An Upper Second Molar from Akhshtyrskaya Cave and Population Affinities of the Middle Paleolithic Hominins of the Caucasus

Автор: Zubova A.V., Moiseyev V.G., Kulkov A.M., Kulakov S.A.
Журнал: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Рубрика: Anthropology and paleogenetics
Статья в выпуске: 2 т.53, 2025 года.
Бесплатный доступ
The Akhshtyrskaya cave site in northwestern Caucasus dates mostly to the Middle Paleolithic. In 1961, during the analysis of faunal assemblage from layer 3a, a permanent upper second molar of Homo (AKH 1) was discovered. Initially, it was attributed to fossil Homo sapiens. Here, we present new findings related to this specimen, specifically to its external and internal morphology, amount of enamel and dentine in the lateral part of its crown, and a comparison of metric and nonmetric characteristics with those of Denisovans, H. ergaster/erectus/georgicus, H. erectus, H. antecessor, H. neanderthalensis, H. sapiens, Middle Pleistocene Homo of Europe, and Middle and Late Pleistocene Homo of China, distinct from H. erectus. At the first stage, frequencies of nonmetric traits were subjected to principal component analysis. At the second stage, obtained PCA scores were integrated with measurements of the same specimens using PC analysis with grouping variable. Results suggest that the molar from Akhshtyrskaya by no means belongs to H. sapiens. Its morphology is very archaic, linking it to mostly Middle Pleistocene hominin teeth from China. The volume of lateral dentine opposes AKH 1 to molars of Late Pleistocene Homo of Europe and those of H. sapiens, revealing a shift toward Denisovans. The specimen likely belongs to an evolutionary line that has thus far not been found in Europe. Probably, it is of Asian origin.
Middle Paleolithic, Caucasus, Neanderthals, Denisovans, microCT, dental anthropology
Короткий адрес: https://sciup.org/145147500
IDR: 145147500 | DOI: 10.17746/1563-0110.2025.53.2.134-143
Текст научной статьи An Upper Second Molar from Akhshtyrskaya Cave and Population Affinities of the Middle Paleolithic Hominins of the Caucasus
The Caucasian region has always, since the most ancient times, been an outpost for human colonization of Eurasia. During the whole history of humanity, it has served as a corridor connecting Europe, Near East, and Central Asia (Lyubin, 1997: 11; Ghasidian et al., 2023). The position of the Caucasus at the crossroad of several migratory passageways provided a possibility of admixture between various groups of ancient hominins. At the same time, the complex mountainous terrain has always been and still is a natural factor of forming refugees and isolating some human populations. Thus, the Caucasus can be
Fig. 1. Location of the Akhshtyrskaya Cave site.
Hajdinjak et al., 2018; King et al., 2016; Pinhasi et al., 2012; Martinón-Torres et al., 2008). Only the Neanderthal fossils from Mezmayskaya Cave in the Northwest Caucasus were preserved enough to study not only their taxonomy but the population affinities as well (Hajdinjak et al., 2018).
This article outlines the results of our study of an upper human molar (hereinafter, AKH 1) found in the Akhshtyrskaya cave site in the Northwest Caucasus (43°31′ N, 39°59′ E) (Kulakov, Baryshnikov, Levkovskaya, 2007).
The first attempt to determine if the tooth belonged to a Neanderthal or a H. sapiens individual was made in the 1960s, but the limitations of the dental anthropology methods available at that time did not allow solving this question. It was concluded that the tooth belonged to Homo sapiens fossilis (Vekilova, Zubov, 1972: 62). Nevertheless, the preliminary results of our studies of AKH 1 morphology (Zubova, Moiseyev, Khartanovich, 2017; Zubova et al., 2018) made this suggestion doubtful.
In the present study, a detailed analysis of external and internal AKH 1 morphology was carried out, and a comparison of the AKH 1 dental characteristics with various Eurasian Middle and Late Pleistocene populations was performed.
