Аналитические и картографические модели функциональной устойчивости лесных экосистем

Автор: Коломыц Э.Г.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Естественные науки
Статья в выпуске: 6 т.11, 2025 года.
Бесплатный доступ
Изложены идеология и методика количественных оценок устойчивости лесных экосистем. Выделены два уровня устойчивости: лабильный фитоценотический и инерционный почвенно-биотический. Для каждого из них разработаны количественные методы расчета и картографирования потенциальной устойчивости лесных биогеоценозов двух основных типов — упруго-пластичной и резистентной. С помощью метрики евклидового расстояния выполнены расчеты индексов устойчивости. Получены статистически значимые связи в уравнениях, описывающих зависимость этих индексов от метаболических параметров лесных сообществ и факторов внешней среды. На основе полученных мер с помощью новых методов геоморфометрии рассчитаны карты-матрицы индексов устойчивости лесных биогеоценозов отдельных экорегионов, а также лесных формаций Окско-Волжского бассейна. Приведены соответственно крупно- и мелкомасштабные карты-матрицы устойчивости лесных фаций и формаций, характеризующие их чувствительность к изменениям климата и последующий функционально-восстановительный потенциал.
Лесная экосистема, функционирование, устойчивость, моделирование, картографирование
Короткий адрес: https://sciup.org/14132782
IDR: 14132782 | УДК: 9.929+57.557+50.502/504 | DOI: 10.33619/2414-2948/115/08
Analytical and Cartographic Models of Functional Stability of Forest Ecosystems
The article presents the ideology and methodology of quantitative assessments of forest ecosystem sustainability. Two levels of sustainability are distinguished: labile phytocoenotic and inertial soil-biotic. For each of them, quantitative methods for calculating and mapping the potential sustainability of forest biogeocoenoses of two main types — elastic-plastic and resistant — have been developed. The Euclidean distance metric has been used to calculate sustainability indices. Statistically significant relationships have been obtained in equations describing the dependence of these indices on the metabolic parameters of forest communities and environmental factors. Based on the measures obtained, matrix maps of sustainability indices of forest biogeocoenoses of individual ecoregions, as well as forest formations of the Oka-Volga River basin, have been calculated using new geomorphometry methods. Large-scale and small-scale matrix maps of forest facies and formations stability are provided, characterizing their sensitivity to climate change and subsequent functional-restorative potential.
Текст научной статьи Аналитические и картографические модели функциональной устойчивости лесных экосистем
Бюллетень науки и практики / Bulletin of Science and Practice
УДК 9.929+57.557+50.502/504
Любой экологический анализ территории не может обойтись без оценок устойчивости рассматриваемых объектов к внешним воздействиям, с трактовкой устойчивости как «… экологического синтеза, … кульминационной точки всех экологических зависимостей …» [1, с. 394].
Еще Г.Ф. Морозов в начале ХХ в. ставил перед лесоводами задачу такого рационального лесопользования, чтобы лес мог сохранить свою «биологическую устойчивость и производительность», с возможностью быстрого естественного возобновления коренных типов леса, которые обладают, как он полагал, наиболее высокой устойчивостью [2].
До настоящего времени в лесной экологии устойчивость экосистем описывается чаще всего качественно, с привлечением количественных признаков-индикаторов, характеризующих устойчивость в неявном виде. Оценки ведутся по скорости роста, продуктивности и плотности древостоя, по видовому и функциональному разнообразию, повреждению растительности и др. [3–7].
Общая экология располагает также достаточно глубоко разработанными математическими моделями устойчивости экосистем. В целом эти модели описывают узкий круг относительно простых экологических конструкций и имеют принципиально детерминистский характер. Объектом анализа служат обычно популяция и биотическое сообщество как совокупность популяций. Мера устойчивости подобных экосистем рассчитывается как отношение отклонения возмущающего воздействия к величине отклика по заданным свойствам на это воздействие [8, 9].
Устойчивость экосистем оценивается также по балансовым уравнениям их равновесных состояний, которые описываются потоками энергии и вещества, проходящими по трофическим цепям [10–13].
К сожалению, методы математического моделирования устойчивости таких идеализированных экосистем до сих пор не нашли широкого применения в ландшафтной экологии. Ландшафтно-экологические образования (в отличие, скажем, от объектов популяционной экологии) — это многокомпонентные системы с так называемой «плохой структурой», с огромным разрывом времен релаксации входящих в них геокомпонентов, с зарегулированной системой прямых и обратных экологических связей, с нелинейностью поведения и с господством принципиально вероятностных связей [14-17].
