Аминокислоты в раннепалеозойских брахиоподах Тимано-Североуральского бассейна

Автор: Шанина С.Н., Безносова Т.М.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Геолого-минералогические науки
Статья в выпуске: 2 (10), 2012 года.
Бесплатный доступ
Исследован аминокислотный состав брахиопод и вмещающих их карбонатных пород из отложений верхнего ордовика, силура и нижнего девона западного склона Северного и Приполярного Урала, гряд Чернышева и Чернова, Север- ного Тимана, а также скважин 76-Лабаганская и 2-Адакская. Содержание аминокислот широко варьирует в изученных раковинах (10-190 мкг/г рако- вины) и практически не изменяется в породах (20-60 мкг/г породы). Уста- новлено, что в раковинах палеозойских брахиопод состав и количество ами- нокислот в значительной степени зависят от первичных условий осадконако- пления, фациальной приуроченности и типа захоронения. Большое влияние оказали процессы катагенетического прогрева пород Тимано-Североураль- ского бассейна. Вторичное перераспределение материала вмещающих пород и окислительные условия в районах естественных обнажений также способст- вовали дальнейшему разрушению и изменению органического вещества иско- паемых организмов. Показано, что увеличение содержания аминокислот в раковинах приурочено к трем интервалам силурийского разреза, формирова- ние которых совпадает с тремя эвстатическими трансгрессиями.
Аминокислоты, брахиоподы, палеозой, ордовик, силур, девон
Короткий адрес: https://sciup.org/14992536
IDR: 14992536 | УДК: 577.1:564.8:551.733.1
Amino acids of the early Paleozoic brachiopods in the Timan-North Urals basin
Amino acid composition of the Upper Ordovician, Silurian and Lower Devonian brachiopods and enclosing carbonate rocks of the western slope of the Northern and Polar Urals, Chernysheva and Chernov ridges, Northern Timan, 76- Labaganskaya and 2-Adakskaya holes was studied. The content of amino acids widely varies in shells (10-190 mg/g shell) and practically no change in the rocks (20-60 mg/g rock). It was established, that composition and quantity of amino acids in the shells of the Paleozoic brachiopods depend on the primary depositional environment, facies confinement, type of burial and the catagenetic processes of heating rocks of the Timan-North Urals basin. Secondary redistribution of host rock material and oxidizing conditions in the areas of natural exposures have also contributed to further destruction and modification of the organic substance of fossil organisms. It is shown that the increase in amino acid content in the shells is timed to three intervals of the Silurian section, which formation coincides with three eustatic transgressions.
Текст научной статьи Аминокислоты в раннепалеозойских брахиоподах Тимано-Североуральского бассейна
Корреляция пограничных толщ морских карбонатов верхнего ордовика, силура и нижнего девона на европейском Северо-Востоке основана, преимущественно, на последовательности брахио-подовой и конодонтовой фауны. Региональные границы ордовика и силура, лландовери и венлока, силура и девона, рассматриваемые как важнейшие событийные уровни, связанные с вымиранием фауны, определяются эволюционными и экологическими изменениями биоты, отражающими разномасштабные события – локальные и глобальные.
Результаты геохимических исследований показали возможность успешного применения данных по изменению изотопов углерода δ13С в палеозойских карбонатах с целью межбассейновой корреляции. В настоящее время получены изотопно-углеродные маркеры для биостратиграфически датированных интервалов верхнего ордовика, нижнего силура и нижнего девона Западного Урала [1–2]. Применение нового для региона метода исследования аминокислотного состава раковин брахиопод, наряду с палеонтологическим, седиментологическим и изо- топным анализами, послужит дополнительным критерием при распознавании важных событийных уровней в раннем палеозое и может способствовать более успешному решению ряда задач, связанных с эволюцией и экологией морской биоты. Важность таких комплексных исследований особенно значима для отложений нижнего палеозоя, таксономический состав биоты в которых ограничен, по сравнению с более молодыми образова- ниями фанерозоя.
Брахиоподы – одна из самых распространенных и хорошо изученных групп фауны, содержащихся в отложениях всех горизонтов рассматриваемого стратиграфического интервала Тимано-Североуральского бассейна. Их существование и расселение на шельфе Тимано-Североуральского морского бассейна было обусловлено множеством факторов, среди которых важнейшими были обширные территории мелководных областей седиментации с выровненным рельефом дна и низкой энергией водной среды, формирование органогенных построек, процессы миграции. Особенности рас- пространения брахиопод отражают их приурочен- ность к определенным фациальным зонам бассейна (прибрежным, литоральным, мелководно-шельфовым). Нередко они являются единственными ископаемыми и породообразующими в отложениях различных фаций. Изученность брахиопод, установленные рубежи существенного преобразования морфо-экологической структуры и фациальная приуроченность их сообществ в Тимано-Северо-уральском морском бассейне [3], послужили материалом для исследований по выявлению биохимической реакции этих древних бентосных орга- аминокислот в раковинах современных брахиопод побережья Японии и раковинах плейстоценового возраста (1.47 млн. лет) показали, что практически все типы брахиопод характеризуются высоким содержанием глицина [4]. В некоторых случаях его количество достигает 40 % от общего содержания аминокислот, при этом в отличие от моллюсков аминокислотный состав брахиопод характеризуется низкими концентрациями кислых аминокислот – аспарагиновой и глутаминовой. В современных брахиоподах Новой Зеландии доминирующей аминокислотой также является глицин [5]. На его долю приходится от 30 до 50 % всех аминокислот, далее следуют аланин, пролин и валин или же аспарагиновая и глутаминовая кислоты. В современных брахиоподах Шотландии установлено присутствие всех белковых аминокислот [6]. Их распределение меняется в зависимости от минерального состава раковины. В апатитовой раковине доминируют аспарагиновая кислота, глицин и аланин, а в кальцитовых раковинах брахиопод – глицин (рис. 1). Высокие содержания аланина, аспарагиновой кислоты, глицина, а также присутствие гидроксипролина за-
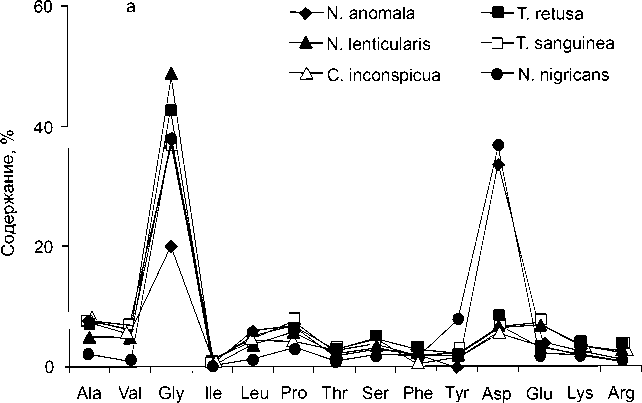
низмов на изменение внешних условий среды в течение позднего ордовика, силура и раннего девона .