Material and methods
Archaeological context. The Akhshtyrskaya site is located in the Adlersky District of Sochi, in Bolshaya Kazachebrodskaya Cave, at the left bank of the Mzymta River (Fig. 1). This karst cavity belongs to “corrosion-erosion caves—ponors” and has two entrances: the southern lies 100–102 m above the river level and the eastern 95–97 m. The total length of the cave is more than 150 m, while its width varies from 7 m at the entrance to 2 m in the narrowest points of the passages. The thickness of Pleistocene deposits reaches 3 m and more (Kulakov, 2015).
The cave was discovered in 1936 by S.N. Zamyatnin and M.Z. Panichkina and then excavated by the same specialists in 1937–1938, and by E.A. Vekilova and M.Z. Panichkina in 1961 (Zamyatnin, 1961: 104–117; Vekilova et al., 1978). The study of the cave was resumed in 1996–2008 by the Black Sea expedition of the Institute for the History of Material Culture RAS. The site has 9 stratigraphic layers. The lower three (7–9) are archaeologically sterile. Layers 3–6 contain Middle Paleolithic stone assemblages. The upper part of layer 3 (3/1, previously 3a) reveals a mixture of Upper and Middle Paleolithic finds (Kulakov, 2005). The AKH 1 molar was discovered in the faunal sample of layer 3/1. The layer was dated to 35,000 ± 2000 BP (Cherdyntsev et al., 1965), but the reliability of this date is doubtful.
AKH 1 molar (Fig. 2). The specimen is a well-preserved upper right second molar of an adult individual. Post-mortem damage of crown enamel is minor; ante-mortem attrition corresponds to grade 4 on Molnar’s scale (Molnar, 1971). The vestibular roots were broken and restored in the 1960s. A small enamel fragment from the cervical area (where the root was broken) was lost. Though the external enamel pattern is worn, the position of the main fissures and presence of several accessory grooves (reaching the surface of the crista oblique in the central fovea area) could be determined.
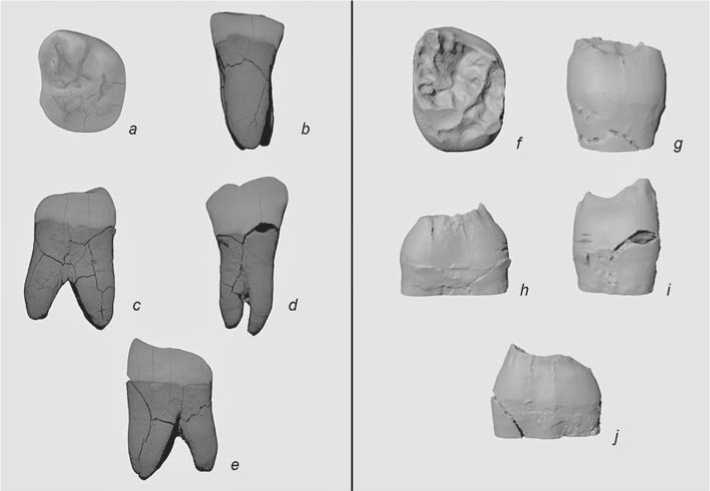
Fig. 2. 3D-model of the AKH 1 molar based on microtomographic images.
a–e – external morphology; f–j – internal morphology. a , f – occlusal plane; b , g – lingual plane; c , h – distal plane; d , i – vestibular plane; e , j – mesial plane.
Methods. External AKH 1 morphology was described following standard protocols (Zubov, 2006; Turner, Nichol, Scott, 1991; Bailey, 2002; Martinón-Torres et al., 2012). The following traits were scored: number of main cusps, relative size and shape of the metacone and hypocone, presence or absence of the parastyle, positions of the main intertubercular fissures, number and shape of the roots. The mesio-distal and buccolingual diameters of the crown were measured. Other traits could not be scored owing to the enamel wear.
