An unusual burial of a Xiongnu-Sarmatian era warrior in the Western Siberian forest-steppe: results of a paleogenetic analysis

Author: Pilipenko A.S., Cherdantsev S.V., Trapezov R.O., Pozdnyakov D.V., Chikisheva T.A., Molodin V.I.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Ethnography
Article in issue: 4 т.46, 2018.
Free access
Short address: https://sciup.org/145145391
IDR: 145145391 | DOI: 10.17746/1563-0110.2018.46.4.123-131
Text of the article An unusual burial of a Xiongnu-Sarmatian era warrior in the Western Siberian forest-steppe: results of a paleogenetic analysis
Fig. 1. Location of the Sopka-2 site in the Baraba forest-steppe.
fell in the Xiongnu-Sarmatian era (i.e. last centuries BC to the first half of the I millennium AD). Large-scale migrations of human populations were accompanied by the establishment of new cultural and genetic connections. Those migrations were not limited to steppe only, but involved vast neighboring areas, including the foreststeppe zone of Western Siberia (for instance, Baraba forest-steppe at the interfluve of the Ob and Irtysh rivers). However, archaeological sites directly relating to the steppe nomads of the Xiongnu-Sarmatian time are rarely found in the south of the West Siberian Plain. The warrior burial (No. 688) at the chronologically diverse funerary and ritual site of Sopka-2 is one of the most outstanding archaeological complexes of that type. The site is located in the Vengerovsky District of the Novosibirsk Region (Fig. 1). On the basis of grave goods, the burial was dated to the late phase of the Xiongnu-Sarmatian period (4th to 5th centuries AD) (Molodin, Chikisheva, 1990), which was later confirmed by radiocarbon dating (described in detail below). The abundance and good preservation of the weapons found in the burial clearly point to the military status of the deceased.
In terms of grave goods and funerary rite, as well as the anthropological traits of the skeleton, burial 688 from Sopka-2 is clearly different from the Xiongnu-Sarmatian burials typical of Baraba (Molodin, Elagin, 1990), which unambiguously suggests a foreign origin for the individual. But a final decision regarding the ancestry of the warrior from Sopka-2 requires a more detailed study of various elements of the burial complex. Some features of the burial rite and grave goods find a wide spectrum of analogs in sites of both Eastern and Western regions of the Eurasian steppe belt (Molodin, 1995). The cranial morphology of the individual places him in the Central Asian anthropological type of Mongoloids, and points towards his possible origin from southern part of Eastern Siberia (Molodin, Chikisheva, 1990).
This study outlines the results of a molecular genetic analysis of a sample from the skeleton from burial 688 at Sopka-2, and provides a discussion of the findings in the light of previous archaeological (Ibid.; Molodin, 1995) and anthropological (Molodin, Chikisheva, 1990) studies of this burial complex.
Material and methods
Brief archaeological and anthropological description of the burial. Burial 688, belonging to the complex of diachronic archaeological sites commonly referred to as Sopka-2 (Vengerovsky District, Novosibirsk Region, junction of the Om and Tartas rivers), was excavated in 1989 by V.I. Molodin. In this burial, the remains of an adult male, 35–40 years old, were found (all anthropological descriptions were made by T.A. Chikisheva). The human remains and artifacts from the burial are stored in the Institute of Archaeology and Ethnography of SB RAS (Novosibirsk). As a thorough description of the burial complex was published earlier (Molodin, Chikisheva, 1990; Molodin, 1995); only significant information important for the interpretation of the genetic data will be provided.
