Анализ полиморфизма гена BoLA-DRB3 у симментальской породы крупного рогатого скота

Автор: Козлов Александр Леонидович, Смазнова Ирина Александровна, Заякин Владимир Васильевич, Нам Ирина Янгуковна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Генетика
Статья в выпуске: 5-3 т.13, 2011 года.
Бесплатный доступ
Проанализирован аллельный полиморфизм гена BoLA-DRB3 в стаде крупного рогатого скота симментальской породы. Проанализирована связь некоторых аллелей с молочной продуктивностью. Не было выявлено корреляций между удойностью и аллелями устойчивости или чувствительности.
Симментальская порода, аллели устойчивости, аллели чувствительности
Короткий адрес: https://sciup.org/148200458
IDR: 148200458 | УДК: 575.174.015.3+575.113.2
Analysis of gene BoLA-DRB3 polymorphism in simmental cattle
Gene BoLA-DRB3 polymorphism has been analyzed in Simmental cattle. Correlation between alleles and milk yield has been carried out. There wasn't found any correlation between milk production and resistance or susceptibility alleles.
Текст научной статьи Анализ полиморфизма гена BoLA-DRB3 у симментальской породы крупного рогатого скота
На сегодняшний день проблема лейкоза крупного рогатого скота (КРС) является одной из наиболее актуальных в животноводстве. Это обусловлено в первую очередь значительными экономическими потерями, причиняемыми данным заболеванием. При этом за последнее десятилетие наблюдается значительный рост числа заболевших животных [1, 2]. Проблема усугубляется также и тем, что лейкоз КРС во многом является малоизученным заболеванием, безопасность которого для человека четко не установлена. Вирус лейкоза крупного рогатого скота (ВЛ КРС) относится к группе ретровирусов типа С и по своей организации очень похож на вирус, вызывающий лейкемию у человека. Известны случаи проявления клинических признаков лейкоза у овец, свиней и шимпанзе при заражении их ВЛ КРС [3]. Кроме того, традиционная селекционная работа направлена в основном на улучшение признаков, связанных с продуктивностью, тогда как признаки, ответственные за устойчивость к болезням, не учитывались. Это стало причиной увеличения подверженности животных за болеваниям, в частности лейкозу [4].
Главный комплекс гистосовместимости (ГКГ) является ключевым в организации иммунного ответа организма. Входящий в него ген BoLA-DRB3 определяет устойчивость животных к лейкозу. Он высокополиморфен и к моменту исследования известно более 90 его аллелей, из которых для трех установлена их связь с устойчвостью к лейкозу, а для четырех – с восприимчивостью [5,6].
В работах, проведенных ранее, показано различие в полиморфизме гена BoLA-DRB3 у различных пород КРС [7,8]. При этом устойчивость породы к лейкозу в целом определяется долей аллелей устойчивости в её генофонде .
Цель данного исследования – анализ полиморфизма второго экзона гена BoLA-DRB3 у
животных симментальской породы в сравнении с айрширской и черно-пестрой, а также выявление связи его аллелей с молочной продуктивностью.
МАТЕРИАЛ И МЕТОДЫ
Исследование было проведено на КРС симментальской породы из колхоза «Большевик» Стародубского р-на Брянской обл. Всего проанализировано 77 особей.
ДНК животных выделяли из цельной крови коров согласно методике [9]. Для выявления различных аллелей гена BoLA-DRB3 использовали метод ПЦР-ПДРФ. Температурный профиль реакции для анализа полиморфизма гена BoLA-DRB3:денатурация ДНК – 940С, 3 мин. – первый цикл, денатурация – 940С, далее по 1 мин. Отжиг праймеров – 650С, 1 мин., элонгация ДНК – 720С, 60 сек. количество циклов – 28. Достраивание одноцепочечной формы ДНК – 720С – 5 мин согласно [10, 11]. Рестрикция фрагмента гена BoLA-DRB3 длиной 284 п.о., полученного при амплификации, осуществлялась с помощью эндонуклеаз рестрикции RsaI, BstIY и HaeIII. Продукты рестрикции анализировали электрофорезом в 9% полиакриламидном геле.
Связь с молочной продуктивностью возможно было проследить только для тех аллелей, которые имели наибольшую частоту в выборке. При этом общая совокупность генотипов разбивалась на две. Первая была представлена генотипами гомозиготными в отношении анализируемого аллеля в сумме с генотипами, несущими данный аллель вместе с редкими нейтральными в отношении лейкоза аллелями, не вошедших в выборки других анализируемых аллелей. Вторая совокупность особей, большая по количеству, была представлена всеми остальными генотипами выборки, не несущими анализируемый аллель. Каждая совокупность была проанализирована на характер распределения с помощью теста Колмогорова-Смирнова.
Статистическое различие между парами совокупностей оценивалось с помощью U-теста Манна-Уитни: при значении его p-уровня меньше 0,05 различие между совокупностями считалось статистически достоверным. Для оценки характера данного различия использовали среднее арифметическое. Все вычисления проводились с помощью пакета SPSS 15.0.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В ходе исследования выявлено неравномерное распределение аллелей в выборке (табл. 1.).
