. POU является бинарным ДНК-связывающим доменом. Он включает два высоко консервативных региона. Эти регионы ответственны за высокое сродство к ДНК генов соматотропина, пролактина и др. [8].
У млекопитающих Pit-1 имеет три варианта сплайсинга. Основной тип, Pit1-альфа, два других – Pit1-бета и Pit-1T. Все варианты сплайсинга дают биологически активные продукты. Каждый из вариантов избирательно воздействует на промоторные зоны генов мишеней [9].
В настоящее время выявлено несколько вариантов гена Pit-1, определяемых точечными мутациями. Три мутации - Pit1I3H (C- и D–аллели), Pit1I3N (M- и N–аллели) и Pit1I3NL (G- и H– аллели) локализованы в третьем интроне. Эти мутации выявляются с помощью рестриктаз HinfI, NciI и NlaIII, соответственно. Также по одной мутации Pit1I4N (E- и F–аллели), Pit1I5, Pit1E6H (A- и B–аллели) обнаружено в четвертом, пятом интронах и шестом экзоне гена Pit-1. Нуклеотидные замены в четвертом интроне и шестом экзоне идентифицируются эндонуклеазами BstNI и HinfI, соответственно. Аллель B определяет точечная мутация, приводящая к замене аденина на гуанин (А→G) [10,11].
Полиморфизм Pit1 может быть информативным маркером молочной и мясной продуктивности. Однако этот вопрос изучен недостаточно. Обнаружена некоторая породоспецифичность действия генов, их вклада в количественные полигенные признаки. Существуют также противоречия в литературных данных. Так, А-аллель связан с преимуществом по удою и более высоким качеством молока [12]. В то же время одни исследователи обнаруживают, что В-аллель может увеличивать морфометрические показатели молодых животных, а другие вообще не выявили влияния аллелей Pit-1 на рост животных [10,13].
Целью работы является изучение аллельного полиморфизма фактора PIT-1 в некоторых стадах крупного рогатого скота (КРС) Брянской обл. и проверка связи аллелей его гена с молочной продуктивностью.
МАТЕРИАЛ И МЕТОДЫ
Работа выполнялась на случайных выборках животных: 55 коров и 16 телят черно-пестрой породы в ОАО «Снежка-Госома»; 50 коров айрширской породы СПХ «Сельцо»; 146 частных беспородных коров на территории трех деревень (Колягино, Норино и Морочево) Жирятинского р-на Брянской обл. Кровь у животных отбирали из яремной вены. В качестве антикоагулянта использовали 0,5М раствор ЭДТА до конечной концентрации 10 мМ.
Исследования влияния генотипов на молочную продуктивность проводились в группе крупного рогатого скота ОАО «Снежка-Госома». Животные в течение проведения опыта были здоровыми и находились в одинаковых условиях кормления и содержания. По данным зоотехнического учета анализировали показатели удоя и содержание жира в молоке коров опытной группы.
Праймеры для проведения ПЦР синтезировали в ЗАО «Синтол», Россия. Их состав соответствовал составу праймеров в описанной в литературе методике [7]: PIT-1 /HinfI прямой: 5/- CAA-TGA-GAA-AGT-TGG-TGC-3/; PIT-1 /HinfI обратный: 5/- TCT-GCA-TTC-GAG-ATG-CTC-3.
В состав ПЦР смеси кроме праймеров и ДНК входили 200 мкМ каждого нуклеозидтрифосфата; 2,0 единицы Taq-полимеразы и 2,0 мкл 10-кратного буфера для полимеразы. Состав буфера: 700 mM Трис-HCl, pH 8.6/25°C, 166 mM (NH 4 ) 2 SO 4 , 25 mM MgCl 2 . Оптимальное количество каждого
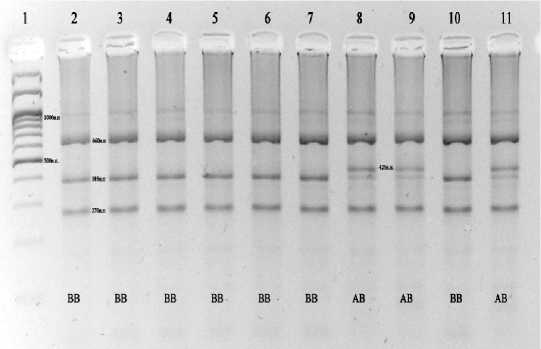
Рис. 1 . Электрофоретический анализ продуктов рестрикции, при определении генотипов PIT-I. 1 – маркер
молекулярных масс М27 (СибЭнзим); генотипы образцов 2-11 указаны на фотографии.