Цель данной работы заключалась в выявлении возможных отклонений в содержании аминокислот в раковинах ископаемых брахиопод на территории Тимано-Северо-уральского осадочного бассейна, а также сопоставление полученных данных с выполненными ранее палеонтологическими, биостратиграфи-ческими и геохимическими исследованиями [1, 3].
Состав аминокислот изучен во многих современных и
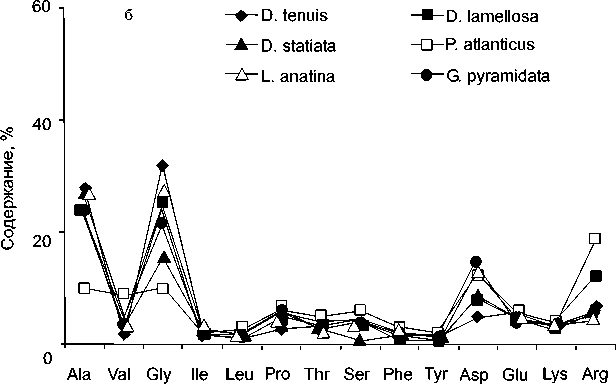
Рис. 1. Распределение аминокислот в современных брахиоподах: а – кальцитовая раковина, б – апатитовая раковина [6].
ископаемых раковинах бра-хиопод. Исследования состава
фиксировано в фосфатсодержащих раковинах бра-хиопод и другими исследователями [7]. Аминокислотный состав раковин современных брахиопод успешно используется для отнесения их к тому или иному таксономическому ряду, а также в качестве геотемпературного индикатора [4–5, 7–8].
Заметные изменения в составе и содержании аминокислот наблюдаются в раковинах брахиопод в течение первых нескольких сотен тысячелетий после захоронения. По данным исследователей [4], процесс деградации макромолекул довольно значителен в течение первых 0.13 млн. лет, затем наступает устойчивая фаза. Следующая фаза деградации начинается после 0.62 млн. лет и отчетливо прослеживается до образцов с возрастом в 1.47 млн. лет. Наиболее сильные диагенетические изменения белковой составляющей раковин – деградация белков до пептидов и потеря менее стабильных аминокислот в ходе перекристаллизации раковинного вещества – происходят еще до наступления верхнемелового периода [6]. В определенных условиях осадконакопления наличие белковых соединений в ископаемых раковинах может сохраняться вплоть до юры и мела [4]. Однако обычно в фоссилизированных створках содержание аминокислот падает, а их групповой состав часто представлен только алифатическими и кислыми аминокислотами [6]. После полной перекристаллизации ископаемые раковины характеризуются малым количественным и качественным составом аминокислот [6, 9], в связи с чем их использование как таксономического признака и геотемпературного индикатора становится практически невозможным, особенно в палеозойских и более древних образцах.
Большое влияние на состав аминокислот оказывают геохимические условия осадкообразования и дальнейшей геологической и геохимической истории осадочных пород [9]. Этим объясняются факты значительного содержания аминокислот в некоторых раковинах ископаемых организмов древнейших геологических эпох и ние или полное отсутствие аминокислот в раковинах более молодого геологического возраста. Так, исследования аминокислот в брахиоподах рода Crania [6] показали, что в верхнемеловых раковинах аминокислотный состав значительно отличается от состава современных раковин, несмотря на достоверно установленное присутствие в них белка (рис. 2). Cодержание аланина в раковинах, имеющих возраст 130 тыс. лет и более, увеличивается вдвое, по-сравнению с современными образцами. В раковинах карбона и ордовика распределение индивидуальных аминокислот претерпевает дальнейшее изменение, однако общее содержание аминокислот в ордовике более чем в два раза превышает их ко- малое содержа-
личество в раковинах карбона, тогда как содержание аминокислот во вмещающих породах находится на одном концентрационном уровне (11 пмоль/мг для карбона и 7–11 пмоль/мг для ордовика). Значительные содержания аминокислот в ордовикских раковинах объясняются особыми условиями их захоронения в углистых аргиллитах (Северная Эстония, карадок, кукруский горизонт и верхний арениг, волковский горизонт, Санкт-Петербург).
Методы исследования
Изучение состава аминокислот в раковинах Тимано-Североуральских брахиопод раннего палеозоя и вмещающих их породах проводились по методике [10] . Анализ включает в себя гидролиз образцов, их последующую очистку и перевод выделенных аминокислот в сложные эфиры с последующим разделением методом газовой хроматографии.
Гидролиз. Предварительно образцы растирались и обрабатывались бидистиллированной водой при 100 ° С. Для извлечения связанных аминокислот образцы подвергаются кислотному гидролизу в 6М HCl при температуре 105 ° С в течение 12 час. Данная температура была выбрана с той целью, чтобы исключить риск рацемизации аминокислот.
Очистка образцов. Полученные гидролизаты фильтруются, добавляется внутренний стандарт, в качестве которого использовался L-норвалин. Затем содержимое отгоняется на роторном испарителе при температуре 45 ° С и перерастворяется в 0.05М HCl. Очистка полученного раствора проводится на колонке, заполненной Dowex 50WX8 Na + (катионообменник). Для удаления ионов металлов смолу промывают 0.01М HCl. Элюирование аминокислот осуществляется с помощью 2.5М NH 4 OH. Полученный раствор вновь отгоняется на роторном испарителе при температуре 45 ° С.