The enamel-dentin junction (EDJ) was reconstructed after micro-CT scanning, to score the following traits: presence or absence of the anterior and posterior fovea, plagyocristids, epicrists, C5, postparaconule, accessory mesial cusps, Carabelli’s cusp, central shift and inclination of the apexes of the dentin horns at the main crown cusps, odontoglyphic pattern.
The volumes of enamel and dentin of the lateral part of the crown were measured in CTAn (Bruker-micro CT). According to the protocols (Olejniczak, 2006: 125–128, Martín-Francés et al., 2018: 3, 7) measurements were taken in the area between the cervical plane, i.e., the plane in halfway between the most apical continuous ring of enamel and the plane containing the last hint of enamel, and the occlusal basin plane, which is parallel to the cervical plane and tangent to the lowest enamel point of the occlusal basin.
Micro-CT scanning was carried out at the Research Centre for X-ray Diffraction Studies of the Saint-Petersburg State University, employing a NEOSCAN 80 X-ray scanner with the following settings: tube voltage 101 kV, current 40 μA, with an aluminum filter. Spiral scanning was performed with a rotation step of 0.25° and averaging 4 frames. Resolution: 6 μm. The reconstruction of the raw data was carried out in NRecon (Bruker-microCT). The 3D-model used for the virtual separation between enamel and dentin was built in CTAn (Bruker-microCT) and visualized in CTvox (Bruker-microCT).
The morphological pattern of АКН 1 was compared with upper second molars of Denisovans (DEN), Homo ergaster/erectus/georgicus (HE/E) from Dmanisi, H. erectus (HE), H. antecessor (HA), H. neanderthalensis (NEA), early and modern H. sapiens (HS), Middle Pleistocene European Homo (EMPH), non- H. erectus Middle and Late Pleistocene Homo from China (EAMLPH), and MC 8 specimen from Manot Cave (probably a hybrid individual).
The dentin and enamel volumes were compared using box-and-whiskers diagrams, depicting the mean values and ranges of variation (standard deviation, standard error of the mean) for both variables.
The individual non-metric data were compared via principal components analysis (PCA) in Statistica for Windows, 8.0 (module “Multivariate Exploratory Techniques”), based on trapezoid shape of the crown,
Table 1. Reference sample of upper second molars
|
Taxon |
Fossils |
Publication |
|
H. ergaster/erectus/georgicus |
Dmanisi (2700, 2282) |
(Martinón-Torres et al., 2008; Gabunia, Vekua, Lordkipanidze, 2001) |
|
Homo antecessor |
Atapuerca, Gran Dolina (TD6 12, TD6 69) |
(Bermúdez de Castro et al., 1999; Martín-Francés et al., 2018) |
|
Homo erectus |
Zhoukoudian (40, 41); Yiyuan, Hexian (PA 833, PA 837) |
(Weidenreich, 1937; Xing et al., 2015, 2016) |
|
East Asian Middle and Late Pleistocene Homo |
Hualongdong 6, Chaoxian, Xujiayao (PA 11480-6), Diushan 1 |
(Bailey, Liu, 2010; Liao et al., 2019; Wu et al., 2019; Xing et al., 2015) |
|
European Middle Pleistocene Homo |
Atapuerca, Sima de los Huesos (SH) (AT-46, AT-960, AT-824, AT-2179, AT-407, AT-4319, AT-12, AT-2175, AT-815, AT-588, AT-817, AT-821, AT-15, AT-170), Visogliano (Vis 3), Biache-Saint-Vaast |
(Martinón-Torres et al., 2012; Martín-Francés et al., 2018, 2020, 2022; Zanolli et al., 2018) |
|
Neanderthals |
