Анализ сцепления гена устойчивости к расе G заразихи с микросателлитными локусами у линии-донора подсолнечника селекции ВНИИМК RGP1

Автор: Гучетль С.З., Савиченко Д.Л.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (186), 2021 года.
Бесплатный доступ
Одним из главных биотических факторов, лимитирующих получение высоких урожаев подсолнечника, является заразиха (Orobanche cumana Wallr.). Наиболее эффективным и экологически безопасным методом борьбы считается возделывание устойчивых сортов и гибридов подсолнечника. Создание резистентных генотипов включает в себя поиск и использование источников устойчивости в селекционном процессе, а также обеспечение точных и производительных процедур оценки материала. Целью исследования являлся анализ сцепления гена Or7 с микросателлитными локусами у линии-донора устойчивости к расе G заразихи коллекции ВНИИМК. Материал исследования - линия-донор устойчивости к расе G заразихи RGP1 и восприимчивая к этой расе линия ВК678 селекции ВНИИМК. Растения подсолнечника были скрещены в полевых условиях для получения поколения F1. Также проведено самоопыление растений F1 с получением потомства F2. Растения тестировали в теплице методом ранней диагностики на инфекционном фоне из семян заразихи расы G. ДНК подсолнечника экстрагировали из верхушечных листьев молодых побегов вегетирующих растений. Для ПЦР анализа использовали три SSR-праймера, показавшие полиморфизм у родительских линий: ORS 683, ORS 1040, ORS 1112. Был проведен тест на совместное наследование гена Or7 c этими локусами, а также SSR-локусов друг с другом. Показано независимое наследование гена Or7 c локусами ДНК ORS 683, ORS 1040, ORS 1112, а также SSR-локусов ORS 1040 с ORS 1112 и ORS 1040 с ORS 683. Локусы ORS683 - ORS 1112 сцеплены с частотой рекомбинации 0,27 ± 0,41 (27 сМ). В результате проведенных исследований было исключено расположение гена Or7 в области непосредственной близости к микросателлитным локусам ORS 683, ORS 1040, ORS 1112. На основании изученных литературных источников и репрезентативного генома подсолнечника HanXRQr2.0-SUNRISE составлена частичная физическая карта LG3 для определения области дальнейшего поиска локализации гена Or7 и ДНК-маркеров, косегрегирующих с ним.
Подсолнечник, заразиха, раса G, устойчивость, микросателлитные локусы, ПЦР, сцепление
Короткий адрес: https://sciup.org/142229254
IDR: 142229254 | УДК: 633.854.78:575:632.9 | DOI: 10.25230/2412-608X-2021-2-186-3-9
A linkage of gene of resistance to a broomrape race G with microsatellite loci of a sunflower line-donor RGP1 of VNIIMK's breeding
Broomrape (Orobanche cumana Wallr.) is one of the main biotic factors limiting high sunflower yield formation. The most effective and environmentally safe method of protection is cultivation of resistant varieties and hybrids of sunflower. Development of resistant sunflower genotypes includes search and usage of sources of resistance in breeding process as well as accurate and productive procedures of material assessment. The purpose of the research is to analyze a linkage of a gene Or7 with microsatellite loci of the line-donor of resistance to broomrape race G from the VNIIMK’s collection. The objects of the research are the line RGP1 - a donor of resistance to broom-rape race G and a susceptible to this race line VR 678 from the VNIIMK’s collection. Sunflower plants were crossed in field to produce F1. Also we conducted self-pollination of F1 plants to obtain F2 progeny. Plants were tested in a greenhouse in soil infected with seeds of broomrape race G using a method of early diagnostic. Sunflower DNA was extracted from the top leaves of the young sprouts of the vegetative plants. For PCR-analysis we used three SSR-primers demonstrated polymorphism in parental lines: ORS 683, ORS 1040, and ORS 1112. We tested joint inheritance of the gene Or7 and these loci, and inheritance between SSR-loci. An independent inheritance of the gene Or7 with DNA-loci ORS 683, ORS 1040, and ORS 1112, as well as SSR-loci between ORS 1040 and ORS 1112, ORS 1040 and ORS 683 was showed. Loci ORS683 - ORS 1112 are linked with a frequency of recombination of 0.27 ± 0.41 (27 cM). As a result of our research location of the gene Or7 in the nearest area to microsatellite loci ORS 683, ORS 1040, and ORS 1112 was excluded. Basing on studied literary sources and a representative sunflower genome HanXRQr2.0-SUNRISE we made a partial physical map LG3 for determination of an area for the further search of a localization of the Or7 and DNA-markers cosegregating with this gene.