При этом для генотипа АА характерны фрагменты – 660, 425, 270 н.п.; для ВВ – 660, 385 и 270 н.п.; для АВ – 660, 425, 385 и 270 н.п.
В приведенном на рисунке примере электрофореграммы видно, что в проанализи- специфического праймера составляло 25-30 пмолей на пробу.
Реакцию проводили в амплификаторе «Терцик» фирмы «ДНК-технология». ДНК денатурировали при 94°С в течение 4 мин, далее проводили 35 циклов амплификации в следующем режиме: 94°С – 1 мин, отжиг праймеров при 54°С – 1 мин, элонгация при 72°С – 1 мин. Конечный этап синтеза проводили при 72°С в течение 4 мин.
Для определения полиморфизма гена Pit1 3 мкл ПЦР смеси обрабатывали 5 ед. эндонуклы рестрикции HinfI в 1×буфере «O» фирмы СибЭнзим (Россия) при 37 0С течение 1 ч.
Появление фрагментов – 660 н.п., 425 н.п. и 270 н.п. после обработки продуктов ПЦР рестриктазой соответствует (А) - аллелю гена; фрагментов - 660 н.п., 385 н.п. и 270 н.п. указывает на (В)-вариант.
Для разделения фрагментов ДНК проводили горизонтальный электрофорез при 10 В/см в 1×ТВЕ буфере на 1,0 % агарозной пластине содержащей 0,1 мкг/мл этидия бромида. В качестве маркера молекулярных масс использовали стандартный набор М27 (СибЭнзим).
Фиксацию и обработку результатов ПЦР осуществляли с помощью системы GelDoc фирмы BioRad.
Статистическую обработку и оформление результатов осуществляли с помощью программ «Statistika 6.0.», «Popgene 1.31.», MS Excel 2003 (Microsoft) и «Pop Stat».
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
При проведении ПЦР в указанных выше условиях амплифицировался фрагмент ДНК длиной 1355 п.о, что соответствует литературным данным. После его расщепления эндонуклеазой HinfI на электрофореграммах появлялись специфические наборы полос (рис. 1).
рованных образцах значительно преобладает по встречаемости В-аллель. Это соответствует результатам генетического анализа изучаемых групп КРС (табл. 1).
Таблица 1. Генетическая структура анализируемых групп КРС по гену Pit-1
|
Группы животных |
n |
Частота встречаемости генотипов |
2* χ2 |
Частота встречаемости аллелей |
Hobs |
Hex |
|||
|
АА |
АВ |
ВВ |
А |
В |
|||||
|
Коровы черно-пестрой породы |
55 |
0,06 |
0,12 |
0,81 |
6,75 |
0,11 |
0,88 |
0,13 |
0,23 |
|
Телята черно-пестрой породы |
16 |
0 |
0 |
1 |
18,53 |
0 |
1 |
0 |
0,50 |
|
Коровы айрширской породы |
50 |
0,11 |
0,23 |
0,66 |
6,33 |
0,33 |
0,77 |
0,23 |
0,35 |
|
Частный скот |
146 |
0,04 |
0,25 |
0,71 |
1,23 |
0,16 |
0,83 |
0,25 |
0,28 |
Максимальное преобладание частоты генотипа ВВ наблюдается у животных черно-пестрой породы: в группе коров (0,81), а у телят чернопестрой породы это единственный встреченный генотип. Наибольшая частота встречаемости А-аллеля у коров Айширской породы.