Таблица 1 . Частоты аллелей гена BoLА-DRB3 в исследованных стадах
|
№ алле ля |
Частота,% |
χ 2 |
Частота,% |
χ 2 |
Частота,% |
χ 2 |
|
симментальская |
айрширская |
черно-пестрая |
||||
|
1 |
0,65 |
5,02 |
- |
- |
0,79 |
2,91 |
|
2 |
0,65 |
5,02 |
1,06 |
4,30 |
0,79 |
2,91 |
|
3 |
0,65 |
5,02 |
- |
- |
0,79 |
2,91 |
|
5 |
2,60 |
2,13 |
- |
- |
- |
- |
|
6 |
- |
- |
- |
- |
0,79 |
2,91 |
|
7 |
16,23 |
15,95 |
39,36 |
175, 42 |
2,38 |
0,89 |
|
8 |
7,14 |
0,13 |
1,06 |
4,30 |
9,52 |
6,16 |
|
9 |
3,90 |
0,89 |
- |
- |
- |
- |
|
10 |
11,04 |
3,67 |
1,06 |
4,30 |
5,56 |
0,34 |
|
11 |
12,99 |
7,26 |
4,26 |
0,64 |
14,29 |
22,72 |
|
12 |
0,65 |
5,02 |
4,26 |
0,64 |
0,79 |
2,91 |
|
13 |
1,95 |
2,96 |
- |
- |
2,38 |
0,89 |
|
14 |
- |
- |
- |
- |
1,59 |
1,75 |
|
15 |
1,95 |
2,96 |
3,19 |
1,50 |
6,35 |
0,92 |
|
16 |
11,69 |
4,73 |
2,13 |
2,72 |
1,59 |
1,75 |
|
18 |
5,84 |
0,03 |
- |
2,72 |
- |
- |
|
19 |
- |
- |
4,26 |
0,64 |
0,79 |
2,91 |
|
20 |
1,95 |
2,96 |
2,13 |
2,72 |
5,56 |
0,34 |
|
22 |
2,60 |
2,13 |
- |
- |
6,35 |
0,92 |
|
23 |
- |
- |
8,51 |
0,82 |
15,08 |
26,49 |
|
24 |
10,39 |
2,74 |
9,57 |
1,77 |
13,49 |
19,23 |
|
25 |
0,65 |
5,02 |
6,38 |
0,00 |
0,79 |
2,91 |
|
26 |
- |
- |
- |
- |
0,79 |
2,91 |
|
28 |
1,30 |
3,92 |
8,51 |
0,82 |
3,97 |
0,03 |
|
30 |
- |
- |
- |
- |
1,59 |
1,75 |
|
32 |
1,30 |
3,92 |
- |
- |
- |
- |
|
33 |
1,95 |
2,96 |
- |
- |
- |
- |
|
36 |
0,65 |
5,02 |
2,13 |
2,72 |
3,97 |
0,03 |
|
46 |
0,65 |
5,02 |
- |
- |
- |
- |
|
53 |
0,65 |
5,02 |
- |
- |
- |
- |
Прим.: 1) аллели устойчивости, 2) аллели чувствительности
Для сравнения приведены данные полученные нами ранее для айрширской и черно-пестрой пород. Наибольшей частотой для симментальской породы характеризуются аллели *7, *10, *11, *16, *24. Из них аллель *11 относится к аллелям устойчивости, *16, *24 - чувствительности, а *7 и *10 являются нейтральными. Частота аллеля *11
составила 12,99%, *16 - 11,69%, *24 - 10,39%. Довольно высока доля нейтральных аллелей: *7 -16,23%, *10 - 11,04%. Что касается остальных нейтральных аллелей, то их частота значительно ниже.
Распределение аллелей различно среди разных пород, что возможно связано с различным их происхождением. Для симментальской породы характерно наличие ряда аллелей, не присутствующих у других пород: *5, *9, *32, *33, *46, *53. Профиль аллелей связанных с устойчивостью к лейкозу также различен среди пород. Для исследованной выборки симментальской породы характерна высокая частота аллелей чувствительности. В то же время ввиду доминантного характера наследования аллелей устойчивости [12] важно то, в каком состоянии находятся аллели. Диаграмма, отображающая распределение аллелей по генотипам, представлена на рис.
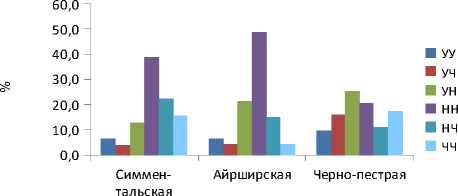
Рис. Соотношение генотипов устойчивых, нейтральных и чувствительных к вирусу лейкоза животных в изученных стадах
Ввиду сложной структуры главного комплекса гистосовместимости, интересным является проанализировать связь между полиморфизмом гена BoLA-DRB3 и удойностью. В работах, посвященных исследованию структуры ГКГ, отмечается сцепление между различными локусами этого региона [13, 14]. Некоторые исследователи отмечают связь между отдельными аллелями и параметрами молочной продуктивности [15].