The burial is flat (Fig. 2). The burial pit is oval, oriented NNE-SSW. Equine bones were found in the eastern part of the grave (in the filling). The deceased was buried in an extended supine position, with his head oriented NNE. Rich accompanying materials were found in the grave (Fig. 3–5). Almost all items, except a bone two-holed cheek-piece (see Fig. 4, 9 ), originate from territories outside of the Western Siberian forest-steppe. An iron broadsword (see Fig. 3, 1 ) with a composite pommel was found between the skeleton and western wall of the grave (see Fig. 4, 1–4 ; 5). The sword had been inhumed in its sheath, of which a massive silver onlay and pieces of wood with traces of red dye were preserved in the grave (see Fig. 3, 2 ). The broadsword was attached to the belt by a nephrite brace (see Fig. 4, 10 ). Elements of the belt-set (see Fig. 4, 5–7) show analogs with the finds from sites in the western part of the Eurasian steppe belt. Judging by these elements, the burial complex can be reliably dated to the 4th to 5th centuries AD, i.e. the
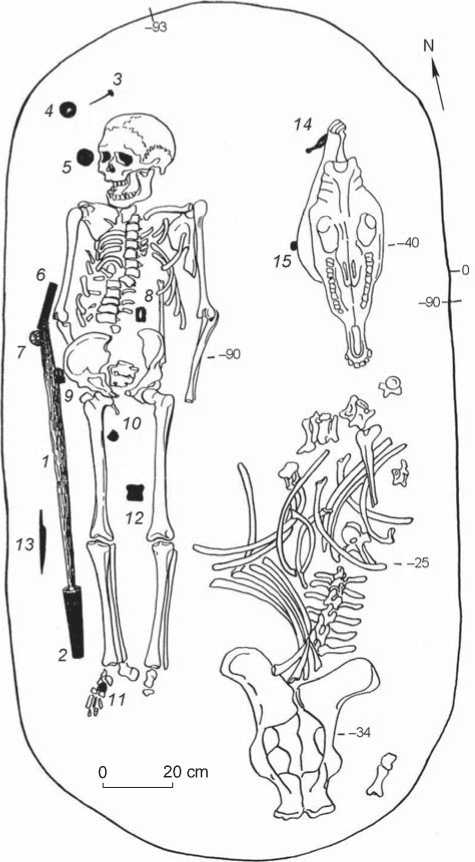
Fig. 2. Plan of burial 688 at Sopka-2.
1 – iron broadsword; 2 – silver tip of sheath; 3 – bronze nail; 4 – nephrite disc; 5 – gold pommel; 6 – nephrite brace; 7 – rock-crystal pommel; 8 – iron clip; 9 – silver buckle; 10, 11 – silver belt tips; 12 – equine pastern;
13 – iron knife; 14 – bone cheek-piece; 15 – bone clasp.
late Xiongnu-Sarmatian period. This is confirmed by a radiocarbon dating of a dental sample from the skeleton: ca 402–430 (1σ) and 393–474 (2σ) AD (lab index UGAMS23100/1214). In the burial were found also: an iron knife (see Fig. 4, 11 ), with a wide range of analogs, a bone clasp (see Fig. 4, 12 ), and an equine pastern (see Fig. 4, 13 ), placed between the legs of the buried man.
Summarizing the spectrum of analogs for burial 688 and its grave goods, it comes out that the complex exhibits only a few parallels with the Xiongnu burials of Central Asia: an almost northern orientation of the burial pit and the deceased, and the presence of the nephrite swordbelt brace. But the main features of the burial are more typical of the west of the Eurasian steppe: in other words, of the Sarmato-Alanian world. It is of note, however, that an analogy for the broadsword (specifically, its composite handle) is found in the Tugozvonovo burial, in the lowland Altai, West Siberian Plain (Umansky, 1978).
On the basis of cranial morphology, the individual from burial 688 can be assigned to the Mongoloid Central Asian anthropological type (Molodin, Chikisheva, 1990), which is widespread in more eastern regions of Siberia and Central Asia. This type is not common for the Early Iron Age population of Western Siberia and Altai. The skull from burial 688 displays the closest similarity, if not identity, to the cranial sample from the Xiongnu-Sarmatian cemetery of Kokel, in the Tyva Republic (Ibid.). Thus, the morphological features of the individual suggest his origin from a population clearly different from the majority of the inhabitants of the Western Siberian forest-steppe of the Xiongnu-Sarmatian era. They rather point towards a possible connection with the populations of eastern parts of Southern Siberia and Central Asia.