Rozhok I, Chagyrskaya Cave (CH1141, CH10, CH57), Leuca, Le Moustier, Moula-Guercy (M-G2-117 M-F3-215), Krapina (F/H MX M, KRD98, D96, D165, D166, D169), Stajnia (S5000) |
(Benazzi, Bailey, Mallegni, 2013; Hlusko et al., 2013; Martín-Francés et al., 2018; Picin et al., 2020; Zubova et al., 2022) |
|
Denisovans |
Denisova Cave (Denisova 4, 8), Harbin |
(Zubova, Chikisheva, Shunkov, 2017; Ni et al., 2021) |
|
Homo sapiens |
Daoxian (PA 1554, PA 1556, PA 1587, PA 1592), Qafzeh 11, Sungir 2, 3, Kostyonki 18, Cherepakha 13 (MAE NVF 95-2, 95-18, 95-22, 95-27), modern Papuans (MAE 212-11, 20), Bosco-Pontini 5, 6, 9, Pigorini |
(Gromov, Zubova, Moiseyev, 2017; Khaldeeva, Kharlamova, Zubov, 2010; Liu et al., 2015; Martín-Francés et al., 2018; Zubova et al., 2022). Unpublished data of A.V. Zubova and V.G. Moiseyev |
|
Unknown |
Manot, МС 8 |
(Sarig et al., 2021) |
Carabelli’s trait (grades 2–5), accessory distal cusp, hypocone reduction (grades 3, 3+), posterior fovea, parastyle, epicrista, plagyocrista, odontoglyphic grooves of the third order. All the traits were scored in a binary form: presence or absence.
To reconstruct the taxonomic position and biological affinities of the AKH 1 molar, the nonmetric and metric (crown diameters) data were integrated in the same analysis. This was also carried out using PCA but employing the following set of variables: PC1–4 coordinates of individual fossils from the non-metric analysis (see above), mesio-distal and buccolingual diameters of the crown. The PCA with a grouping variable (Statistica 8.0, module “PLS/ PCA Multivariate/Batch SPC”) was used. The name of the groups and samples included are listed in Table 1.
Results
Morphology of AKH 1. The crown (Fig. 2) is large: mesio-distal diameter 11.2 mm, buccolingual diameter 13 mm. It exhibits a trapezoid shape. Its vestibular margin is oblique due to the reduction of the metacone, while the hypocone is moderately reduced (4-). On the external enamel surface the following features are present: posterior fovea, a deep fissure separating the metacone and paracone, and several small transverse grooves reaching the surface of the continuous crista oblique.
The mesial and distal marginal ridges of occlusal surface at EDJ run close to the margins of the crown. The basal part of the crown is only subtly wider than the occlusal one. The hypocone is connected with the protocone and metacone by continuous ridges. The marginal ridges do not cross the intertubercular fissures. There are small dentin protrusions on the distal marginal ridge marking the presence of two accessory cusps (C5). Two segments marking the presence of accessory mesial cusps branch from the mesial marginal ridge towards the central fovea. The transverse crest (epicrista) is interrupted by an intertubercular fissure.
A Carabelli’s cusp was detected in the protocone. The cusp reaches grade 2 or larger: a more precise definition is not possible due to attrition. A deep triangular talon basin present in the distal area. The protocone and metacone are connected by a continuous crista oblique. The apex of the dentin horn of the metacone is slightly inclined inwards. The degree of inclination of the other cusps’ apexes cannot be determined due to attrition. A postparaconule is present in the paracone.