Текст научной статьи Анализ сцепления гена устойчивости к расе G заразихи с микросателлитными локусами у линии-донора подсолнечника селекции ВНИИМК RGP1
Введение. Одним из главных биотических факторов, лимитирующих получение высоких урожаев подсолнечника, является паразитическое растение заразиха ( Orobanche cumana Wallr.). При отсутствии резистентности у возделываемых гибридов ущерб урожаю, наносимый данным паразитом, может составить до 100 % [1]. На данный момент сообщается об обнаружении, по крайней мере, восьми рас, обозначенных буквами от А до Н в порядке возрастания вирулентности [2].
Наиболее эффективным и экологически безопасным методом борьбы с заразихой считается возделывание устойчивых сортов и гибридов подсолнечника. Создание резистентных генотипов включает в себя поиск и использование источников устойчивости в селекционном процессе, а также обеспечение точных и производительных процедур оценки материала. Учитывая, что устойчивость подсолнечника быстро преодолевается с появлением более вирулентных рас паразита, необходимо непрерывно вести процесс внедрения и комбинирования генов устойчивости [3; 4].
Устойчивость подсолнечника к расам заразихи контролируется различными способами: одним доминантным [5], од-4
ним или двумя рецессивными и двумя частично доминантными аллелями генов [4; 6]. Это говорит о наличии различных источников устойчивости, обусловленных происхождением генетического материала. Тем самым для обеспечения оценки селекционного материала с помощью ДНК-маркеров (маркер-ассо-циированной селекции) необходимо картировать ген устойчивости для каждого источника. Пять локусов количественных признаков (QTL), обеспечивающих устойчивость к расе E, и шесть QTL – к расе F, были обнаружены в семи разных хромосомах [5]. Расоспецифичный ген, контролирующий устойчивость к расе E – Or 5 картирован в теломерной области хромосомы 3, находится на 7,5 сМ выше маркера ORS1036 [7]. В той же хромосоме был обнаружен ген устойчивости к расе выше F, который предварительно обозначен как or ab-vl-8 и локализован на 1,5 сМ ниже маркера ORS683 и 19,6 сМ ниже маркера ORS1036 [4]. Несмотря на близкое расположение к гену Or 5 , доказано, что эти гены различны. Позже, I. Ime-rovski et al. [8] картировали от 2 до 23 значимых QTL в геноме подсолнечника. Два основных QTL располагались в хромосоме 3. Так же один ген устойчивости к расе F O. cumana был описан и картирован в хромосоме 7 [9].
Во ВНИИМК были созданы линии подсолнечника, устойчивые к расе G заразихи. Установлено, что устойчивость у этих линий контролируется одним геном Or 7 с неполным доминированием [10]. Эти линии представляют ценный источник для переноса генов с использованием ДНК-маркеров (МАС). Использование данных генов в селекции позволит создавать безопасные и качественные продукты питания с использованием современных генетических методов. В связи с этим целью исследования является анализ сцепления гена Or 7 с микросателлитными локусами у линии-донора устойчивости к расе G заразихи RGP1 коллекции ВНИИМК.
Материалы и методы. Материал исследования – линия-донор устойчивости к расе G заразихи RGP1 и восприимчивая к этой расе линия ВК678 селекции ВНИИМК. Растения подсолнечника были скрещены в полевых условиях для получения гетерозиготного поколения F 1. Затем проведено самоопыление растений F 1 с получением потомства F 2. Растения тестировали в теплице на устойчивость и восприимчивость на инфекционном фоне из семян заразихи расы G, применяя метод ранней диагностики [10].