В трех анализируемых группах, кроме группы частного скота, наблюдается смещение генетического равновесия. В этих группах наблюдаемая гетерозиготность достоверно ниже теоретически рассчитанной.
Из гистограммы видно, что по гену Pit-1 во всех группах скота генетическая структура практически одинаковая – наблюдается преимущественная гомозиготизация в сторону В-аллеля (рис. 2). Наиболее сильно это выражено в стаде КРС ОАО "Снежка-Госома". Самый высокий уровень гетерозигот зафиксирован в группе частного скота (25%), но в этой, же группе самая низкая частота животных с АА-генотипом (4%).
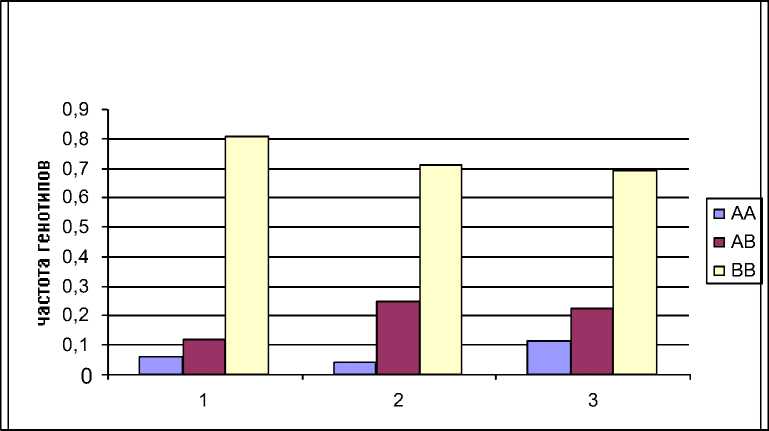
Рис. 2. Соотношение частот генотипов Pit-I в анализируемых группах КРС. 1 – группа коров черно-пестрой породы, ОАО «Снежка-Госома; 2 – частный скот (Жирятинский р-н Брянской обл.); 3 – группа коров айрширской породы, СПХ Сельцо.
В таблице 2 приведены данные о частоте В-аллеля у различных пород КРС из разных регионов мира. В европейских и северо-американских породах наблюдается преобладание В-аллеля, в большинстве случаев очень значительное. При анализе таблицы 2 видно, что из всех групп особо выделяется КРС Ирана. Там наблюдается обратная тенденция – преобладание частоты А-аллеля. Также из таблицы 2 видно, что частота аллелей Pit-1 не является строгой породоспецифичной характеристикой. Так, например, в группах коров черно-пестрой породы частота В-аллеля находится в диапазоне от 0,640 (Беларусь) до 0,880 (Россия, Брянск).
Результаты анализа связи молочной продуктивности с аллельными вариантами Pit-1 показаны в таблице 3 и на рис. 3. Согласно таблице 3, особи с генотипом АА гена Pit-1 по среднему надою за месяц достоверно превосходят животных с другими генотипами.