0 6 cm 0 2 cm
Fig. 3. Iron broadsword ( 1 ) and silver tip of sheath ( 2 ).

Fig. 4. Grave goods.
1–4 – details of the broadsword’s pommel; 5–7 – silver belt-set; 8 – iron clip; 9 – bone cheek-piece; 10 – nephrite brace; 11 – iron knife; 12 – bone clasp; 13 – equine pastern.
Bone sample used for the molecular genetic analysis. The macroscopically determined state of preservation of the skeletal remains was good, which indirectly suggested potentially good preservation of the DNA. Fragments of the left femur were sampled for the paleogenetic study.
Preliminary treatment of the skeletal samples and DNA extraction. All molecular genetics research were carried out in the paleogenetic laboratory of the Interinstitutional Center of Molecular Paleogenetics (Institute of Cytology and Genetics of the SB RAS, and Institute of Archaeology and Ethnography of the SB RAS, Novosibirsk, Russia). The protocol of this study is described in our previous publications (Pilipenko et al., 2015; Pilipenko, Cherdantsev, Trapezov et al., 2018). In order to eliminate contamination from modern DNA, the external surface of the sample was treated by 5 % sodium hypochlorite and then irradiated with UV. The external bone layer (~1–2 mm thick) was mechanically removed. Fine bone powder was then drilled out from the cortical layer. Then, the powder was incubated in 5M guanidine thiocyanate buffer at 65 ºC, while being constantly mixed during incubation. DNA extraction was performed using a phenol/chloroform protocol with subsequent sedimentation with isopropanol.
Genetic marker analysis. As the main informative loci, this study employed two systems of genetic markers: mtDNA (HVR I region and the appropriate single nucleotide polymorphism in the coding region of mtDNA) and 17 STR-loci of the Y-chromosome. As an additional locus, the polymorphic fragment of amelogenin gene (sex marker) and a set of nine autosomal STR-loci were analyzed. Genotyping protocols for all marker systems are described below.
Amplification of the mtDNA HVR I region was performed using two different protocols: four short overlapping fragments in one round of PCR (Haak et al., 2005), and one long fragment by nested PCR (consisted of two reaction rounds) (Pilipenko et al., 2008). DNA sequencing was carried out with an ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, USA). Sequencing extracts were analyzed with an ABI Prism 3100XL Genetic Analyzer automatic capillary sequencer (Applied Biosystems, USA) at the SB RAS Genomics Core Facility . Amplification of the fragments of interest of the mtDNA coding region was performed using oligonucleotide primers described in (Wilde et al., 2014). The phylogenetic position of the obtained mtDNA variant was determined based on the existing classification of mtDNA structural variants ( (van Oven, Kayser, 2009).
Profiles of 17 STR-markers of the Y-chromosome were determined using a commercial AmpFlSTR® Y-filer® PCR Amplification Kit (Applied Biosystems, USA), following the manufacturer’s protocol. Haplogroups of the

Fig. 5. Pommel of the broadsword.
STR haplotypes of the Y-chromosome were determined using two programs: Whit Athey’s Haplogroup predictor and Vadim Yurasin’s YPredictor 1.5.0 .
Profiling of nine autosomal STR-loci and analysis of the amelogenin gene region polymorphism (sex marker) was performed using commercial AmpFlSTR® Profiler® Plus PCR Amplification Kit (Applied Biosystems, USA), following the manufacturer’s protocol.