Эти объекты относятся к обширной группе природных систем с относительной независимостью нелинейного поведения слагающих их компонентов, с неравновесной волновой динамикой, самоорганизацией и иерархией [18, 19].
Они представляют собой относительно слабо связные природные образования, что является одним из главных условий проявления в них свойств полиморфизма как одного из важнейших факторов их устойчивой организации [20].
Функционирование и устойчивость таких систем определяются не численностью или динамикой биологических видов, а в первую очередь геохимическими циклами, т.е. годовыми и многолетними круговоротами органического вещества [13].
Именно в этом направлении и был предпринят поиск единых показателей (индексов) потенциальной устойчивости элементарных биохорологических единиц, по Н.В. Тимофеев-Ресовскому, и А.Н. Тюрюканову, — биогеоценозов как целостных образований [21], в духе учения В.Н. Сукачева [13].
Различают, как известно, устойчивость структурную и функциональную, а также резистентную (сопротивляемость к воздействию) и упругую, а точнее упруго-пластичную (восстановительный потенциал) [7, 22–24].
Предлагается рабочая концепция многоуровенного анализа устойчивости гео (эко-) систем На первом уровне рассматривается структурная устойчивость двух категорий — ареальная (геотопическая) и миграционная (трансгрессивная). Первая категория соответствует состоянию устойчивого равновесия системы в данном местоположении, вторая — ее вынужденному перемещению в пространстве как реакции на внешнее воздействие (в данном случае климатическое). Второй уровень анализа относится к функциональной устойчивости так же двух категорий — фитоценотической и почвеннобиотической . Их особенности связаны с различными характерными временами двух циклов биологического круговорота — лабильного и инерционного. Наконец, на третьем уровне рассчитываются меры фитоценотической и почвенно-биотической устойчивости двух типов — резистентной и упруго-пластичной . Первая мера характеризует чувствительность гео(эко-)системы к внешнему воздействию, вторая — ее сукцессионно-восстановительный потенциал. Количественная оценка функциональной устойчивости лесных экосистем развивалась нами на протяжении ряда лет и прошла неоднократную апробацию [20, 25, 26].
Предметом настоящего сообщения является функциональная устойчивость лесных экосистем как наиболее фундаментальная характеристика, определяющая их глобальные биосферные функции [27].
Такой подход позволяет рассчитать устойчивость непосредственно по дискретным параметрам биологического круговорота.
Меры функциональной устойчивости лесных экосистем
Биологический круговорот — сложный полициклический процесс, слагающийся из разнопорядковых по характерному времени циклов производства живого органического вещества, его разложения, минерализации и гумификации [28, 29].
Соответственно и устойчивость экосистем носит разнопорядковый характер. Для бореальных и суббореальных лесов нами выделены два уровня устойчивости: лабильный фитоценотический и инерционный почвенно-биотический .
Лабильная устойчивость выражена двумя комплексными дискретными параметрами метаболизма — коэффициентом годового оборота надземной фитомассы KR и подстилочно-опадным индексом, т.е. коэффициентом годичной деструкции [25].
Индекс лабильной упруго-пластичной I упр (1) устойчивости лесного биогеоценоза рассчитывался как мера евклидового расстояния от его оптимального функционального состояния:
Iупр(1) = 1 - [ 7(AKR)2 + (AKY)2 ] / V2, где (∆KR) = (KRmax─ KRi) / (KRmax─ KRmin); (∆KY) = (KYi ─ KYmin) / (KYmax─ KYmin).
Здесь оба предиктора рассматриваются как равноправные, т.е. берутся с единичными «весами». Аналогично вычислялся индекс потенциальной резистентной устойчивости I рез (1) экосистемы. В этом случае за оптимумы принимались минимальное значение KR и максимальное — KY . По данным 377 пробных площадей, охватывающих Окско-Волжский бассейн от юга подтаежной зоны до подзоны южной лесостепи, установлено, что I упр(1) имеет весьма тесную параболическую связь с параметром KR , между тем как Ι рез (1) наиболее отчетливо коррелирует с массой подстилки. Если первоначальная реакция лесного биогеоценоза на внешние сигналы зависит главным образом от резервного фонда в виде подстилочной массы, то дальнейший адаптивно-восстановительный потенциал определяется почти исключительно годовым оборотом надземной фитомассы. Рассчитаны времена проявления I рез (1) и I упр (1) лесных биогеоценозов при климатических изменениях.
Инерционный почвенно-биотический уровень устойчивости I рез (2) и I упр (2) охватывает более обширный и более медленный метаболический цикл [26].