Таблица 2. Частота В-аллеля Pit-1 в группах КРС Брянской обл. и в других регионах
|
Порода |
Частота В-аллеля |
Источник данных |
Страна |
|
Mazandarani, |
0,173 |
Javanmard A. et al ., 2005 |
Иран |
|
Taleshi (Bos indicus) |
0,228 |
Javanmard A. et al. , 2005 |
Иран |
|
Golpayegani |
0,254 |
Javanmard A. et al ., 2005 |
Иран |
|
Sarabi |
0,378 |
Javanmard A. et al. , 2005 |
Иран |
|
Nanyang |
0,535 |
Kai Xue et al. , 2006 |
Китай |
|
Бурая карпатская |
0,540 |
Копылова К.В., 2006 |
Украина |
|
Черно-пестрая |
0,640 |
Михайлова М.Е. и др., 2008 |
Беларусь |
|
Ангусская |
0,67 |
Zhao Q. et al ., 2010 |
США |
|
Гоштино-фризская |
0,725 |
Vargas L.D. et al ., 2004 |
Чили |
|
Голштинская |
0,744 |
Edriss et al ., 2008 |
Иран |
|
Черно-пестрая |
0,750 |
Oprz^dek J. et al., 2003 |
Польша |
|
Liupan |
0,757 |
Ju-cai L. et al ., 2009 |
Китай |
|
Черно-пестрая |
0,757 |
Dybus A. et al ., 2004 |
Польша |
|
Айрширская |
0,770 |
Собственные данные |
Россия |
|
Romanian Simmental breed |
0,780 |
Viorica C. et al. , 2008 |
Румыния |
|
Голштино-фризская |
0,812 |
Renaville R. et al., 1997 |
Италия |
|
Частный скот |
0,830 |
Собственные данные |
Россия |
|
Голштинская |
0,845 |
Hori-Oshima S. et al ., 2003 |
США |
|
Черно-пестрая |
0,849 |
Малиенко В.А.,и др., 2008 |
Украина |
|
Польская красная |
0,863 |
Zatoń-Dobrowolska M. et al ., 2007 |
Польша |
|
Черно-пестрая |
0,880 |
Собственные данные |
Россия |
|
Немецкая красная |
0,921 |
Zatoń-Dobrowolska M. et al ., 2007 |
Польша |
|
Indian zebuine cattle |
0,937 |
Mukesha M. et al ., 2007 |
Индия |
|
Чешская красная |
0,951 |
Zatoń-Dobrowolska M. et al. , 2007 |
Польша |
Относительно показателей процентного содержание жира в молоке не было выявлено статистически достоверных различий между особями с разными генотипами.
Таблица 3. Молочная продуктивность коров чёрнопёстрой породы с различными генотипами Pit-1
|
Геноти п |
Средний надой за мес |
Контр. дойка, кг |
Жирность,% |
|
ВВ |
488,9±4,0 |
16,90±0,03 |
3,662±0,002 |
|
АВ |
492,6±4,3 |
13,80±1,70 |
3,663±0,002 |
|
АА |
508,9±7,8* |
17,65±0,05 |
3,645±0,017 |
Прим. *- Р-0.05
По спектру распределения животных с разными генотипами по величине среднего надоя за месяц было установлено, что в группе коров с генотипом АА присутствуют особи с продуктивностью более 550 кг молока в месяц (рис. 3).
В группах с другими генотипами таких коров не выявлено.
В группах коров с АВ и ВВ генотипами наблюдали примерно одинаковые доли коров с минимальной и средней продуктивностью.
Таким образом, аллельная структура гена Pit-1 может вносить существенный вклад в полигенный признак молочной продуктивности, по крайней мере, у черно-пестрой породы КРС.
При этом А-аллель гена, связанный с повышенной продуктивностью, может рассматриваться как относительно редкий.
В анализируемой группе телят он отсутствует, то есть существует вероятность его исчезновения в этом хозяйстве уже в ближайших поколениях КРС. Поэтому можно рекомендовать при проведении племенной работы учитывать генотипы животных по этому гену для повышения генетического потенциала молочной продуктивности стад и сохранения биоразнообразия КРС.
Работа выполнена при поддержке грантов ФЦП № 02.740.11.0285, АВЦП № 2.1.1/224 и гранта администрации Брянской области № 215.
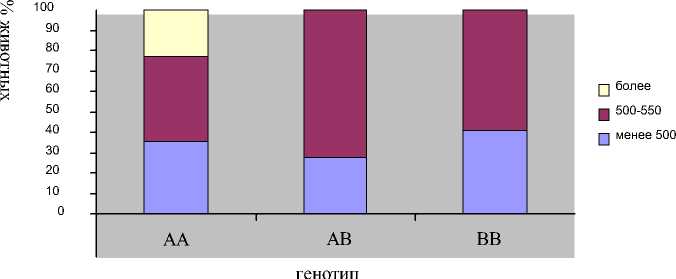
Рис. 3. Распределение коров с разными генотипами фактора PIT-1 в зависимости от уровня среднего надоя за месяц.