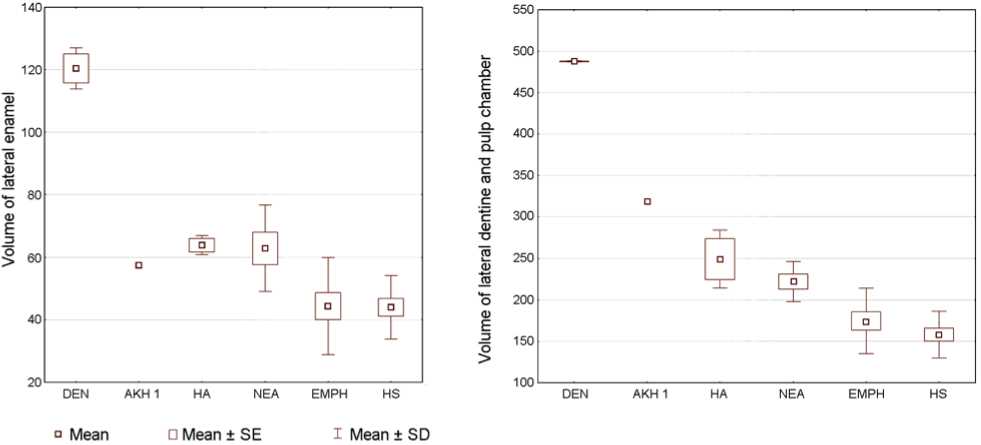
Fig. 3. Enamel and dentin volumes of AKH 1 and in the reference samples.
DEN – Denisovans; HA – Homo antecessor ; NEA – Neanderthals; EMPH – European Middle Pleistocene Homo ; HS – H. sapiens .
о
о о
-2
АКН 1 .
Manot о □
DEN4 “
□ Ж
ж
ж
о
-2
О iPC 1
□
О
° • ° л
° □ ®
• НА а НЕ/Е а НЕ О EMPH a NEA
*EAMLPH +DEN • HS oother
Fig. 4. Principal component analysis of the non-metric variables.
DEN – Denisovans; HA – Homo antecessor ; NEA – Neanderthals; EMPH – European Middle Pleistocene Homo ; HS – H. sapiens ; EALMPH – East Asian Middle and Late Pleistocene Homo ; HE – H. erectus ; HE/E – H. ergaster / erectus / georgicus .
The odontoglyphic pattern of AKH 1 is specific. All the fissures are very deep. The cusp fissures observed at EDJ form small “pools” around themselves, widening with distance from the intertubercular fissures. Such morphology was only observed in Chinese H. erectus so far (Bermudez de Castro et al., 2021: Tab. 4, fig. 11).
Proportions of enamel and dentin. The volumes of enamel and dentine of the lateral part of the AKH 1
crown can be compared with a limited number of samples (Fig. 3). Its enamel volume is average for a Middle Paleolithic, and falls into the transgression zone between Neanderthals and Middle Pleistocene Homo from Atapuerca (SH). This volume is larger than in H. sapiens and smaller than in Denisovans. The dentin volume is rather large: AKH 1 stands out of all the European samples, differing sharply from H. sapiens and shifting towards Denisovans.
PCA results. The results of the first stage of PCA reveal that the non-metric features of upper second molar effectively differentiate almost all the reference taxonomic groups (Fig. 4). Exclusion is EMPH, demonstrating an extremely wide range of variation. Along PC1, they transgress with Neanderthals, EAMLPH, H. antecessor and, mostly, with H. sapiens . The similarity with the individuals of the modern Homo is due to the high frequency of reduced hypocone in the EMPH sample (Table 2). The authors of the original publication of the Atapuerca (SH) fossils explained the reduction of the hypocone by homoplasy but not an affinity with the direct ancestors of H. sapiens (Arsuaga et al., 2014). Nevertheless, the presence of such homoplasy only in Atapuerca SH make this explanation doubtful.
All the samples are distributed in a predictable manner, although a small transgression zone between Neanderthals and the Chinese hominins can be seen in the center of the plot. PC 1 contrasts H. sapiens and EMPH, on the one hand, and Denisovans and the Chinese hominins, on the other hand (Fig. 4). The former exhibit an exceptionally high frequency
of the marked hypocone reduction, while the latter display the maximum frequencies of all the accessory cusps, fissures, and ridges of the crown except for the epicrista (Table 2). Neanderthals occupy an intermediate position in the plot.