Дериватизация. Образцы растворяют в 0.1М HCl, полученные растворы замораживают и высушивают в замороженном состоянии. Высушенные
Neocrania
Crania craniolaris
53.4
11.7
19.4
Pseudocrania petropolitana o1
21.7
Q.
АК, пмоль/мг 260.4
K
2
C
O
3
Crania rhykholtiana Orthisocrania planissima
Рис. 2. Содержание аминокислот в современных и ископаемых бра-хиоподах рода
Crania
по [6].
Ala Val Gly Ile Leu Pro Thr Ser Phe Tyr Asp Glu Lys Arg
образцы этерифицируются изопропиловым спиртом, а затем ацилируются пентафторпропионовым (PFP) ангидридом при температуре 110
°
С. Полученные N-пентафтор-пропионовые изопропиловые эфиры аминокислот растворяют в дихлорметане и переносят в виалы для хроматографии.
Газохроматографический анализ.
Анализы выполняются на газовом хроматографе GC-17A с пламенно-ионизационным детектором. Для разделения энантиомеров полученных эфиров аминокислот используется колонка Chirasil-L-Val (длина 25 м, внутренний диаметр 0.25 мм). В качестве газа-носителя применяется гелий. Температура испарителя - 250
°
С, температура пламенно-ионизационного детектора - 275
°
С. Анализ проводится при следующих условиях программирования температуры термостата колонок: 4 мин в изотермическом режиме при температуре 90
°
С, затем подъем температуры до 210
°
С со скоростью 4
°
С/мин. Продолжительность анализа – 37 мин.
Все реагенты, используемые в работе, имеют высокую степень чистоты (Sigma, Aldrich, Fluka компаний), для приготовления растворов применяется бидистиллирован-ная вода. Кроме того, параллельно с образцами ведется холостой опыт, чтобы исключить попадание аминокислот из реагентов. Объекты исследования
Исследования аминокислотного состава выполнены для брахиопод и вмещающих их карбонатных пород из отложений верхнего ордовика, силура и нижнего девона западного склона Северного и Приполярного Урала, гряд Чернышева и Чернова, Северного Тимана, а также скважин 76-Лабаганская и 2-Адакская (рис. 3). В качестве объектов для изучения характера изменения состава аминокислот в раковинах были выбраны представители отрядов пентамерид, строфоменид, атрипид, спи-риферид и ринхонеллид. Все образцы имеют точную стратиграфическую и фациальную привязку
.
Основная доля исследований выполнена для гряды Чернышева и Приполярного Урала (бассейн р. Кожим), где проанализированы образцы из обнажений, выходящих на дневную поверхность практически для всех стратиграфических горизонтов силура. Из-за отсутствия достаточного количества брахиоподовой фауны в коллекциях не удалось изучить раковины лола-шорского горизонта на гряде Чернышева, а также маршрутнинского и войвывского горизонтов на западном склоне Приполярного Урала.
Тимано-Североуральские карбонаты (преимущественно известняки доломитизи-рованные и доломиты вторичные) претерпели значительные катагенетические преобразования от стадий МК
2
на Северном
Тимане до АК
1
–АК
3
на западном склоне Северного и Приполярного Урала. В большинстве случаев перекристаллизованные остатки брахиопод и вмещающие их породы представлены кальцитом и доломитом, часто с небольшими примесями кварца и слюды (дифрактометр Shimadzu XRD-6000)
.
Характеристика образцов дана в табл. 1.
Состав и содержание аминокислот Содержание аминокислот широко варьирует в изученных раковинах (190–10 мкг/г раковины) и практически не изменяется в породах (60–20 мкг/г породы). Максимальное содержание аминокислот выявлено в раковинах венлокских брахиопод р. Шаръю (100–190 мкг/г раковины). Состав аминокислот в ископаемых раковинах неоднороден (табл. 2) и в значительной степени отличается от состава белковых аминокислот современных брахиопод. В большинстве изученных образцов наблюдается ус- Рис. 3. Схема расположения изученных разрезов верхнего ордовика и силура. Обнажения: 1 – руч. Третий, обн. 1; 2 – руч. Безымянный, обн. 2; 3 – руч. Дершор, обн. 1, 2; 4 – р.Уса, обн.246; 5 – р. Поварница, обн. 192; 6 – р. Неча, обн. 7; 7 – р. Шарью, обн. 64, 24; 8 – р. Изъяю, обн. 479, 481 ; 9 – р. Сыня, обн. 4; 10 – р. Илыч, обн. 23; 11 – р.Унья; р. Кожим: 12 – обн.108, 13 – обн.116, 14 – обн. 221, 15 – обн.196; 16 – обн. 236. Скважины: 1 – 76-Ла-баганская; 2 – 2-Адакская. Таблица 1 Характеристика изученных образцов и содержание аминокислот в раковинах брахиопод и вмещающих породах
Ярус, горизонт
Место отбора
Номер образца
Вид раковины
∑АК, мкг/г раковины
∑АК, мкг/г породы
Состав раковины
Состав породы
Стадия катагенеза
Приполярный Урал
D
1
Лохков Овин-пармский
р. Кожим
236/228
Protathyris praecursor
24.1
–
–
–
МК
4
-МК
5
S
2
Пржидол
Карповский
236/20a
Collarothyris cana-liculata
42.3
34.9
кальцит кварц
кальцит кварц
МК
4
-МК
5
Пржидол
Белушьинский
236/193
Collarothyris cana-liculata
40.7
30.4
–
–
МК
4
-МК
5
236/248
Grebenella par-vula
10.6
–
–
–
МК
4
-МК
5
236/20b
Atrypoidea scheii
26.0
28.7
–
–
МК
4
-МК
5
Лудлов Сизимский
214/1
Didymothyris di-dyma
50.9
–
–
–
МК
4
-МК
5
Лудлов Падимейты-висский
211/173
Greenfieldia uberis
14.7
–
–
–
МК
4
-МК
5
S
1
Лландовери Филиппьель-ский
196/107
Pseudoconchidium kozhimicum
45.2
33.4
кальцит
доломит кальцит
МК
4
-МК
5
194/1028
Pseudoconchidium kozhimicum
46.5
20.3
кальцит кварц доломит
кальцит доломит
МК
4
-МК
5
Лландовери, Лолашорский
108/23
Pentamerus (?) sp.