Он включает компонент устойчивости, связанный с формированием и динамикой органического вещества почвы, поэтому в расчетные формулы помимо параметра KR входят масса лесной подстилки ML и масса гумуса HU :
I 616(2) = 1
(1 - KR )2
+ a • ML 2 + b • (1 - HU )2
1 + a + b
1 6a?(2) = 1 -
KR 2 + a • (1 - ML )2 + b • HU 2
1 + a + b
Вначале принято, что «весовые» коэффициенты a = b = 1. Это по-прежнему означает равноправное участие всех рассматриваемых факторов в формировании устойчивости лесных биогеоценозов. Результаты расчетов Ι i упр(2) и Ι i рез(2) уже существенно отличаются от данных, рассчитанных по формуле (1) и ее аналогу для Ι рез(1).
Дальнейшее приближение в расчетах индексов инерционной устойчивости экосистем связано с введением «весовых» коэффициентов при предикторах ML и HU . Весовые коэффициенты характеризуют вклад каждого метаболического признака в ту или иную устойчивость. Эти коэффициенты отыскивались путем минимизация некоего «потенциала», который становился бы константой для идеально однородной в пространстве экосистемы. С нею в данном случае ассоциируется лесной биогеоценоз. В качестве такого «потенциала» в работе «Ряды стабильностей пар элементов в серой лесной почве» был предложен коэффициент вариации Kvar , введенный в 1895 г. K. Пирсоном и представляющий собой отношение стандартного отклонения данной величины к ее среднему значению, умноженное на 100% [30].
Проведено сравнение двух индексов упругой устойчивости: I упр(1), рассчитанных по формуле (1), и I упр(2) — по формуле (3). Параметры имеют весьма высокую корреляцию — R2 = 0,913 (Рисунок 1).
Таким образом, лабильная фитоценотическая устойчивость, может быть принята (в первом приближении) в качестве показателя общей устойчивости лесной экосистемы, в том числе инерционной. Полученный вывод представляется важным в методическом отношении. Меры лабильной устойчивости основываются на более простых и более строгих соотношениях параметров биологического круговорота по сравнению с мерами устойчивости инерционной.
Функциональная устойчивость лесов в системе их экологических связей
На примере Среднего Поволжья показано, что территориальные контрасты в индексах обоих типов инерционной устойчивости лесов носят достаточно четкий зональный характер (Таблица 1).
На границе лесостепной и степной зон в резистентной устойчивости ведущую роль играет масса лесной подстилки, а в упругой — масса гумуса, при одинаково слабом влиянии параметра KR . В направлении от южной лесостепи к северной границе лесостепной зоны масса подстилки сохраняет доминирующий положительный вклад в распределение I рез(2) лесных биогеоценозов. Одновременно резко возрастает ее отрицательная роль в восстановительном потенциале лесов, на фоне все более снижающейся роли скорости годичного оборота фитомассы. Резкий перелом в факторной ординации индексов обоих
Бюллетень науки и практики / Bulletin of Science and Practice Т. 11. №6 2025 типов устойчивости по метаболическим параметрам происходит на границе лесостепной и подтаежной зон, которая образует известный Главный ландшафтный рубеж Русской равнины. Здесь происходит скачкообразное падение роли подстилочной массы. При этом для Iупр(2) в разряд доминирующего фактора выходит масса гумуса.
Таблица 1
ДОЛИ ПОЛОЖИТЕЛЬНОГО ИЛИ ОТРИЦАТЕЛЬНОГО УЧАСТИЯ РАЗЛИЧНЫХ НОРМИРОВАННЫХ ПРЕДИКТОРОВ ( KR , ML и HU, )
в индексах инерционной устойчивости лесных экосистем Среднего и Верхнего Поволжья
|
Региональная экосистема (экспериментальный полигон)*) |
Инерционная устойчивость |
|||||
|
резистентная Iрез(2) |
упругая Iупр(2) |
|||||
|
KR |
ML |
HU |
KR |
ML |
HU |
|
|
Южная граница южной лесостепи, сосняки, дубравы, липняки (низкогорный массив Жигули) |
–0.289 |
+0.565 |
–0.146 |
+0.346 |
–0.461 |
+0.193 |
|
Приволжская возвышенность, типич-ная лесостепь, сосновые и липово-дубовые леса (Присурье) |
–0.107 |
+0.798 |
–0.095 |
+0.149 |
–0.617 |
+0.234 |
|
Нижегородское Предволжье, северная лесостепь, дубово-липовые и сосновошироколиственные леса (Кудьма) |
–0.132 |
+0.507 |
–0.136 |
+0.131 |
–0.449 |
+0.420 |
|
Нижегородское Предволжье, северная лесостепь, дубравы, липняки (возвышенный массив Щелоковский Хутор) |
–0.168 |
+0.711 |
–0.121 |
+0.124 |
–0.698 |
+0.178 |
|
Нижегородское Предволжье, северная лесостепь, смешанные и хвойные леса (урочище Зеленый Город) |
–0.158 |
+0.194 |
–0.648 |
+0.222 |
–0.050 |
+0.728 |
|
Нижнее низменное Приочье, южная граница подтайги, елово-сосновые зандровые полесья (Выкса) |
–0.139 |
+0.187 |
–0.674 |
+0.245 |
–0.118 |
+0.637 |
|
Среднее низменное Приочье, южная граница подтайги, смешанные, хвойные и широколиственные леса (Приокско-Террасный заповедник) |
–0.334 |
+0.365 |
–0.301 |
+0.391 |
–0.174 |
+0.435 |
|
Нижегородское низменное Заволжье, южная полоса под-таежной зоны, сосновые и еловые леса (Керженец) |
–0.177 |
+0.484 |
–0.339 |
+0.252 |
–0.263 |
+0.485 |
|
Основной водосбор Волжского бассейна в целом (статистическая выборка по всем полигонам) |
–16.2 |
+55.8 |
–18.6 |
+61.6 |
–36.1 |
+2.3 |
*) Более подробное описание экспериментальных полигонов и их расположение приведены в работе Dı́az S., Cabido M. [6].