PC 2 differentiates Denisovans and Neanderthals (negative values) from the other samples. The negative extreme of this axis is occupied by the Denisova 8 and Harbin individuals (Fig. 4) possessing parastyles but lacking the trapezoid shape of the crown. Neanderthals are characterized by moderate negative values owing to the absence of the parastyle, while the trapezoid shape of the crown is rare. The H sapiens are plotted around the zero. All the other specimens display positive values of the PC 2 (Fig. 4).
The AKH 1 molar falls into both PC1 and PC2 values typical of the Chinese Homo , while being substantially distant from H. sapiens and, to a lesser extent, from Neanderthals. The most similar fossils to AKH 1 are Denisova 4 and the upper second molar from Manot Cave in the Near East (Fig. 4). The upper second molars from Longtan (Hexian) and Yiyuan in China, as well as the single EMPH specimen (Visogliano 3), plot more distantly from AKH 1 and closer to the European fossils.
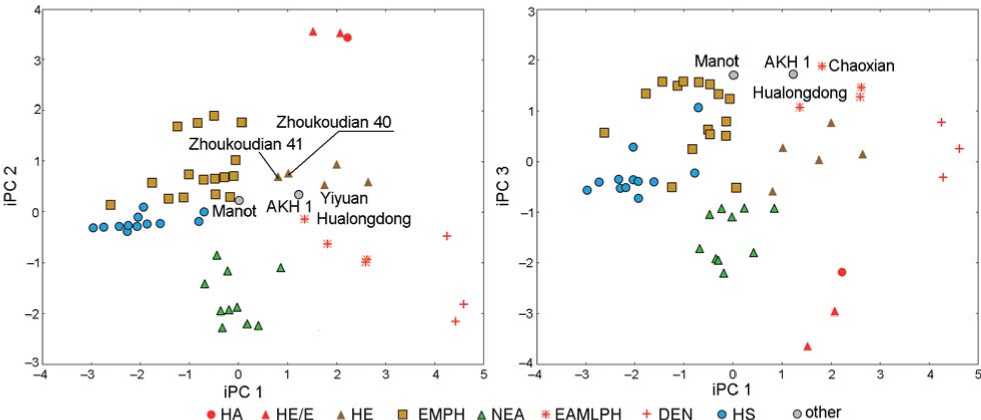
The PCA integrating metric and non-metric traits (iPCA) revealed similar picture of biological distances between the samples and individual fossils,
Table 2. PCA loadings of the non-metric variables
|
Trait |
PC 1 |
PC 2 |
PC 3 |
PC 4 |
|
Trapezoid crown shape |
0.37 |
0.74 |
0.05 |
0.09 |
|
Hypocone reduction |
0.80 |
–0.11 |
–0.04 |
0.05 |
|
Carabelli’s cusp |
–0.52 |
0.46 |
0.43 |
0.12 |
|
Accessory distal cusp |
–0.35 |
0.14 |
–0.44 |
0.78 |
|
Parastyle |
–0.34 |
–0.60 |
–0.42 |
–0.04 |
|
Transverse crest (epicrista) |
0.04 |
0.50 |
–0.56 |
–0.47 |
|
Posterior fovea |
–0.70 |
0.34 |
–0.28 |
–0.08 |
|
Continuous crista oblique |
–0.59 |
–0.14 |
0.41 |
–0.04 |
|
Accessory fissures |
–0.75 |
–0.05 |
0.00 |
–0.26 |
though the zones of inter-sample transgression became narrower, and the polarity of PC1 changed (Fig. 5). As in the previous analysis, the main trend was the differentiation between the H. sapiens and Asian fossils. This difference is related to both non-metric PC1 variation and the crown diameters very strongly correlated with iPC1 (Fig. 5; Table 3). Neanderthals occupy an intermediate position between European and Asian groups, but closer to EMPH and HS.