Measures against contamination and verifi cation of the results. All procedures with the skeletal specimens were carried out in a specially-equipped laboratory for molecular paleogenetics (Institute of Cytology and Genetics of the SB RAS, and Institute of Archaeology and Ethnography of the SB RAS, Novosibirsk, Russia). A description of the anti-contamination measures and verification of the results can be found in our previous publication (Pilipenko, Trapezov, Polosmak, 2015). Employment of those measures and consistency of the obtained results ensure the reliability of our experimental data.
Results and discussion
As it was previously shown by our studies, the skeletal specimens from archaeological sites of the Baraba foreststeppe, which display a high degree of macroscopic preservation, contain authentic DNA suitable for both mtDNA and nuclear loci analysis (Pilipenko et al., 2017). The results of the present study confirm this conclusion. Using three extracts of total DNA from the femoral sample of the individual from Sopka-2, burial 688, we obtained reproducible data on mtDNA and nuclear loci structure (polymorphic region of the amelogenin gene, STR-loci of autosomes and the Y-chromosome).
The HVR I region of the control region of DNA in position 15996–16403 contained five nucleotide substitutions relative to the Cambridge Reference mtDNA Sequence and displayed haplotype 16223T-16239T-16243C-16319A-16362C. Such a haplotype structure unequivocally suggests that the studied mtDNA sample belongs to East Eurasian haplogroup D. This is also in accordance with the status of position 5178A in the coding region of mtDNA, which is specific for this haplogroup. The presence of a G16319A substitution points towards the belonging of the line under study to subgroup D4b1. We believe that variants with additional substitutions C16239T and T16243C constitute a separate subcluster of this subgroup, though it has not been annotated as a part of the united mtDNA phylogenetic tree so far (van Oven, Kayser, 2009). It is established that the main stages of the diversification of variants of haplogroup D4b1 have taken place in populations of Central and Southern Siberia, as well as in East Asia (Derenko et al., 2010).
The presence of both allele variants of the amelogenin gene (i.e. associated with both X- and Y-chromosomes) detected in this study clearly confirms that the individual was male, which is in agreement with his skeletal morphology. The high level of DNA preservation in the remains permitted a reconstruction of the complete allele profile of the Y-chromosome STR-loci:
|
DYS456 |
16 |
|
DYS389I |
13 |
|
DYS390 |
24 |
|
DYS389II |
28 |
|
DYS458 |
18 |
|
DYS19 |
13 |
|
DYS385a/b |
13/16 |
|
DYS393 |
13 |
|
DYS391 |
10 |
|
DYS439 |
12 |
|
DYS635 |
24 |
|
DYS392 |
16 |
|
Y GATA H4 |
11 |
|
DYS437 |
14 |
|
DYS438 |
11 |
|
DYS448 |
23 |
|
Predicted haplogroup |
Q (Q1a) |
On the basis of these data, the predictor software has unequivocally shown that this variant of the Y-chromosome belongs to subgroup Q1a of haplogroup Q. This subgroup, in the Eurasian context, is more typical of East Eurasian populations. The most probable place of formation of the Y-chromosome haplogroup Q (and likely its subcluster Q1a) are Central Asia and Southern Siberia (Huang et al., 2018).
Thus, in general, our phylogenetic analysis has demonstrated that both the mtDNA and the Y-chromosome of the male from burial 688 at Sopka-2 belong to clusters that emerged and dispersed in the gene-pools of populations from the regions of Siberia and Central Asia lying to the east of the Baraba forest-steppe. So, this individual likely had eastern ancestry. This conclusion coincides with the results of the craniological survey suggesting his possible origin eastward.
The analysis of the distribution of the Y-chromosome and mtDNA variants in ancient and modern Eurasian populations is an effective tool for potential specification of the vectors of genetic relatedness of the studied individual. Owing to the presence of large reference datasets, such a phylogenetic comparison is typically carried out using the data on modern populations. Phylogeographic analysis has shown that the variant of mtDNA with haplotype 16223T-16239T-16243C-16319A-16362C of haplogroup D4b1 is a rare component of the Eurasian gene-pool. Interestingly, this variant is found in eastern (relative to the Western Siberian foreststeppe) modern populations, like the Tuvans or Kalmyks of historically Central Asian origin (Derenko et al., 2007), and also in western ones, i.e. in the Bashkirs from the Volga-Ural region (Bermisheva et al., 2002). The same applies to the STR allele profile of the Y-chromosome belonging to haplogroup Q (Q1a): individuals with identical or similar profiles are present in the east of Eurasia (e.g. in China) but also in Eastern Europe (e.g. in the Poles).