ДНК подсолнечника экстрагировали из верхушечных листьев молодых побегов вегетирующих растений. Выделение ДНК производилось по методу Vroh et аl. с добавлением активированного угля [11]. Для проведения полимеразной цепной реакции использовали 25 мкл реакционной смеси следующего состава: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфата аммония; 1,5–3 мM MgCl 2 ; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозидфосфатов; по 10 пМ праймеров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК полимеразы (Си-бэнзим, Россия). Для амплификации использовали термоциклер S1000тм (BioRad, США). Для увеличения специфичности гибридизации праймеров с матрицей ДНК использовали ступенчатую ПЦР (touchdown PCR). Условия амплификации: начальная денатурация 95 °C в течение 2 мин, 1 цикл при соблюдении температурно-временного режима: 94 °C – 30 сек, 64 °C – 30 сек, 65 °C – 45 сек, затем шесть циклов с уменьшением температуры отжига на 1 °C на цикл, следом 32 цикла при соблюдении температурновременного режима: 94 °C – 30 сек, 58 °C – 30 сек и 65 °C – 45 сек. Финальная элонгация 65 °C – 20 мин. Для ПЦР анализа использовали три SSR-праймера, показавшие полиморфизм у родительских линий в предыдущем исследовании: ORS 683, ORS 1040, ORS 1112 [12].
Электрофорез продуктов амплификации проводили в полиакриламидном геле
(8 %, 1хТБЕ) с использованием камеры VE-20 для вертикального электрофореза (Хеликон, Россия) в течение 2,5–3 часов при силе тока 40–50 mA и напряжении 200–230 V. Результаты электрофореза документировали при помощи гель-доку-ментирующей видеосистемы BIO-PRINT (Vilber Lourmat, Франция). Размер фрагментов ДНК определяли с использованием программного обеспечения BioCapture (Vilber Lourmat, Франция) относительно маркера длины фрагментов ДНК GeneRuler 100 bp DNA Ladder Thermo Scientific (Сибэнзим, Россия). Математическую обработку результатов расщепления проводили с использованием χ2-критерия соответствия фактических расщеплений теоретически ожидаемым в моно- и дигибридных скрещиваниях. Расчет частоты рекомбинации r и ошибку рекомбинации s p по результатам расщепления в F 2 рассчитывали по методу максимального правдоподобия [13].
Результаты и обсуждение. Для установления локализации гена, контролирующего устойчивость к расе G заразихи, I. Imerovski et al. была составлена частичная SSR карта LG3 подсолнечника, содержащая ген or ab-vl-8 . Согласно карте, данный ген локализуется в верхней части хромосомы [4]. С использованием BSA-seq I. Imerovski et al . также были обнаружены предполагаемые гены-кандидаты устойчивости – HanXRQChr03g0076321 и HanXRQChr03g0065841 [8]. Основываясь на полученных этими авторами результатах, мы проводили подбор генетических маркеров к гену Or 7 [12]. В рамках настоящего исследования используемые в работе SSR-локусы были сопоставлены со сборкой репрезентативного генома подсолнечника HanXRQr2.0-SUNRISE. По этим данным с помощью инструмента Basic Local Alignment Search Tool (BLAST®) составлена физическая карта положения локусов в LG3 (рисунок).
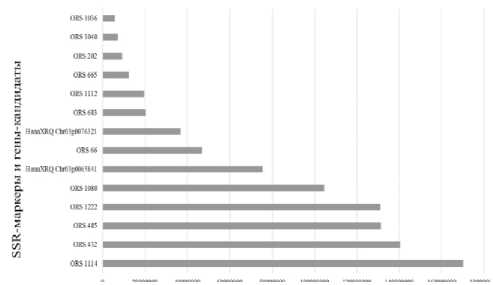
Таблица 1
Аллельные состояния SSR-локусов ДНК подсолнечника у гибрида F 1 (ВК678 × RGP1) и его родительских линий, восприимчивой и устойчивой к расе G заразихи
Рисунок – Положение SSR-локусов, предположительно сцепленных с геном Or 7 , и генов-кандидатов в LG3 H. annuus
Согласно этой карте, в верхней части LG3 располагается группа локусов (ORS 683, ORS 1112, ORS 1040, ORS 665, ORS 202, ORS 1036), находящихся выше предполагаемого нахождения гена устойчивости к расе G заразихи (рисунок).