50.4
59.6
–
–
МК
4
-МК
5
116/10
Pentamerus (?) sp
75.2
60.6
–
–
МК
4
-МК
5
196/17
Pentamerus (?) sp
82.8
26.4
кальцит кварц доломит
кальцит кварц
МК
4
-МК
5
Лландовери Яренейский
196/38
Zigospiraella du-boisi
72.8
–
–
–
МК
4
-МК
5
O
3
Ашгил
Кырьинский
224/4
Holorhynchus giganteus
38.6
47.6
доломит кальцит
доломит кальцит
МК
5
-АК
1
114/6
Proconchidium muensteri
37.2
17.9
–
доломит кальцит
МК
5
-АК
1
Гряда Чернышева
D
1
Лохков
Овинпармский
р. Изъяю
481/510
Howellella angusti-plicata
39.5
–
–
–
МК
3
-МК
4
S
2
Пржидол
Карповский
руч. Дэр-шор
2/15
Atrypoidea scheii Collarothyris cana-liculata
20.7
29.7
кальцит кварц
кальцит кварц доломит
МК
3
-МК
4
1/11
Collarothyris cana-liculata
32.1
49.3
–
кальцит доломит кварц
МК
3
-МК
4
Пржидол
Белушьинский
2/17
Atrypoidea scheii
17.0
52.7
кальцит кварц
кальцит кварц
МК
3
-МК
4
Лудлов Сизимский
скв. 2-
Адакская
4232.0
4238.8 м
Didymothyris di-dyma
92.6
18.4
–
–
МК
3
-МК
4
Лудлов Падимейты-висский
р. Изъяю
479/1631
Greenfieldia uberis
40.6
–
–
–
МК
3
-МК
4
S
1
Венлок
Войвывский
р. Шаръю
64/257
Spirinella norden-sis
102.9
54.4
кальцит кварц доломит
кальцит доломит кварц
МК
3
-МК
4
24/42
Protatrypa inflata
194.4
43.1
кальцит кварц
кальцит доломит кварц
МК
3
-МК
4
Лландовери, Маршрутнин-ский
64/294
Leptaena sp.
41.0
64.8
кальцит доломит кварц
кальцит доломит кварц
МК
3
-МК
4
Лландовери
Филиппьельский
р. Неча
7/12
Virgianella vai-gatschensis
49.3
54.9
–
–
МК
3
Лландовери Яренейский
р. Повар-ница
192/663
Virgiana barrandei
53.4
57.3
кальцит кварц доломит
кальцит кварц доломит
МК
3
-МК
4
р. Б.Уса
246/Л
Becscia mennery
36.9
37.9
–
–
МК
3
-МК
4
246/881
Virgiana adakia
37.1
49.8
кальцит кварц доломит
кальцит доломит кварц
МК
3
-МК
4
Окончание табл. 1 АК – аминокислоты
Ярус, горизонт
Место отбора
Номер образца
Вид раковины
∑АК, мкг/г раковины
∑АК, мкг/г породы
Состав раковины
Состав породы
Стадия катагенеза
O
3
Ашгил Кырьинский
р. Сыня
4/15
Proconchidium muensteri
23.0
44.0
–
–
МК
4
-МК
5
Северный Урал
S
2
Лудлов Сизимский
р. Илыч
23/118
Conchidium no-vosemelicum
76.9
42.3
кальцит
кальцит кварц
МК
5
-АК
1
S
1
Лландовери Филиппьельский
р. Унья
37/2823
Virgianella vai-gatschemsis
56.4
22.9
–
кальцит доломит
МК
5
-АК
1
Гряда Чернова
S
1
Венлок
Войвывский
руч. Безымянный
2/387
Spirinella nordensis
72.1
–
–
–
Северный Тиман
S
1
Лландовери Лолашорский
руч. Третий
1/3
Pseudocama rotechia (?) sp
165.4
52.6
доломит кварц кальцит
доломит кварц
МК
2
-МК
3
Ва
рандей-Адзьвинская зона
D
1
Лохков Овинпармский
скв. 76-Лабаган-ская
4117
4123 м
Lenatoechia kuliki
81.7
35.6
кальцит кварц доломит
кальцит доломит кварц
нач. МК
3
S
2
Пржидол Карповский
4483 м
Collarothyis cana-liculata
52.3
44.3
–
кальцит кварц доломит
нач. МК
3
Таблица 2 Содержание аминокислот в раковинах брахиопод (% от общего количества аминокислот)
№ образца
Ala 1
Val 1
Gly 1
Ile 1
Leu 1
Pro
Thr 1
Ser
Phe
Tyr 1
Asp 1
Glu 1
Lys 1
Met
Приполярный Урал
236/228
11
8
15
0
11
3
0
14
17
2
4
9
4
0
236/20a
7
9
15
1
10
1
0
25
11
0
12
5
2
0
236/193
12
6
14
1
7
7
0
9
34
0
3
6
1
0
236/248
8
8
13
2
10
3
0
9
36
2
2
7
1
0
236/20b
8
7
15
1
11
2
0
11
27
0
4
9
3
0
214/1
12
7
19
0
9
3
0
19
12
3
5
8
2
0
211/173
19
10
20
10
10
4
2
17
3
0
0
5
0
0
196/107
6
7
14
7
3
11
10
9
21
5
0
8
0
0
194/1028
5
3
8
1
6
4
3
2
24
0
3
41
0
0
108/23
12
9
12
1
6
7
0
12
14
2
3
18
3
0
116/10
11
9
12
1
7
6
0
9
15
7
4
16
3
0
196/17
3
0
8
0
10
1
0
15
5
29
5
19
5
0
196/38
13
8
20
1
9
11
0
11
5
1
6
11
4
0
224/4
8
7
13
4
9
5
1
9
15
2
6
17
3
0
4/114
12
8
17
2
12
5
1
7
10
4
5
11
6
0
Гряда Чернышева
481/510
12
12
15
0
10
6
0
10
17
2
4
8
1
1
2/15
12
8
26
1
9
6
0
6
6
4
4
12
5
0
1/11
11
6
20
2
14
5
0
14
9
2
4
12
2
0
2/17
14
5
21
0
11
5
0
12
12
0
3
13
3
0
4232
11
9
21
0
7
5
0
17
7
5
5
9
4
0
479/1631
13
10
20
0
8
6
0
16
13
2
4
7
1
1
64/257
9
7
12
4
9
0
4
19
18
2
2
8
1
5
24/42
7
7
14
1
7
1
1
22
7
2
8
23
0
1
64/294
6
18
7
2
9
8
0
24
6
4
2
11
1
3
7/12
12
12
12
1
9
8
0
5
20
4
5
10
3
0
64/663
10
0
21
1
15
1
0
26
8
1
3
10
5
0
246/Л
8
9
15
17
6
0
1
4
20
0
5
13
0
2
246/881
10
7
13
11
9
5
2
6
19
2
5
9
3
0
4/15
9
10
17
0
11
9
2
7
16
0
3
11
1
3
Северный Урал
23/118
9 1
5 1
7 1
18 1
6 1
0
0
30
9 1
0 1
9 1
8 1
0 1
0
37/2823
9 1
3 1
12 1
6 1
14 1
0
0
15
12 1
8 1
2 1
15 1
3 1
0
Гряда Чернова
2/387
1
13 1
10 1
13 1
4 1
10 1
4 1
0 1
8 1
17 1
4 1
3 1
10 1
4 1
0
Северный Тиман
1/3
1
12 1
7 1
19 1
1 1
14 1
7
0 1
6 1
10 1
1 1
7 1
12 1
2 1
1
Скважина 76-Лабаганская
4117
15 1
0 1
4 1
3 1
11 1
0 1
0 1
31 1
14 1
2 1
2 1
13 1
6 1
0
4483
8
5
17
2
18
5
0
8
9
3
5
19
0
1