Thus, while the analysis of the uniparental markers at the level of phylogenetic clusters revealed the eastern origins of the individual, the phylogeographic study of the exact structural variants of the Y-chromosome and mtDNA did not completely clarify his position among modern Eurasian populations and, hence, his geographical ancestry. This apparent contradiction can be resolved if the historical and archaeological context is taken into account. Also, it is important to incorporate data on ancient Eurasian populations into our phylogeographic analysis.
As has been mentioned already, the burial is archaeologically dated to the 4th to 5th centuries AD, while the age of haplogroup D4b1 is estimated from 23.9 to 31.5 thousand of years. The minimum ages of its subclusters vary substantially, but even for the youngest it is not less than 2500 years, i.e. all of them are substantially older than the burial complex 688 of Sopka-2 (Derenko et al., 2010). This means that the diversification of the mtDNA cluster D4b1 (in more eastern regions of Eurasia, according to phylogenetic data) was taking place well before the lifetime of the studied individual. The same is true for the Y-chromosome haplogroup Q1a. Consequently, the conclusion regarding the eastern origins of the Y-chromosome and mtDNA clusters, detected in the Sopka-2 individual, applies to his deep genetic roots, which most probably connect him with southern regions of Siberia and Central Asia. This result is in perfect agreement with anthropological data (Molodin, Chikisheva, 1990).
After the clusters emerged, their pattern of dispersal in Eurasia might have been substantially changed. Before the Sopka-2 individual was buried, there had been a thousand years (counting from the middle of the 1st millennium BC) of intense migration in the Eurasian steppe, involving neighboring forest-steppe areas. This migratory period included the Scythian and Xiongnu-Sarmatian times, and it was characterized by long-range movements of different populations (and their genes) in various directions (see, e.g., (Botalov, 2000: 207–233)). Numerous groups of early nomads were the main driving force of those movements. One of the key directions was the dispersal of eastern nomads’ genes into western regions of the Eurasian steppe. This is confirmed by mtDNA data for both Scythian (Unterländer et al., 2017; Pilipenko, Trapezov, Cherdantsev et al., 2018) and Xiongnu-Sarmatian (Pilipenko, Cherdantsev, Trapezov et al., 2018) periods. Thus, at the final stage of the Xiongnu-Sarmatian period (i.e. the time of burial 688 at Sopka-2), East Eurasian variants of the Y-chromosome and mtDNA might well be widespread throughout the whole Eurasian steppe belt. Nevertheless, their frequency must have been higher in the eastern than the western part of the region. Owing to the extremely high mobility of nomadic populations, relocation of genetic, as well as archaeological, material could often have been similar to Brownian motion (Botalov, 2000: 285, fig. 19).
The above-mentioned difficulties relating to localizing the origins of a migrant based on his genetic features can be illustrated if data on ancient Eurasian populations are included in phylogeographic analysis. The mtDNA variant with haplotype 16223T-16239T-16243C-16319A-16362C of haplogroup D4b1, among ancient Siberian populations studied to date, was found in two individuals of the Pazyryk culture of Scythian times in the Altai Mountains (Unterländer et al., 2017) and in an individual from Stepushka-1 cemetery of the Bulan-Koba culture in the Altai piedmont (authors’ unpublished data). This directly confirms the presence of the variant in the gene-pool of the population from neighboring regions of Southern Siberia of the Scythian and Xiongnu-Sarmatian times. Unfortunately, however, ancient population of Tyva (including Kokel cemetery, where individuals very similar to Sopka-2 in terms of cranial morphology were studied) still remains unexplored by paleogenetic methods. On the other hand, this variant of mtDNA was detected in the gene-pool of the classic Scythians from the Northern Pontic area (!) (Der Sarkissian, 2011). Clearly, it was dispersed far to the west of the Eurasian steppe belt in the times before the studied individual was buried.