На Рисунке 1 дано сравнение двух индексов упругой устойчивости: I упр(1), рассчитанных по формуле (2), и I упр(2) — по формуле (6). Параметры в целом имеют весьма высокую корреляцию, однако они адекватны друг другу только в своих центральных значениях (около 0,50). При устойчивости менее 0,40 параметр I упр(1) оказывается на 0,05– 0,10 единиц больше, чем I упр(2), а при общих высоких показателях устойчивости (> 0,75– 0,80) соотношение обратное. Запасы гумуса способствуют усилению восстановительного потенциала только для лесных экосистем с высокими значениями KR . Когда же эти значения малы, уход части мертвой органики в гумус снижает упругую устойчивость экосистемы.
Таким образом, устойчивость ежегодного возобновления живой надземной фитомассы, т.е. лабильная фитоценотическая устойчивость, может быть принята (в первом приближении) в качестве показателя общей устойчивости лесной экосистемы , в том числе инерционной, связанной с гораздо более длительным почвенно-биотическим циклом метаболизма. Полученный вывод представляется важным в методическом отношении. Меры лабильной устойчивости основываются на более простых и более строгих соотношениях параметров биологического круговорота по сравнению с мерами устойчивости инерционной.
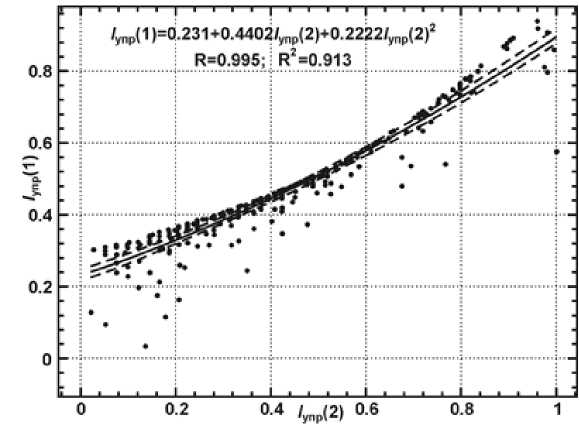
Рисунок 1. Соотношение значений упругой устойчивости лесных экосистем, полученной по разным формулам
Дальнейшее приближение в расчетах индексов инерционной устойчивости экосистем связано с введением «весовых» коэффициентов a и b для предикторов. Эти коэффициенты отыскивались эмпирически — путем минимизация некоего «потенциала», который становился бы константой для идеально однородной в пространстве экосистемы, с которой в данном случае ассоциируется лесной биогеоценоз. В итоге были получены индексы устойчивости I рез(3) и I упр(3). Построены уравнения линейной регрессии I рез(3) и I упр(3) лесных экосистем для двух экорегионов Окско-Волжского бассейна: Самарской Луки (СЛ, полигон Жигули) и Приокско-Террасного заповедника (полигон ПТЗ). В уравнениях предикторы перечислены в порядке убывания их значимости, которая определена модулем t -статистики (нижний индекс при каждом предикторе).