Прнмечание. Ala – аланин, Val – валин, Gly – глицин, Ile – изолейцин, Leu – лейцин, Pro – пролин, Thr – треонин, Ser – серин, Phe – фенилаланин, Tyr – тирозин, Asp – аспарагиновая кислота, Glu – глутаминовая кислота, Lys – лизин, Met – метионин. тановление баланса аминокислот между раковиной и вмещающей ее карбонатной породой. В ряде случаев процесс диффузии аминокислот из скелетных остатков приводит к тому, что содержание аминокислот во вмещающей породе даже несколько больше, чем в самой раковине. Преобладают алифатические аминокислоты, содержание которых в среднем составляет 40–50 %, распределение кислых, гидроксильных и ароматических групп варьирует, однако чаще второй доминирующей группой являются ароматические аминокислоты (15–20 %). Среди индивидуальных аминокислот в изученных раковинах доминируют глицин, фенилаланин, серин и глутаминовая кислота. Обычно на их долю приходится от 45 до 70 % от всего содержания аминокислот. Повышенные концентрации установлены также для аланина и лейцина (до 10%). В большинстве образцов отмечено присутствие D-аллоизолейцина, аминокислоты образующейся при высоких температурах. Повышенные содержания D-аллоизолейцина характерны для образцов Приполярного и Северного Урала, что может быть объяснено более высокими температурами катагенетического преобразования исследованных пород. Отличительная особенность образцов гряды Чернышева – постоянное присутствие в них серосодержащей аминокислоты – метионина. Скорее всего, метионин является новообразованным за счет реакций вторичного биохимического синтеза (в восстановительных условиях, при достаточном доступе сероводорода и сульфатов). Выходы сероводородных источников на гряде Чернышева в долинах рек Уса, Поварница, Адак отмечали Н.А.Кулик [11], А.А.Чернов [12] и другие исследователи. Источники поступления вторичных соединений серы на гряде Чернышева связаны с наличием здесь зон разломов и трещиноватости силурийских доломитов и известняков [13]. Выявленные вариации в содержании и составе аминокислот палеозойских раковин связаны с влиянием ряда факторов. В раковинах брахиопод, захороненных в одно и то же время, состав и количество аминокислот в значительной степени зависят от первичных условий осадконакопления и фациальной приуроченности, типа захоронения. Большое влияние на распределение аминокислот в ископаемых раковинах брахиопод оказали процессы катагенетического прогрева пород Тимано-Севе-роуральского бассейна, а также процессы окисления материала в естественных обнажениях. Для образцов, имеющих практически один уровень катагенетического прогрева пород, одной из причин отличия в содержании аминокислот в раковинах брахиопод могут являться первичные условия их захоронения. Так, плохая сохранность аминокислотного состава в раковинах Leptaena sp. (рис. 4, 7) из маршрутнинского горизонта лландове-ри и Сollarothyris canaliculata из верхов пржидола гряды Чернышева, представленных в виде отдельных не сортированных створок в породе, вероятно, связана с тем, что погребение этих раковин проходило в тиховодных участках дна с глинистыми грун- тами и слабым течением, в результате чего их органическое вещество было изменено или даже разрушено еще на начальных этапах захоронения. В захоронениях сообществ брахиопод войвывского горизонта венлока Spirinella nordensis (рис. 4, 4, 4а) и Protatrypa inflatа (рис. 4, 6) на гряде Чернышева присутствуют скопления целых сочлененных раковин разных размеров и различных возрастных стадий – от ювенильных до зрелых, с хорошо сохранившимися скульптурой, элементами внутреннего строения, а также тонкими иглами на поверхности раковины S. nordensis (рис. 4, 4а). Все перечисленные признаки характерны для прижизненных захоронений брахиопод или погребений вблизи места обитания, например, во время штормов. В подобных условиях белковая составляющая раковин могла остаться практически неизмененной на начальных этапах диганетических преобразований, что и отразилось на столь высоком содержании в них аминокислот, несмотря на дальнейшие вторичные изменения. Следует отметить, что захоронения Virgiana barrandei из яренейского горизонта ллан-довери гряды Чернышева и Virgianella vaigatchensis (рис. 4, 1) из рифогенных отложений филиппьель-ского горизонта лландовери содержат раковины разных возрастных стадий – от мелких (до 1 см) до более крупных (4 см), значительная часть которых сохранилась в прижизненном положении, макушкой вниз на брюшной створке. Однако эти образцы имеют более высокие температуры катагенетиче-ского прогрева, с чем, возможно, и связано более низкое содержание в них аминокислот. Установленное повышенное содержание аминокислот в раковинах Pentamerus sp. (рис. 4, 5) и Zigospiraella duboisi (рис. 4, 9) из лландоверийских отложений Приполярного Урала также хорошо согласуется с прижизненными условиями их захоронения.