The second iPC differentiates Neanderthal, Denisovan, and, to an extent, EAMLPH from the other samples. The values of this vector are highly correlated with values of non-metric PC2. The similarly negative iPC2 values of Neanderthal and Denisovans correspond to their genetic affinities as sister groups. They are differentiated by iPC3, which has the highest loading from the non-metric PC4.
Zhoukoudian 40
0 1 2 3 4 S iPC 1
® HS о other
Yiyuan Hualongdong
ДДА д Ад
° Do p Manot n ■ □ °
О
AKH 1 *
Zhoukoudian 41
□ >B^
Manot AKH 1 -chaoxian
■ ° *
□ □□ Hualongdong +
□ 6 □
▲ *
дАдЛ A
-3 -2 -1 0 1 2 3 4 5 -4 -3 -2 -1
iPC 1 • HA a HE/E a HE □ EMPH a NEA + EAMLPH + DEN
Fig. 5. Integral principal component analysis.
Legend same as on Fig. 4.
Table 3. Integrative PCA loadings of the metric and non-metric variables
|
Variable |
iPC 1 |
iPC 2 |
iPC 3 |
|
Mesio-distal crown diameter |
0.92 |
0.04 |
–0.08 |
|
Buccolingual crown diameter |
0.82 |
0.03 |
0.22 |
|
Non-metric PC 1 |
–0.8 |
0.15 |
0.02 |
|
Non-metric PC 2 |
–0.03 |
0.81 |
0.11 |
|
Non-metric PC 3 |
–0.26 |
–0.32 |
0.22 |
|
Non-metric PC 4 |
–0.02 |
–0.16 |
0.75 |
|
Neanderthals |
–0.02 |
–0.59 |
–0.53 |
|
Homo sapiens |
–0.57 |
–0.08 |
–0.11 |
|
East Asian Middle and Late Pleistocene Homo |
0.46 |
–0.2 |
0.31 |
|
Homo erectus |
0.29 |
0.17 |
0.04 |
|
Denisovans |
0.46 |
–0.19 |
0.02 |
|
European Middle Pleistocene Homo |
–0.28 |
0.4 |
0.43 |
|
Homo antecessor |
0.17 |
0.34 |
–0.23 |
|
H. ergaster/erectus/georgicus |
0.09 |
0.04 |
0.18 |
|
AKH 1 |
–0.09 |
–0.17 |
0.12 |
|
Manot |
0 |
0.03 |
0.18 |
|
Eigenvalue |
3.23 |
1.82 |
1.64 |
|
% of total variation |
18.97 |
10.73 |
9.62 |
The AKH 1 molar falls into the range of the Chinese fossils and separates from either H. sapiens or Neanderthals in any iPCs combination (Fig. 5).
Discussion
The results of the present study have shown the presence of a number of archaic features in AKH 1 morphology, including: specific odontoglyphic pattern, talon and trygonid “basins”, large crown dimensions, and a very large volume of the lateral dentin. Thus, the hypothesis assigning the AKH 1 fossil to H. sapiens must be rejected. In the same time, the absence of specific “Neanderthal” markers prevents diagnosing the tooth as a typical Neanderthal. Both versions of PCA put AKH 1 to the range of Chinese fossils—much more ancient than the published date of layer 3/1 (3а) of the Akhshtyrskaya site.
We acknowledge that the taxonomic interpretation of the results based on only one tooth must be made very cautiously. But the correct distribution of the other fossil (including isolated teeth), with established taxonomic diagnoses, in the PCA plots suggests that the position of AKH 1 can also be considered reliable. Thus, the similarity of the molar to the Chinese samples is hardly only a statistical artifact. This finding suggests that the Middle Paleolithic population dynamics in the Caucasus were more complicated than it was previously thought. Besides “classic” Neanderthals, it might have been the home for other members of the Homo genus possessing archaic dental morphology similar to Chinese fossils. The taxonomic status of those hominins can hardly be precisely diagnosed at the moment, but various scenarios of their migration to the Caucasus can be discussed.