I рез(3) СЛ = – 0,6164 ⋅ KR –24.59 – 0,2920 ⋅ HU –13.40 + 0,08769 ⋅ ML +4.36 + 0,8690;
R2 = 0,962; Degr = 1,6%; P < 10–6 .(4)
I упр(3) СЛ = – 0,4463 ⋅ ML –21,26 + 0,,3058 ⋅ KR +11,70 + 0,1242 ⋅ HU +5,46 + 0,,4607;
R2 = 0,954; Degr = 1,9%; P < 10–6 ,(5)
I рез(3) ПТЗ = – 0,4805 ⋅ HU –28,40 – 0,3093 ⋅ KR –18,44 + 0,2150 ⋅ ML +11,26 + 0,7716;
R2 = 0,977; Degr = 0,6%; P < 10–6 ,(6)
Iупр(3)ПТЗ = – 0,7046⋅ML –35,94 + 0,1427⋅HU +8,22 + 0,05522⋅KR +3,21 + 0,7473;(7)
R2 = 0,972; Degr = 0,8%; P < 10–6 .
Здесь R2 и P — соответственно коэффициент детерминации и критерий значимости Пирсона. Верификация моделей проведена по критерию ее деградации Degr, который рассчитывался по методике кросс-валидации Аллена [31]. Отсюда рассчитаны следующие доли участия всех предикторов в индексах устойчивости (для сравнения см. Таблицу 1 по этим же полигонам):
Самарская Лука Приокско-Террасный заповедник
|
KR |
ML |
HU |
KR |
ML |
HU |
|
|
а) для I рез(3) |
–61,9 |
+8,8 |
–29,3 |
–30,8 |
+21,4 |
–47,8 |
|
б) для I упр(3) |
+34,9 |
–50,9 |
+14,2 |
+6,1 |
–78,1 |
+15,8 |
В результате получено более четкое представление о механизмах функционирования лесного сообщества, которые обеспечивают ему как резистентную, так и упругую устойчивость. Ее механизмы не могут срабатывать без участия гумусной массы, роль которой в стабилизации лесного биогеоценоза возрастает при снижении ее содержания, что отвечает известному «закону» минимума лимитирующих факторов Либиха [32].
Выявлено также, что высокая резистентная устойчивость лесного сообщества поддерживается главным образом путем ослабления автотрофного биогенеза (– KR ) и во вторую очередь — замедлением процессов деструкции (+ ML ). Реализация же восстановительного потенциала определяется главным образом увеличением активности детритной ветви метаболизма, о чем говорит высокая отрицательная связь индекса упругой устойчивости с массой подстилки. Этому способствует также рост автотрофного биогенеза (+ KR). Роль его в экогенетических сукцессиях может быть как соразмерной с активным детритогенезом, снижающим значение ML , так и второстепенной. Таким образом, в процессе восприятия неблагоприятных внешних сигналов лесной биогеоценоз переключается с одних ведущих процессов своего функционирования на другие — со скорости автотрофного биогенеза на темпы разложения лесной подстилки. При этом раскрываются два взаимно противоположных механизма проявления известных буферных свойств подстилки [2, 33].
В масштабе всего основного водосбора Волжского бассейна наиболее значимыми метаболическими параметрами-предикторами (с положительной дифференцирующей силой влияния — не менее 55%–60%) являются: для резистентной устойчивости — масса лесной подстилки, и для устойчивости упругой — коэффициент годичного оборота надземной фитомассы (Таблица 1, нижняя графа). Исходя из этого, общий анализ функциональной устойчивости лесных формаций этой территории был проведен по их лабильной фитоценотической устойчивости (Таблица 2), что существенно упростило процедуру анализа.
Территориальная дифференциация устойчивости лесных экосистем
В картографировании устойчивости лесных экосистем были использованы новые методы геоморфометрии — науки о моделировании рельефа, особенно развившейся за последние 30 лет. Применена разработанная П.А. Шарым система из 18 параметров, с более качественными алгоритмами расчета морфометрических характеристик [34].
Были использованы спутниковые данные НАСА о рельефе — матрицы высот земной поверхности разрешения 90 м, полученные в ходе выполнения международного проекта SRTM «США-Италия-Германия» (Shuttle Radar Topography Mission — проект радарной съемки рельефа спутником Шаттл). Проводились процедуры интерполяция и экстраполяция измеренных и рассчитанных данных в точках по матрицам факторов среды, для которых множественная регрессия выявила наиболее тесные статистические связи. Подучены соответствующие уравнения регрессии, по которым и выполнялось крупномасштабное картографирование (Рисунок 2 и 3).