Среди изученных образцов Тимано-Северо-уральского бассейна наиболее низкие температуры прогрева характерны для отложений Северного Тимана и Варандей-Адзьвинской зоны (скв. 76-Ла-баганская), что сразу же отражается на повышенном содержании аминокислот в раковинах
Pseudo-camarotechia
(?) sp. (рис. 4,
12
),
Collarothyris cana-liculata
(рис. 4,
14
) и
Lenatoechia kuliki
(рис. 4,
11
)
.
Кроме того, для образцов, отобранных из скважин, в сравнении с образцами из естественных обнажений в одних и тех же возрастных интервалах характерны более высокие содержания аминокислот. Это, скорее всего, отражает лучшую сохранность скважинного материала и свидетельствует о влиянии окислительных факторов в условиях естественных обнажений, способствующих дальнейшему разрушению органического вещества ископаемых организмов. Анализируя характер захоронений брахиопод в изученных разрезах верхнего ордовика, силура и нижнего девона следует отметить значительное влияние на их развитие и территориальное распространение условий осадконакопления, связанных с возможностью миграции или изолированностью бассейна, опреснением или соленостью его вод, течениями, штормами, увеличением поступления терригенного материала, изменениями
Рис. 4. Брахиоподы силура и нижнего девона. 1 –
Virgianella vaigatshensis
Nikiforova
,
х1, Северный Урал, р. Унья, обн.37, обр. 2822; 2 –
Pseudoconchidium kozhimicum
Nikiforova, х1, Приполярный Урал, р. Кожым, обн.196, обр.498; 3 –
Conchidium novosemelicum
Nalivkin , х1, Северный Урал, р. Илыч, обн. 23Т, обр. 251; 4 –
Spirinella nordensis
(Lashenko), х1,5, р.Шаръю, обн.64; 4a – фрагмент створки раковины с сохранившимися иглами; 5 –
Pentamerus
sp. Приполярный Урал, р. Кожым, обн. 108, обр. 25; 6 –
Protatrypa inflata
T.Beznosova, х0,5. Гряда Чернышева, р. Шаръю, обн. 64, обр. 116; 7 –
Leptaena
sp., х1. Гряда Чернышева, р. Шаръю, обн. 64, обр. 3; 8 –
Becscia menneri
(T.Beznosova), х2. Гряда Чернышева, р. Уса, обн. 246, обр. 146; 9 –
Zygospiraella duboisi
(Verneuil), х0,5. Приполярный Урал, р. Кожым, обн. 196, обр. 153; 10 –
Didymothyris didyma
(Dalman), х1. Приполярный Урал, р. Кожым, обн. 214, обр. 35; 11 –
Lenatoechia kuliki
Nikiforova, х1,5, скв. 76-Лабаганская, гл. 4117-4123 м; 12 –
Pseudocamarote-chia
(?) sp., х2, Северный Тиман, руч.Третий, обн. 1, обр. 3; 13 –
Protathiris praecursos
(Kozlovsky), х1,2, скв. 76-Лабаганская, гл. 4117-4123; 14 –
Collarothyris canaliculata
(Wenjukov), х1,2, скв. 76-Лабаганская, гл. 4483 м.
рельефа дна. По мере масштабности проявления абиотических факторов выделены основные типы захоронений брахиопод [3]. Аминокислоты – индикаторы событийных факторов Результаты изучения раковин брахиопод показали изменение содержания в них аминокислот на рубеже ордовика и силура (рис. 5): увеличение концентрации аминокислот от 25–40 мкг/г в ордовике до 50–80 мкг/г в раннем лландовери (яренейское и ранне лолашорское время), уменьшение до 40 мкг/г в конце лландовери (маршрутнинское время) и резкий подъем до 70–190 мкг/г в венлоке (вой-вывское время), снижение в раннем лудлове до 15– 40 мкг/г (падимейтывисское время), увеличение до 70–90 мкг/г в сизимское время, значительное снижение в раннем пржидоле до 20 мкг/г (белушьин-ское время) и небольшое повышение в карповское и овинпармское время 50 мкг/г. В истории осадконакопления Тимано-Се-вероуральского морского бассейна в позднем ордовике, силуре и раннем девоне выявляется шесть фаз относительного повышения уровня моря, наи- Горизонт Карповский Сизимский Войвывский Яренейский Кырьинский Падимейты-висский Овинпарм-ский Разрез Интервал распрост ранения брахиопод Содержание аминокислот, мкг/г 100 150 200
Приполярный Урал
гр. Чернышева
трансгрессия
содержание, мкг/г
содержание, мкг/г
0 50 100 150 200
0 50 100 150 200
регресиия
■ 0
Ой Лолашорский Маршрутнин-ский Филиппъель-ский Юнкошорские слои -^l lv ill II IV
I
у
I I
1Й
^I ^II
^
III IV V
• раковина о порода
| Ч/ |VI I
/О 1
VII VIII | аа | IX | о~| X | | фауна отсутствует
Рис. 5. Изменение содержаний аминокислот в ископаемых брахиоподах и карбонатных породах Тимано-Североуральского бассейна. I – известняк, II – доломит, III – известняк доломитовый, IV – известняк биогенный, V – рифовые постройки, VI – брахиоподы, VII – водоросли, VIII – табуляты, IX – строматолиты, X – остракоды; брахиоподы: 1 –
Proconchidium muensteri,
2 -
Holorhynchus giganteus
, 3 –
Becscia mennery
, 4 –
Virgiana adakia
, 5 –
Zigospiraella duboisi
, 6 –
Virgiana barrandei
, 7 –
Pseudocamarotechia (?) sp
, 8 –
Pentamerus (?) sp
., 9 –
Pseudoconchidium kozhimicum
, 10 –
Virgianella vaigatschensis
, 11 –
Leptaena sp
., 12 –
Spirinella nordensis
, 13 –
Protatrypa inflata
, 14 –
Greenfieldia uberis
, 15 –
Conchidium novosemelicum
, 16 –
Didymothyris didyma
, 17 –
Atrypoidea scheii
, 18 –
Grebenella parvula
, 19 –
Collarothyris canaliculata
, 20 –
Lenatoechia kuliki
, 21 –
Protathyris praecursor
, 22 –
Howellella augustiplicata
.