It is difficult at the present time to carry out a similar phylogeographic analysis of the variant of haplogroup Q (Q1a) detected in this study. First, there are not enough published data on the Y-chromosome variation in ancient Eurasian populations. Second, we do not have data regarding the status of phylogenetically informative singlenucleotide polymorphisms (SNP) of the Y-chromosome, this being an aim for future research. At the moment, it is only possible to conclude that individuals carrying various subclusters of haplogroup Q1a were reliably detected in ancient nomadic populations of the Xiongnu-Sarmatian period in Northern China (Zhao et al., 2010) and the Altai Mountains (Allentoft et al., 2015).
Summing up, there is no doubt that the deep genetic roots of the male from burial 688 lead eastwards. But if the results of the phylogeographic analysis are taken into account, it becomes problematic to conclude firmly whether the individual arrived in the Western Siberian forest-steppe from the western or from the eastern part of the “steppe belt”: existing paleogenetic data are not enough for localization of his immediate ancestors. There is some evidence for his probable migration from the East: the presence of similar (and also identical) variants of mtDNA haplogroup D4b1 and the Y-chromosome haplogroup Q1a in individuals of the Scythian and Xiongnu-Sarmatian periods in adjacent eastern regions of Southern Siberia (the Altai Mountains); and the concordance of the Y-chromosome and mtDNA variants of the individuals in terms of their East Eurasian origin. But on the other hand, the prevalence of “eastern” genetic lineages in the Late Xiongnu-Sarmatian times makes plausible other scenarios that can explain the apparent contradiction between the anthropological and genetic data suggesting the eastern origin of the Spoka-2 individual, and western analogies of the grave goods of his burial. For instance, a descendant of people who had migrated from the eastern to the western part of the Eurasian steppe belt in preceding centuries (and thus possessing East Eurasian genetic features), might himself have arrived in the Baraba forest-steppe from western steppe areas. As such, he would have been a carrier of other, Sarmato-Alanian, culture and traditions, which was expressed in the inventory of the burial. So, the archaeological pattern of the complex can point to the region where the individual arrived from with a high precision.
The dispersal of genetic components emerging as a result of contacts of remote populations makes it difficult to the determine geographic origin of a single migrant (or a few migrants) by paleogenetic methods. This limitation should be extended to physical anthropology methods as well. This is particularly true when it comes to the ancient nomadic population of the “steppe belt” and adjacent areas of Eurasia of the Early Iron Age and later periods. The use of representative population data (of course, if these are present) might be an effective way to overcome, at least partially, the limitations mentioned above. The resolution of genetic methods of studying single individuals (or small samples) can be increased if their more detailed genetic profile can be obtained. But even a whole-genome analysis of a single individual (or a few individuals) does not remove all limitations. A more general solution to this problem would be a comprehensive investigation of archaeological and skeletal data, employing a wide range of methods and approaches. Full-scale use of the whole arsenal of natural science methods (biological, chemical, physical) must be accompanied by necessarily correct accounting for the archaeological (historical) context of the burial. This would help to overcome the methodological limitations of each of the approaches when interpreting their results, and receive the most objective knowledge regarding the phenomenon under study. This, in turn, would get us as close as possible to reconstruction of one or another historical event.
Acknowledgements
This study was supported by the Russian Science Foundation (Project No. 17-78-20093). The use of paleogenetic facilities of the Institute of Cytology and Genetics of the SB RAS was covered from the funds of the public contract for Project No. 0324-2018-0016.