0,24 0,28 0,29 0,32 0,45 0,52 0,87
0,20
0,56 0,75 0,84 0,87 0,89 0,97
Рисунок 2. Карты индексов упругой (а) и резистентной (б) инерционной устойчивостей для
Самарской Луки, рассчитанные по моделям (4) и (5)


Рисунок 3. Карты индексов упругой (а) и резистентной (б) устойчивостей для Приокско-Террасного заповедника, рассчитанные по моделям (6) и (7). Индексы устойчивости для карты (а): 1 – 0,01–0,44; 2 – 0,44–0,51; 3 – 0,51–0,58; 4 – 0,58–0,63 5 – 0,63–0,69; 6 – 0,69–0,77; 7 – 0,77–0,92. Индексы устойчивости для карты (б): 1 – 0,39–0,47; 2 – 0,47–0,52; 3 – 0,52–0,58; 4 – 0,58–0,63; 5 – 0,63–0,67; 6 – 0,67–0,71; 7 – 0,71–0,94
Установлено, что на зональном экотоне леса и степи (полигон СЛ) максимальным восстановительным потенциалом обладают мезоморфные плакорные дубо-липняки Жигулевского плато и мезогидроморфные супераквальные леса глубоко врезанных долин. Гораздо менее устойчивы ксероморфные трансэлювиальные и транзитные остепненные сосняки и сосново-широколиственные леса. На южной границе подтаежной зоны (полигон ПТЗ) наиболее устойчивы, с одной стороны, ксеро-мезоморфные липово-березово-осиновые леса, произрастающие на карбонатном элювии, а с другой, — заболоченные сосняки, ельники и черноольшаники днищ речных долин и междуречных западин. Плакорные мезоморфные сосново-липово-дубовые леса, а также трансаккумулятивные мезо-гидроморфные ельники с сосной отличаются минимальной упругой устойчивостью. Переход с локального уровня картографирования