Список литературы Аллельный полиморфизм гена PIT-1 в стадах крупного рогатого скота Брянской области и его связь с молочной продуктивностью
- Дроздов Е.В., Заякин В.В., Нам И.Я. Анализ полиморфизма генов каппа-казеина, β-лактоглобулина, пролактина, ген рилизинг-фактора и соматотропина по AluI и MspI маркерам у коров айрширской породы//Вестник Брянского гос. ун-та. 2009. С. 119-125.
- Дроздов Е.В., Заякин В.В., Нам И.Я. Влияние аллельных вариантов гена гормона роста на параметры молочной продуктивности КРС черно-пестрой породы в Брянской области//Высокие технологии, фундаментальные и прикладные исследования в медицине и биологии: Сб. трудов междунар. научно-практ. конф. СПб., 2010. С. 214-219.
- Anderson B., Rosenfeld M.G. Pit-1 determines cell types during development of the anterior pituitary gland//J. Biol. Chem. 1994. V. 269. P. 29335-29338.
- Renaville R., Gengler N., Vrech E. et al. Pit-1. gene polymorphism, milk yield, and conformation traits for Italian Holstein-Friesian bulls//J. Dairy Sci. 1997. V. 80. P. 3431-3438.
- Keiko Kusamura de Mattos, Silvia Nassif Del Lama, Mario Luiz Martinez, Ary Ferreira Freitas. Association of bGH and Pit-1 gene variants with milk production traits in dairy Gyr bulls//Pesq. agropec. bras. Brasilia. 2004. V. 39 (2). P. 147-150.
- Li S., Crenshaw III E.B., Rawson E.J. et al. Dwarf locus mutants lacking three pituitary cell types result from mutations in the POU-domain gene Pit-1//Nature. 1990. V. 347. P. 528-534.
- Moody D.E., Pomp D., Barendse W. Restriction fragment length polymorphism in amplification products of the bovine Pit-1 gene and assignment of Pit-1 to bovine chromosome 1//Anim. Gen. 1995. V. 26. P. 45-47.
- Klemm J.D., Pabo C.O. Oct-1 POU domain-DNA interactions: Cooperative binding of isolated subdomains and effects of covalent linkage//Genes & Dev. 1996. V. 10. P. 27-36.
- Konzak K.E., Moore D. Functional isoforms of Pit-1 generated by alternative messenger RNA splicing//Mol. Endocrinol. 1992. V. 6. P. 241-247.
- Zhao Q., Davis M.E., Hines H.C. Associations of polymorphisms in the Pit-1 gene with growth and carcass traits in Angus beef cattle//J. Anim. Sci. 2004. V. 82. P. 2229-2233.
- Woollard J., Schmitz C.B., Freeman A.E., Tuggle C.K. Communication HinfI polymorphism at the bovine PIT1 locus//J. Anim. Sci. 1994. V. 72. P. 3267.
- Zwierzchowski L., Krzyzewski J., Strzalkowska N. et al. Effects of polymorphism of growth hormone (GH), Pit-1, and leptin (LEP) genes, cow’s age, lactation stage and somatic cell count on milk yield and composition of Polish Black-and-White cows//Anim. Sci. Papers and Reports. 2002. V. 20. P. 213-227.
- Dybus A., Kmiec M., Sobek Z. et al. Associations between polymorphism of growth hormone releasing hormone (GHRH) and pituitary transcription factor 1 (PIT1) genes and production traits of Limousine cattle//Arch. Tierz., Dummerstorf. 2003. V. 46. P. 527-534.
- Javanmard A., Banabazi M.H., Tavakolian J. The allele and genotype frequencies of bovine pituitary-specific transcription factor and leptin genes in Iranian cattle and buffalo populations using PCR-RFLP//Iran. J. Biotechn. 2005. V. 3. N. 2. P. 104-108.
- Zatoń-Dobrowolska M., Čitek J., Filistowicz A. Genetic distance between the Polish Red, Czech Red and German Red cattle estimated based on selected loci of protein coding genes and DNA microsatellite sequences//Anim. Sci. Papers and Reports. 2007. V. 25. N 1. P. 45-54.