более крупное из них – в первой половине лландо-вери. На графике изменения содержания аминокислот в раковинах отмечаются три максимума, которые соответствуют трем фазам повышения относительного уровня моря в мелководно шельфовом Тимано-Североуральском морском бассейне в силуре – раннем лландовери, венлоке и луд-лове (рис. 5). Первый максимум совпадает с самой значительной трансгрессией в начале лландовери. Средняя часть яренейского и нижняя часть лола-шорского горизонтов отвечают максимальному повышению уровня моря с накоплением мощной толщи нормально морских отложений – темноцветных массивных известняков с прослоями глинистых иловых известняков, заключающих сообщества брахиопод, обитавших в зоне нижней сублиторали и характеризующихся значительной экологической специализацией. Массивные известняки заключают ракушняковые прослои и банки с пентамеридами Virgiana, Pentamerus и Borealis, обладающими крупными, толстостенными гладкими и ребристыми раковинами с массивной макушкой и обитавшими на твердых скалистых грунтах. К прослоям иловых известняков приурочены скопления мелких тонкостенных ребристых, уплощенных раковин Zygospi-raella, адаптированных к условиям широкого развития иловых осадков. В отложениях рассматриваемого временного интервала выделяются два основных типа захоронений брахиопод: скопления целых сочлененных раковин разных размеров и различных возрастных стадий, характерные для прижизненных захоронений (рис. 4, 1, 2) или целых раковин вблизи места захоронения (рис. 4, 8а–8в, 12–14) и захоронения в виде скоплений лома раковин и их крупных фрагментов прослеживаемые на значительной территории, что связано с условиями резкой трансгрессии. На таких скоплениях лома раковин нередко наблюдается нарастание пластовых колоний строматопороидей. Очевидно, это происходило в период стабилизации условий осадконакопления и ракушняки служили субстратом для других бентосных сообществ. Постепенное падение уровня моря к концу лолашорского времени обусловившее обмеление всех фациальных зон, продолжалось и в филиппьельское время, с которым связано начало рифообразования в силуре, формирование зарифовых лагун с преимущественно иловым осадконакоплением и широким развитием цианобактериальных сообществ. В наиболее мелководных областях западной части Тимано-Северо-уральского палеобассейна прекратилось карбонатное осадконакопление, прибрежная зона сместилась к восточной части бассейна. В филиппьель-ское время существование брахиопод пентамерид (Virgianella, Pseudoconchidium) было связано лишь с рифовыми биогермными постройками на территории современного Предуральского краевого прогиба и западного склона Урала. Трансгрессия в начале маршрутнинского времени была менее значительна, чем в начале лландовери. За непродолжительной стабилизацией экосистемы в среднемар-шрутнинское время последовало обмеление бассейна с формированием литоральных фаций и широким развитием цианобактериальных сообществ, что привело к исчезновению в конце лландовери бентосной фауны, в том числе брахиопод Leptaena, Atrypa и Fardenia. Захоронения брахиопод в отложениях маршрутнинского горизонта представляют собой скопления отдельных, преимущественно брюшных створок раковин в породе без следов ориентировки. Такой тип захоронения образуется в тиховодных участках бассейна со слабым течением, которое выносит более легкие спинные створки. Биотический кризис на рубеже лландовери и вен-лока был результатом регрессии, достигшей максимума в конце лландовери.
Наиболее отчетливо выраженный максимум содержания аминокислот, достигающий более 100 мкг/г, зафиксирован в образцах, отобранных непосредственно выше уровня границы лландовери-венлок
.
Открытоморские условия начала венлока способствовали существенному обновлению родового и видового состава брахиопод. Морфологические особенности раковин венлокских брахиопод Spirinella свидетельствуют о возможности их адаптации к различным условиям обитания в зоне нижней сублиторали.
Третий максимум повышенных концентраций аминокислот приходится на поздний лудлов (си-зимский горизонт). Начало лудловского века характеризуется накоплением преимущественно карбонатно-терригенных отложений в крайне мелководных условиях и крайне бедным родовым и видовым составом брахиопод(Greenfieldia) и другого бентоса. На этом фоне значительным биоразнообразием выделяются рифовые и биогермные постройки, формировавшиеся в восточной части бассейна в сизимское время. Захоронения брахиопод, обитавших вблизи рифовых построек (пентамеровое сообщество Conchidium), толстостенные грубо скульп-турированные раковины которых обнаруживают большое морфологическое сходство с лландоверий-скими пентамеридами, представлены преимущественно отдельными створками (рис. 4, 3) и их крупными фрагментами. Отложения зарифовых фаций нередко заключают захоронения целых сочлененных раковин брахиопод Didymothyris (рис. 4, 10).