устойчивости лесов Окско-Волжского бассейна на региональный осуществлялся с помощью специально разработанного метода индукционноиерархической экстраполяции [20], основанного на эмпирически установленном явлении полизональности локальных экосистем как формы их реакции на глобальные изменения климата. Построены карты индексов лабильной устойчивости лесных формаций Окско-Волжского бассейна (Рисунок 4 и 5).
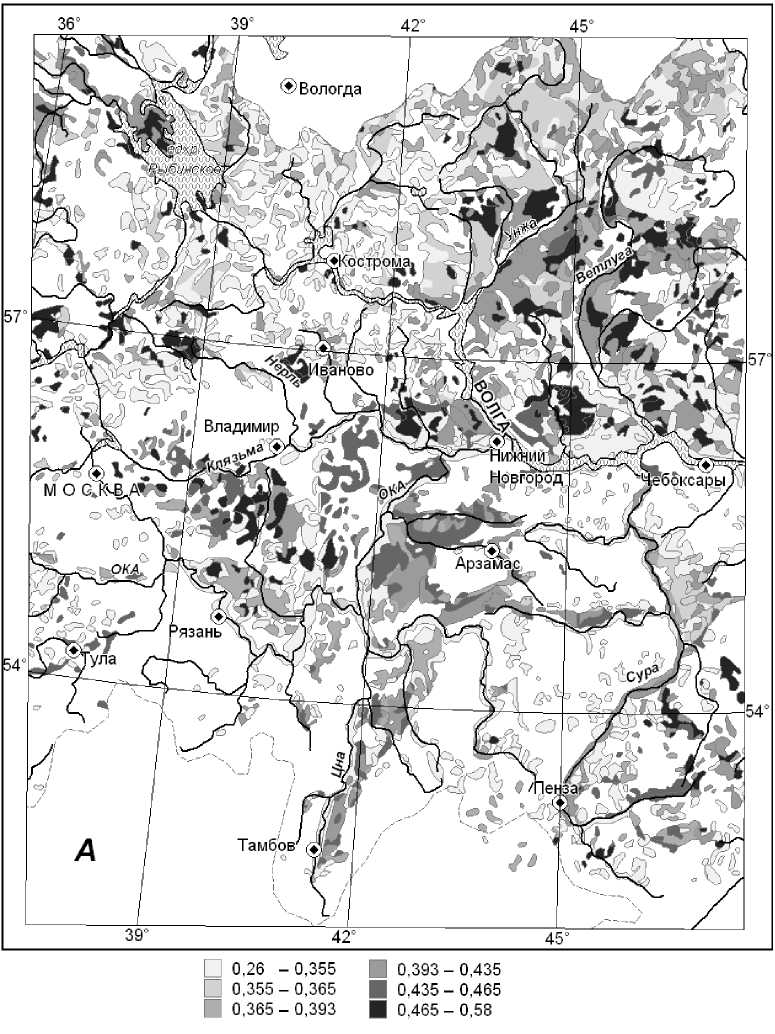
Рисунок 4. Карта индексов резистентной лабильной устойчивости групп лесных формаций территории Окско-Волжского бассейна
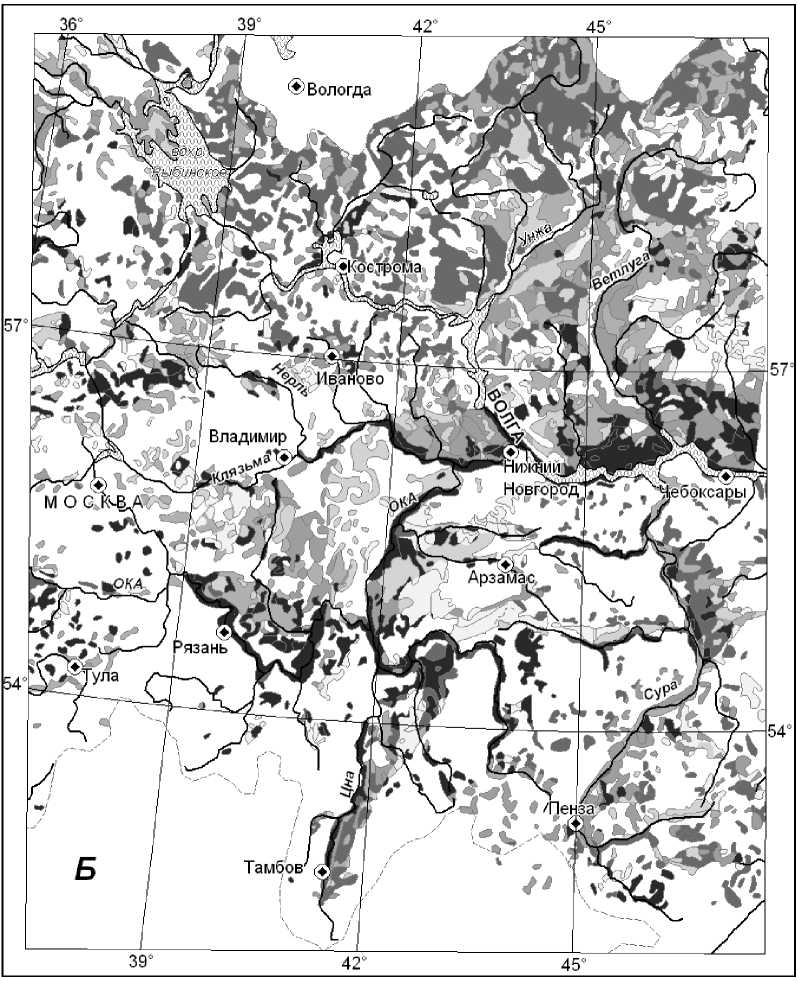
0.26 - 0,355 И 0,393- 0,435 0,355- 0,365 И 0.435- 0.465 0,365- 0,393 И 0,465- 0,58
Рисунок 5. Карта индексов упруго-пластичной лабильной устойчивости групп лесных формаций территории Окско-Волжского бассейна
Общий анализ функциональной устойчивости лесных формаций этой территории был проведен по их лабильной фитоценотической устойчивости (Таблица 2), что существенно упростило процедуру анализа.
Выявлено, что средне- и южнотаежные еловые и широколиственно-еловые подтаежные леса имеют пониженную устойчивость обоих типов. Средне- и южнотаежные сосновые леса отличаются, наоборот, более высокой чувствительностью к воздействиям, но менее развитыми механизмами функционального восстановления. Широколиственно-сосновые и широколиственные леса весьма чувствительны к внешним сигналам, но способны к сравнительно быстрому функциональному восстановлению.