Для Or 7 у были RGP1,
установления локализации гена линий селекции ВНИИМК нами выбраны линии подсолнечника устойчивая к расе G заразихи, и
ВК 678, восприимчивая к этой расе. У этих линий обнаружено три маркерных микросателлитных локуса с контрастными аллельными вариантами [12]. Это локусы ORS 1040, ORS 1112, ORS 683. Полученные после скрещивания родительских линий семена подсолнечника F 1 (ВК678 × RGP1) были проверены на гиб-ридность по ДНК локусам. По результатам анализа было установлено, что два локуса – ORS 683, ORS 1112 – имели кодоминантное наследование аллелей. По этим локусам у гибрида выявили две родительские фракции ДНК. Представляет интерес тот факт, что I. Imerovski et al. [4] определили эти локусы, как локусы с доминантным наследованием аллелей. У локуса ORS 1040 выявлена одна фракция, следовательно, имеет место доминирование одного аллеля над другим (табл. 1).
После получения потомства F 2 анализ расщепления по устойчивости подсолнечника к расе G заразихи для данной комбинации скрещивания показал, что фактически наблюдаемое расщепление соответствовало теоретически ожидаемой модели 3 : 1 при моногенном наследовании признака (табл. 2).
Таблица 2
Наследование признака устойчивости подсолнечника к расе G заразихи в F 2 при скрещивании линий ВК678 и RGP1
|
Ген |
Всего растений, шт. |
Теоретически ожидаемое соотношение |
χ2 |
df |
Р |
|
Or 7 |
107 |
3 : 1 |
0,24 |
1 |
0,7–0,8 |
Ранее было установлено, что у линии подсолнечника RGP1 устойчивость обеспечивается действием одного гена Or 7 с доминантным типом наследования [10]. Наши исследования также подтвердили этот тип наследования у данного генотипа.
Также был проведен гибридологический анализ результатов расщепления в F 2 для трех ДНК-локусов. Локусы ORS 683 и ORS 1112 являются кодоминантными, и во втором поколении расщепление соответствует модели 1 : 2 : 1, а у доминантного локуса ORS 1040 – модели 3 : 1 (табл. 3).
Таблица 3
Наследование SSR-локусов ДНК ORS 683, ORS 1040 и ORS 1112 при скрещивании линий ВК678 и RGP1
|
Локус |
Всего растений, шт. |
Теоретически ожидаемое соотношение |
χ2 |
df |
Р |
|
ORS 683 |
98 |
1:2:1 |
1,16 |
2 |
0,56 |
|
ORS 1112 |
94 |
1:2:1 |
2,37 |
2 |
0,67 |
|
ORS 1040 |
102 |
3:1 |
3,84 |
1 |
0,05 |
Для данных локусов χ2-критерий достоверно подтверждал соответствие фактических расщеплений теоретически ожидаемым. Следовательно, комбинация скрещивания пригодна для дальнейшего анализа. Проведен тест на совместное наследование гена Or 7 c этими локусами, а также SSR-локусов друг с другом (табл. 4).
Таблица 4
Значения χ2 между геном Or 7 и SSR-локусами ДНК в потомстве F 2 (ВК 678 × RGP1)
|
Расщепляющиеся локусы |
Всего растений, шт. |
Теоретически ожидаемое соотношение |
χ2 |
df |
Р |
|
Or 7 –ORS 683 |
98 |
3:6:3:1:2:1 |
4,96 |
5 |
0,08 |
|
Or 7 – ORS 1112 |
89 |
3:6:3:1:2:1 |
7,42 |
5 |
0,19 |
|
Or 7 – ORS 1040 |
102 |
9:3:3:1 |
7,27 |
3 |
0,06 |
|
ORS 683 – ORS 1112 |
94 |
1:2:1:2:4:2:1:2:1 |
19,2 |
8 |
< 0,05 |
|
ORS 1040 – ORS 683 |
97 |
3:6:3:1:2:1 |
8,33 |
5 |
0,15 |
|
ORS 1040 – ORS 1112 |
88 |
3:6:3:1:2:1 |
2,06 |
5 |
0,30 |
В таблице 4 представлены данные по оценке совместного наследования гена Or 7 c ДНК локусами, где отмечено значение χ2, которое характеризует отклонение от независимого расщепления приведенных локусов. Показаны также вероятность нулевой гипотезы о независимом наследовании (P) между анализируемыми генами. Тест на совместное наследование показал независимое наследование гена Or 7 c локусами ДНК ORS 683, ORS 1040, ORS 1112, а также SSR-локусов ORS 1040 с ORS 1112 и ORS 1040 с ORS 683. Независимое наследование пары локусов ORS683 – ORS 1112 не подтвердилось (χ2 = 19,2; Р <0,05) (табл. 4).