Таблица 2. Связь аллелей с молочной продуктивностью
|
№ аллеля |
M±m |
σ2 |
p, Манна-Уитни |
n |
|
7 |
3954,9±90 |
80407,2 |
0,929 |
10 |
|
10 |
4010,4±167 |
139336,3 |
0,811 |
5 |
|
11 |
4011,33±122 |
134923,0 |
0,679 |
9 |
|
16 |
4008,77±133 |
230928,2 |
0,950 |
13 |
|
24 |
3777,4±31 |
4778,8 |
0,121 |
5 |
В представленной выборке симментальского скота были выбраны несколько совокупностей генотипов, несущих анализируемый аллель. Данные анализа представлены в таблице 2. В ходе проведенного анализа не было выявлено каких- либо корреляций между удойностью и наличием определенного аллеля.
Таким образом, проведенное исследование показало необходимость учета аллелей гена BoLA-DRB3.2 при организации племенной работы, т.к. в проанализированных стадах довольно высоко содержание аллелей чувствительности к лейкозу одновременно с низкой долей аллелей устойчивости.
Проведенный анализ связи между молочной продуктивностью и полиморфизмом исследуемого участка не выявил статистически значимых корреляций.
Работа выполнена при поддержке грантов ФЦП № 02.740.11.0285, АВЦП № 2.1.1/224 и гранта администрации Брянской области № 215.
Список литературы Анализ полиморфизма гена BoLA-DRB3 у симментальской породы крупного рогатого скота
- Гулюкин М.И., Симонян Г.А., Замараева Н.В. и др. Научные основы профилактики и борьбы с лейкозом крупного рогатого скота//Труды ВИЭВ. 1999. Т. 72. С. 38-47.
- Коровин Р.Н., Богданов А.И., Борисенкова А.Н. Построение обобщенного показателя оценки эпизоотической ситуации//Доклады РАСХН. 2002. Т. 2. С. 54-55.
- Schwartz I., Levy D. Pathology of bovine leukemia virus//Vet. Res. 1994. V. 25. P. 521-536.
- van Dorp R.T., Martin S.V., Shoukri M.M. et al. Epidemiologic study of disease in 32 registered Holstein dairy herds in British Columbia//Can. J. Veterin. Res. 1999. V. 63. P. 185-192.
- Harris A. L., George C R., Elizabeth L.G. Comparative organization and function of the major histocompatibility complex of domesticated cattle//Immun. Rev. 1999. V. 167. P. 145-158.
- Mirsky M. L., Olmstead C., Da Y., Lewin H.A. Reduced bovine leukaemia virus proviral load in genetically resistant cattle//Anim. Gen. 1998. V. 29. P. 245-252.
- Рузина М.Н. и др. Полиморфизм гена BoLA-DRB3 у крупного рогатого скота монгольской, калмыцкой и якутской пород//Генетика. 2010. Т. 46. № 4. С. 517-525.
- Сулимова Г.Е., Удина И.Г., Шайхаев Г.О., Захаров И.А. ДНК-полиморфизм гена BoLA-DRB3 у крупного рогатого скота в связи с устойчивостью и восприимчивостью к лейкозу//Генетика. 1995. Т. 31. № 9. С. 1294-1299.
- Boom R., Sol C.J.A., Salimans M.M.M. Rapid and simple method for purification of nucleic acids//J. Clin. Microbiol. 1990. V. 28. P. 495-503.
- van Eijk M.J.T., Stewart-Haynes J.A., Lewin H.A. Extensive polymorphism of the BoLA-DRB3 gene distinguished by PCR-RFLP//Anim. Gen. 1992. V. 23. P. 483-496.
- Xu A., Van Eijk M.J.T., Park Ch., Lewin H.A. Polymorphism in BoLA-DRB3 exon 2 correlates with resistance to persistent lymphocytosis caused by bovine leukemia virus//J. Immunol. 1993. V. 151. P. 6977-6985.
- Удина И.Г., Карамышева Е.Е., Туркова С.О., Орлова А.Р., Сулимова Г.Е. Генетические механизмы устойчивости и чувствительности к лейкозу у айрширской и черно-пестрой пород крупного рогатого скота, установленные на основе распределения аллелей гена BoLA-DRB3//Генетика. 2003. Т. 39. С.383-396.
- Beever J.E., Lewin H.A., Barendse W. et al. Report of the first workshop on the genetic map of bovine chromosome 23//Anim. Gen. 1996. V. 27. N 2. P. 69-75.
- Leprovost F. Prolactin gene expression in ovine and caprine mammary gland//Neuroendocrinology. 1994. V. 60. P. 305-313.
- Carlos Souza do Nascimento, Marco Antonio Machado, Mario Luiz Martinez. Association of the bovine major histocompatibility complex (BoLA) BoLA-DRB3 gene with fat and protein production and somatic cell scorein Brazilian Gyr dairy cattle (Bos indicus)//Gen. Mol. Biol. 2006. V. 29. № 4. P. 641-647.