|
ТАКСОНОМИЧЕСКИЕ (СРЕДНЕВЗВЕШЕННЫЕ) НОРМЫ РЕЗИСТЕНТНОЙ И УПРУГОЙ ЛАБИЛЬНОЙ УСТОЙЧИВОСТИ РАСТИТЕЛЬНЫХ ФОРМАЦИЙ ОКСКО-ВОЛЖСКОГО БАССЕЙНА |
Таблица 2 |
|||
|
Класс (подкласс) растительных |
Тип (подтип) растительности |
Группа растительных формаций |
Индекс устойчивости |
|
|
формаций |
резисте нтной |
упругой |
||
|
Североевропейские еловые леса |
А. Еловые южнотаежные и широколиственно- |
(48 б). Ельники южнотаежные зеле-номошно-травяно-кустарничковые |
0,568 |
0,390 |
|
еловые (подтаежные) леса |
(52). Ельники с сосной, сложные. неморальнотравяные |
0,606 |
0,284 |
|
|
(53). Широколиственно-еловые неморальнотравяные леса |
0,338 |
0,460 |
||
|
(54). Березово-черноольховые с елью болотно-крупнотравные леса |
0,270 |
0,627 |
||
|
Сосновые и широколиственно- |
Б. Сосновые средне-и южнотаежные леса |
(76). Сосняки. с елью и березой. кустарничково-зеленомошные |
0,446 |
0,398 |
|
сосновые леса и производные |
(78). Сосняки. с березой, кустарничковые долгомошно-сфагновые |
0,388 |
0,587 |
|
|
сообщества на их месте. Североевропейские |
(763). Сосняки с елью, зеленомошно-сфагновые, евтрофные болота |
0,384 |
0,601 |
|
|
сосновые леса |
В. Широколиственно- |
(81). Сосняки кустарничково-травяные. с дубом и липой в подлеске |
0,514 |
0,434 |
|
сосновые (подтаежные) леса |
(84). Широколиственно-сосновые леса чернично-разнотравнозлаковые |
0,419 |
0,457 |
|
|
(812). Сосняки кустарничковотравяные. с евтрофными болотами |
0,300 |
0,547 |
||
|
Широколиственны е леса и производные сообщества на их месте. Европейские широколиственные |
Г. Восточноевропейски е широколиственные леса |
(118 а). Дубовые и липоводубовые леса северные, с примесью ели (118 б). Дубовые и липоводубовые леса южные, остепненные |
0,420 0,245 |
0,500 0,609 |
|
леса |
(118 в). Приволжские. дубовые и липово-дубовые осветленные леса |
0,311 |
0,541 |
|
|
(118 г). Приволжско-заволжские липово-дубовые осветленные леса |
0,398 |
0,567 |
||
|
(120 а). Липняки, с дубом и вязом. разнотравно-злаковые, мезофитные |
0,320 |
0,517 |
||
Примечание: номера групп формаций указаны по Исаченко, Лавренко (1974) [35]
Итак, большинство лесных формаций Окско-Волжского бассейна характеризуются высокой чувствительностью к внешним возмущениям, но в то же время — достаточно развитыми механизмами, олицетворяющими их упругую устойчивость. Это означает, что экзогенная динамика функциональных и структурных параметров лесных экосистем региона отражает смену их первоначальной адаптивной стратегии в меняющейся окружающей среде на стратегию последующего устойчивого развития, стремящегося привести их в первоначальное либо новое устойчивое функциональное состояние после «снятия» или даже
Бюллетень науки и практики / Bulletin of Science and Practice Т. 11. №6 2025 ослабления воздействий (например, стабилизации климата, уменьшения загрязнения природных сред, снижения рекреационной или пастбищной нагрузки).
Заключение
-
1. Впервые в ландшафтной экологии и лесоведении выделены две категории функциональной устойчивости лесных экосистем: лабильную фитоценотическую и инерционную почвенно-биотическую. По каждой категории разработаны количественные методы расчета, а также крупно- и мелкомасштабного картографирования потенциальной устойчивости лесных экосистем двух основных типов – резистентной и упруго-пластичной. Методы позволяют оценивать устойчивость лесных биогеоценозов и формаций как целостных хорологических единиц (в духе учения В.Н. Сукачева и концепции Н.В. Тимофеева-Ресовского), а не по отдельным их структурным или функциональным характеристикам, как это практикуется до сих как в нашей стране, так и за рубежом. Новизна и эффективность разработанных методов состоит в том, что они опираются на дискретные параметры биологического круговорота.
-
2. Проведена количественная оценка относительного вклада каждого метаболического фактора в тот или иной индекс устойчивости лесов на бореальном экотоне Волжского бассейна и проведена верификация расчетных моделей. На основе полученных мер с помощью новых методов геоморфометрии рассчитаны карты-матрицы индексов устойчивости биогеоценозов. Картирование осуществлено путем интерполяция и экстраполяция индексов по матрицам наиболее значимых факторов среды. Для регионального картографирования устойчивости разработан метод индукционноиерархической экстраполяциии
-
3. Моделирование устойчивости лесных биогеоценозов и формаций в лесном поясе Среднего и Верхнего Поволжья показало следующее. В начальный период воздействия неблагоприятного внешнего сигнала лесная экосистема, стремясь к выживанию, реагирует снижением интенсивности годового оборота надземной фитомассы и/или усилением минерального питания растений из гумусового горизонта почвы. Последующий функционально восстановительный потенциал реализуется с помощью активизации процессов деструкции мертвой органики в лесной подстилке и дебрисе, а затем — путем увеличения годичной продукции зеленой массы, т.е. общим ускорением биологического круговорота.