Была выполнена оценка частоты рекомбинации между парами локусов Or 7 – ORS 683, ORS 1040 – ORS 1112, ORS 683 – ORS 1112 (табл. 5).
Таблица 5
Значения частот рекомбинации между геном Or 7 и SSR-локусами ДНК в потомстве F 2 (ВК678 х RGP1)
|
Расщепляющиеся локусы |
r |
s p |
|
Or 7 – ORS 683 |
0,39 |
0,06 |
|
ORS 683 – ORS 1112 |
0,27 |
0,41 |
|
ORS 1040 – ORS 1112 |
0,56 |
0,60 |
Тест подтвердил сцепление локусов ORS 683 – ORS 1112 с частотой рекомбинации 0,27 ± 0,41 (27 сМ). Частота рекомбинации локусов Or 7 – ORS 683 составила 0,39 ± 0,06 (39 сМ). Вероятно, сцепление между генами отсутствует или очень слабое. Коэффициент рекомбинации между локусами ORS 1040 – ORS 1112 составил более 50 %, что говорит об отсутствии сцепления между этими локусами. Мы предполагаем следующий порядок расположения генов в данном сегменте хромосомы подсолнечника LG3: Or 7 – ORS 683 – ORS 1112 – ORS 1040 с расстояниями между ними 39, 27 и 56 сМ соответственно. Такой порядок расположения локусов согласуется с их расположением на SSR [4] и физической карте, сконструированной нами. Однако полученные данные частот рекомбинации между исследуемыми генами не совпадают с результатами исследований других авторов. Так, согласно I. Imerovski et al. [4], SSR-локус ORS 683 находится на расстоянии 1,5 сМ выше гена or ab-vl-8 . У этих же авторов дистанция между ORS 683 и ORS 1112 составляет 9,8 сМ. Tang et al. [7] определили дистанцию между ORS 683 и ORS 1112 в 0,9 сМ, а между ORS 683 и ORS 1040 – 7 сМ. Этими авторами было отмечено, что на оценку расстояния между локусами могут оказать влияние фенотипические ошибки, потому что идентификация восприимчивого фенотипа как устойчивого при классификации приводит к увеличению класса рекомбинантных генотипов и искажает дистанции на генетической карте [7]. Кроме того, оценка частоты рекомбинации зависит от генотипов, которые используются для анализа [14].
Вероятно, ген, который контролирует устойчивость к расе G заразихи у линии RGP1, не является идентичным гену orab-vl-8, описанному сербскими учеными. Исходя из литературных данных, мы предполагаем, что он локализуется в этой же хромосоме ниже области, в которой производился поиск. Предложенные гены-кандидаты располагаются также в области ниже маркера ORS 683 [8]. В связи с этим будет продолжена работа по поиску местонахождения гена Or7 и локусов ДНК, сцепленных с ним, в нижних областях LG3.
Выводы. В результате проведенных исследований было исключено расположение гена Or 7 в области непосредственной близости к микросателлитным локусам ORS 683, ORS 1040, ORS 1112. Локусы ORS 683 – ORS 1112 показали сцепление с частотой рекомбинации 0,27 ± 0,41 (27 сМ). На основании изученных литературных источников и репрезентативного генома подсолнечника HanXRQr2.0-SUNRISE составлена частичная физическая карта LG3 для определения области дальнейшего поиска локализации гена Or 7 и ДНК-маркеров, косегрегирующих с ним.