The first option is long-term preservation of the dental features typical of the earliest waves of hominin colonization of Eurasia in this mountainous region. It is feasible from the theoretic point of view, since during the last 2 millions years the Caucasus has been inhabited, besides H. sapiens and Neanderthals, by H. ergaster/erectus/georgicus and H. heidelbergensis (Martinón-Torres et al., 2008; King et al., 2016). For how long they retained in the region and how possible was their admixture with more advanced hominin groups is not clear.
Thus, the possibility of such long-term preservation of their dental pattern in the Middle Paleolithic Caucasian population cannot be completely excluded as well. But according to both distribution of all the variables and PCA results (see Fig. 4, 5) AKH 1 differs from the upper second molars from Dmanisi as well as from the H. antecessor and EMPH samples. As these fossils represent all the archaic evolutionary lineages in Europe potentially synchronous with AKH 1, the first hypothesis does not seem to be well substantiated at the moment.
The second scenario is a migratory one. The origin and timing of this migration cannot be established at present. As was thoroughly demonstrated and discussed above, the Chinese H. erectus upper second molars are similar to AKH 1. But the Chinese fossils are dated to much earlier times (430–160 ka BP) making a direct population relationships between them and AKH 1 unlikely. Of all the Asian specimens analyzed, only Xujiayao and Denisova 4 are close to AKH 1 chronologically. But those are not among the fossils most similar to it according to the integral PCA results. Nevertheless, some “shift” of AKH 1 towards Denisovans in the volume of lateral dentin and the similarity between it and Denisova 4 in some non-metric traits (see Fig. 4) is worth noting.
The third scenario implies a wider Eurasian areal of the Middle Paleolithic populations related with Chinese H. erectus than previously thought. There are some reasons to assume that hominins similar to AKH 1 were present not only in the Caucasus but in the Near East as well: non-metric traits align this molar with MC 8 specimen from Manot Cave in Israel (see Fig. 4). The comparison of EDJ features has shown that the odontoglyphic pattern of MC 8 is not as complicated as in AKH 1, but a number of accessory fissures in the mesial area display similar archaic morphology (Sarig et al., 2021: Fig. 7). The integral PCA demonstrates that the molar from Manot aligns with EMPH owing to its small dimensions. Manot Cave is dated to the early phase of the Upper Paleolithic; thus, the “AKH 1-related” component might have been “blurred” by a later H. sapiens migration wave. Such admixture events are not excluded for the Manot Cave population (Ibid.: 14).
Summing up, the upper second molars morphologically similar to AKH 1 but belonging to different periods, except the Caucasus, were found in two geographic zones: China and Near East. Thus, the areal of the AKH 1 evolutionary lineage can only be delimited very broadly. The earliest fossils of this possible lineage came from China: Hexian, Yiyuan, Chaoxian, Hualongdong. The latest (AKH 1 and MC 8) were excavated in the Caucasus and Near East.
The question of the relationships between the “AKH 1 lineage” and the Altaian Denisovans remains open. Some connection with Neanderthals also cannot be ruled out completely for two reasons. First, some of the Neanderthal teeth, according to the PCA results, are shifted towards EAMLPH and H. erectus (see Fig. 4, 5). Second, only the upper second molar morphology has been studied, and other teeth of the AKH 1 individual might have displayed more Neanderthal-like features.
Conclusions
Our analysis has shown that the AKH 1 molar does not belong to a H. sapiens individual . Rather, it likely refers to an archaic evolutionary lineage not represented in European dental fossil collections so far. The AKH 1 molar morphology is more similar to the late Chinese H. erectus than to H. antecessor, EMPH, or Dmanisi hominins. Thus, an Asiatic origin of the “AKH 1 lineage” can be suggested. We acknowledge that the existence of this lineage, as well as determining its taxonomic status or the relationship with the Neanderthal lineage, require new fossil samples and comparisons based on wider arrays of dental traits.
Acknowledgements