Полученные результаты не противоречат палеонтологическим данным, свидетельствующим о крупных изменениях Тимано-Североуральской морской биоты, в том числе и сообществ брахиопод, приуроченных к рубежам ордовик–силур, лландо-вери–венлок, силур–девон. Эти изменения, сопровождающиеся преобразованием таксономического состава разных групп организмов, их численности, вымиранием отдельных таксонов, имеют как региональные, так и глобальные проявления. Так существенные изменения западноуральской биоты, сокращение ее разнообразия, зафиксированные в разрезе пограничных толщ ордовика и силура, совпадают с крупнейшей биосферной перестройкой на этом рубеже (событие хирнантия) [3]. Перестройка тимано-североуральской биоты на рубеже лландо-вери и венлока сопоставляется с масштабным океанографическим событием Иревикен [1]
.
Переход от силура к девону хорошо диагностируется в разрезах по вымиранию силурийских таксонов и появлением новой фауны в девоне. Проявление этих биотических событий в регионе распознается и по данным изотопного анализа [1]
.
Заключение Результаты исследований аминокислот раннепалеозойских брахиопод свидетельствуют о том, что возраст ископаемых раковин не является решающим для сохранности и преобразования органического вещества. Состав и содержание аминокислот в древних раковинах брахиопод зависят от ряда факторов. В первую очередь – это первичные условия осадконакопления, которые связаны с возможностью миграции или изолированностью бассейна, опреснением или соленостью его вод, течениями, штормами, увеличением поступления терригенного материала, изменениями рельефа дна. Несомненно, что большое влияние на состав органического вещества оказали процессы катагенети-ческого прогрева пород Тимано-Североуральского бассейна. Кроме того, вторичное перераспределение материала вмещающих пород, а также окислительные условия в районах естественных обнажений также способствовали дальнейшему разрушению и изменению органического вещества ископаемых организмов. Увеличение содержания аминокислот в раковинах раннепалеозойских брахиопод приурочено к трем интервалам силурийского разреза, формирование которых совпадает с тремя эвстатическими трансгрессиями. Анализ полученных данных пока- зал, что изменения происходили на фоне разномасштабных экосистемных перестроек, границы которых приурочены к границам ярусов и зон. Дальнейшие исследования позволят выявить возможность применения аминокислотного анализа как дополнительного критерия при распознавании важных событийных уровней в раннем палеозое. Авторы выражают признательность академику Н.П.Юшкину за помощь в работе, к.г.-м.н. Л.А.Ани-щенко за предоставленные данные по катагенезу осадочных толщ Тимано-Североуральского бассейна, Б.А. Макееву за проведение аналитических измерений, В.П. Волковой за фотографии брахиопод. Работа выполнена при поддержке Программы фундаментальных исследований РАН № 28 проекты № 12-П-5-1011, 12-П-5-1015 и гранта Президента РФ НШ-1310.2012.5.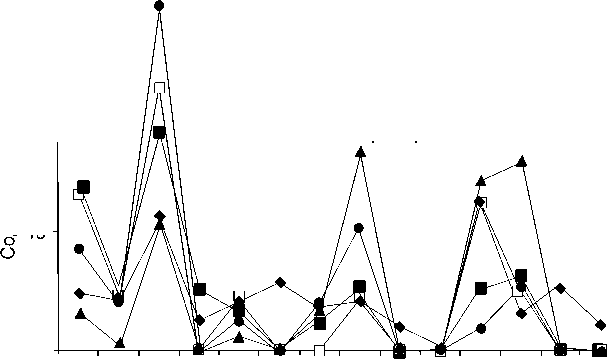


Список литературы Аминокислоты в раннепалеозойских брахиоподах Тимано-Североуральского бассейна
- Безносова Т.М., Мянник П.Э. Граница лландовери и венлока на севере палеоконтинента Балтия//ДАН. 2005. Т. 401. № 5. С. 1-4.
- Чехович П.А., Живкович А.Е., Медведовская Н.И., Степанова Н.А. Изотопные реперы в палеозойских разрезах Урала//ДАН СССР. 1990. Т. 313. № 2. С. 423-426.
- Безносова Т.М. Сообщества брахиопод и биостратиграфия верхнего ордовика, силура и нижнего девона северо-восточной окраины палеоконтинента Балтия. Екатеринбург: УрО РАН, 2008. 217 с.
- Endo K., Walton D., Reyment R.A., Curry G.B. Fossil intra-crystalline biomolecules of brachiopod shells: diagenesis and preserved geo-biological information//Organic Geochemistry. 1995. Vol. 23, No. 7. P. 661-673.
- Walton D., Cusak M., Curry G.B. Implication of the amino acid composition of recent New Zeland brachiopods//Palaeontology. 1993.Vol. 36, No. 4. P. 883-896.
- Cusack M., Williams A. Evolutionary and diagenetic changes in the chemico-structure of the shell of cranioid brachiopods//Palaeontology. 2001.Vol. 44. No. 5. P. 875-903.
- Jope M. The protein of brachiopod shell-I. Amino acid composition and implied protein taxonomy//Comparative Biochemistry and Physiology. 1967. Vol. 20. P. 593-594.
- Jope M. The protein of brachiopod shell-II. Shell protein from fossil articulates: Amino acid composition//Comparative Biochemistry and Physiology. 1967. Vol. 20. P. 601-605.
- Дроздова Т.В. Геохимия аминокислот. М.: Наука, 1977. 200 с.
- Шанина С.Н., Бушнев Д.А. Определение энантиомеров аминокислот в природных органических соединениях//II Российское совещание по органической минералогии: материалы докладов. Петрозаводск: ИГ КарНЦ, 2005. С. 166-167.
- Кулик Н.А. Источники «Пымва-шор»//Изв. Арханг. общ. научн. русск. Севера. 1909. № 12.
- Чернов А.А. Сероводородные источники некоторых новых районов Печорского края, имеющие признаки нефтеносности и газоносности//Природные газы, 1932. Вып. 3.
- Митюшева Т.П., Патова Е.Н., Стенина А.С. и др. Биоминералообразование на сероводородных источниках гряды Чернышева//Минералогия и жизнь: происхождение биосферы и коэволюция минерального и биологического миров, биоминералогия: Материалы IV Международного минералогического семинара. Сыктывкар: Ин-т геологии, 2007. С. 119-120.