Список литературы Анализ сцепления гена устойчивости к расе G заразихи с микросателлитными локусами у линии-донора подсолнечника селекции ВНИИМК RGP1
- Molinero-Ruiz L., Delavault P., Pérez-Vich B., Pacureanu-Joita M., Bulos M., Altieri E., Domínguez J. History of the race structure of Orobanche cumana and the breeding of sunflower for resistance to this parasitic weed // A review. J. Agric. Res. - 2015. - V. 13.- e10R01 DOI: 10.5424/sjar/2015134-8080.
- Martín-Sanz A., Malek J., Fernández-Martínez J., Pérez-Vich B., Velasco L. Increased virulence in sunflower broomrape (Orobanche cumana Wallr.) populations from Southern Spain is associated with greater genetic diversity // Front. Plant Sci. - 2016. - V. 7. - P. 589. DOI: 10.3389/fpls.2016.00589.
- Fernández-Martínez J., Pérez-Vich B., Ve-lasco L. Sunflower broomrape (Orobanche cu-mana Wallr.) // Sunflower (Urbana: AOCS Press) 2015. - P. 129-155.
- Imerovsk, I., Dimitrijevic A., Miladinovic D., Dediv B., Jocic S., Kocis Tubic N., Cvejic S. Mapping of a new gene for resistance to broom-rape races higher than F // Euphytica. - 2016. -V. 209. - P. 281-289.
- Pérez-Vich B., Akhtouch B., Knapp S., Leon A., Velasco L., Fernández-Martínez J., Berry S.T. Quantitative trait loci for broomrape (Oro-banche cumana Wallr.) resistance in sunflower // Theor. Appl. Genet. - 2004. - V. 109. - P. 92102.
- Akhtouch B., del Moral L., Leon A., Velasco L., Fernández-Martínez J., Pérez-Vich B. Genetic study of recessive broomrape resistance in sunflower // Euphytica. - 2016. - V. 209. - P. 419-428.
- Tang S., Heesacker A., Kishore V., Fernandez A., Sadik E., Cole G., Knapp S. Genetic mapping of the Or5 gene for resistance to Orobanche race E in sunflower // Crop Sci. - 2003. -V. 43. - P. 1021-1028.
- Imerovsk I., Dedic B., Cvejic S., Miladinovic D., Jocic S., Owens G., Tubic N., Rieseberg L. BSA-seq mapping reveals major QTL for broomrape resistance in four sunflower lines // Mol. Breeding. - 2019. - V. 39. - P. 3941. DOI.org/10.1007/s11032-019-0948-9.
- Duriez P., Vautrin S., Auriac M., Bazerque J., Boniface M., Callot C., Carrére S., Cauet S., Chabaud M., Gentou F. A receptor-like kinase enhances sunflower resistance to Orobanche cumana // Nat. Plants. - 2019. - V. 5. - P. 12111215.
- Гучетль С.З., Арасланова Н.М., Антонова Т.С., Челюстникова Т.А., Питинова Ю.В. Генетический анализ устойчивости к расе G Orobanche cumana Wallr. в F2 и ВС1 линий подсолнечника RGP1, RGP2, RGB, RGL1, RGL2 // Масличные культуры. - 2019. -Вып. 4 (180). - С. 23-28. DOI: 10.25230/2412-608Х-2019-4-180-23-28.
- Vroh Bi I., Harvengt L., Chandelier A., Mergeai P. Improved RAPD amplification of recalcitrant plant DNA by the use of activated charcoal during DNA extraction // Plant breeding. - 1996. - Vol. 115. - Is. 3 - P. 205-206.
- Савиченко Д.Л., Гучетль С.З., Голо-ватская А.В. Поиск и анализ нуклеотидных последовательностей в геноме подсолнечника, ассоциированных с генами устойчивости к расам заразихи Orobanche cumana Wallr. // Масличные культуры. - 2020. - Вып. 3 (183). -С. 11-19. DOI: 10.25230/2412-608X-2020-3-183-11-19.
- Гершензон С.М. Основы современной генетики. - Киев: Наукова думка, 1979. - 508 с.
- Карабицина Ю.И., Гаврилова В.А., Ал-патьева Н.В., Кузнецова Е.Б., Анисимова И.Н. Особенности наследования признака восстановления фертильности пыльцы подсолнечника при цитоплазматической мужской стерильности // Генетика. - 2019. - Т. 55. - № 11. - C.1327-1334.
