Анатомо-морфологические особенности листовых органов некоторых видов рода Astragalus L. в их онтогенезе

Автор: Маматкулов О.И., Коланов О., Эркебаева Ж.Н.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 10 т.9, 2023 года.
Бесплатный доступ
Актуальность: изучение анатомического строения листовых органов растений рода Astragalus L. в их онтогенезе является актуальным в связи с тем, что результаты исследования могут быть успешно использованы при решении систематических и эволюционных вопросов таксономических категорий высших растений. Цели исследования: изучить количественные показатели признаков анатомического строения листовых органов в онтогенезе у 5 видов рода Astragalus L., относящиеся к разным биоморфам: А. filicaulis Fisch. et C. A. Mey. ex Ledeb. (однолетник), А. campylorrhynchus Fisch. et C. A. Mey. (однолетник), А. rhacodes Bunge (многолетник), А. dendroides Kar. et Kir. (кустарничек) и А. fedtschenkoanus Lipsky (кустарник) с целью установления их анатомо-морфологических особенностей, при помощи общепринятых онтогенетических и статистических методов исследования растительных объектов. Материалы и методы исследования: виды рода Astragalus L., относящиеся к разным биоморфам были посажены в почву, без какой-либо предварительной обработки. При уходе за ростом и развитием растений и учете количественных показателей морфологических и анатомических структур листовых органов и их статистической обработке использовались общепринятые методы. Результаты исследования: виды отличаются друг от друга некоторыми количественными показателями строения листовых органов, как размерами эпидермальных клеток, числом устьичных аппаратов на единицу площади, высотой верхних и нижних эпидермальных клеток, числом слоев клеток мезофилла и общей толщиной листовой пластинки. Выводы: общими для изученных видов являются, что листовые органы в онтогенезе имеют амфистоматную структуру.
Вид, онтогенез, семядольный лист, ювенильный лист, дефинитивный лист, эпидерма, устьица, волоски, мезофилл
Короткий адрес: https://sciup.org/14128645
IDR: 14128645 | УДК: 581.4:581.84 | DOI: 10.33619/2414-2948/95/03
Anatomic-morphological structure features of leaf organs of some species of the genus Astragalus L. in their ontogenesis
Research relevance: anatomical structure study of Astragalus L. genus plants leaf organs in their ontogenesis is relevant for the reason that results obtained after study can be successfully used in solving systematic and evolutionary issues of higher plants taxonomic categories. Research objectives: to study quantitative indicator characteristics of anatomical structure of leaf organs ontogenesis in 5 species of the Astragalus L., genus belonging to different biomorphs: A. filicaulis Fisch. et C. A. Mey. ex Ledeb. (annual), A. campylorrhynchus Fisch. et C. A. Mey. (annual), A. rhacodes, A. rhacodes Bunge (perennial), A. dendroides Kar. et Kir. (shrub) and A. fedtschenkoanus Lipsky (shrub) in order to establish their anatomical and morphological features, using generally accepted ontogenetic and statistical methods. Research materials and methods: Astragalus L. genus species belonging to different biomorphs planted in the soil without any pre-treatment. In plants growth care and development quantitative indicators of the morphological and anatomical structures of leaf organs and their statistical processing, generally accepted methods were used. Research results: species differ from each other in some quantitative indicators of the leaf organs structure, such as the epidermal cells size, the number of stomatal apparatus per unit area, the height of the upper and lower epidermal cells, the number of layers of mesophyll cells and the total thickness of the leaf blade. Conclusions: common feature to the studied species is that the leaf organs in ontogenesis have an amphistomatous structure.
Текст научной статьи Анатомо-морфологические особенности листовых органов некоторых видов рода Astragalus L. в их онтогенезе
Бюллетень науки и практики / Bulletin of Science and Practice
Изучение онтогенетических особенностей каждой из таксономических категорий высших растений дает довольно достоверные данные, которые надежно служат при решении систематических, экологических и эволюционных вопросов. Анатомо-морфологические данные индивидуального развития отдельной особи являются основными при установлении путей приспособительной эволюции тех или иных таксонов и таксономических групп [1–8]. Цель работы: изучить анатомо-морфологические особенности некоторых видов рода Astragalus L., представляющие разные жизненные формы. Задачи работы:
-
1) сбор и подготовка семян к посеву в условиях естественного произрастания и опыта;
-
2) проведение фенологического наблюдения за ростом и развитием растений, путем замера размеров органов, фиксации на различных этапах развития и изучения их микроскопической структуры;
-
3) произвести сравнительный анализ структурных особенностей листовых органов на различных этапах развития.
Материалы и методы исследования
Семенной материал был посажен в почву, без какой-либо предварительной обработки. При уходе за ростом и развитием растений и учете количественных показателей морфологических и анатомических структур листовых органов и их статистической обработке использовались общепринятые методы [9]. Для изучения волосков, устьиц, эпидермальных клеток, жилкования, материал просветляли по методике М. П. Пахомовой [10], очертание клеток эпидермы описали по С. Ф. Захаревичу [11]. Анатомическое исследование изученных листовых органов и подсчет их количественных показателей проводили в 10-кратной повторности.
Результаты и обсуждение
Всходы у изученных видов в лабораторных условиях появляются через 20–35 дней после посева. У однолетних видов для прорастания семян требуется 20–27 дней, а у многолетних трав и кустарников — 25–35 дней. Прорастание семян у всех видов гипокотилярное. Семядольные листья проростков растут очень медленно в течение от 7–9 дней ( A. campylorrhynchus ) до 25–30 дней ( A. dendroides и A. fedtschenkoanus ), т. е. их размеры очень мало изменяются с момента распускания. Форма семядольных листьев у однолетних видов удлиненно-яйцевидные. Что касается многолетних трав и кустарников, то они у A. rhacodes — яйцевидные, у A. dendroides — обратнояйцевидные, а у A. fedtschenkoanus — удлиненно-обратнояйцевидные. Здесь необходимо отметить наличие некоторой корреляции между формами и длиной листовой пластинки семядольных листьев: так, у однолетних они довольно короткие — 5–6 мм, а у многолетних трав и кустарников заметно длинные. Что касается ширины листовой пластинки семядолей, то изученные виды резко не отличаются друг от друга (Таблица).
Таблица
МОРФОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ ЛИСТОВЫХ ОРГАНОВ В РАЗНЫХ ЭТАПАХ ОНТОГЕНЕЗА У НЕКОТОРЫХ ВИДОВ РОДА Astragalus L. (Fabaceae)
|
виды Astragalus L. |
||
|
A. filicaulis |
A. campylorrhynchus A. rhacodes A. dendroides |
A. fedtschenkoanus |
|
Появление всходов после посева, дни |
||
|
23–27 |
20–25 25–30 30–35 |
25–30 |
|
Тип прорастания |
||
|
гипокотилярное |
гипокотилярное гипокотилярное гипокотилярное |
гипокотилярное |
|
Форма семядольных листьев |
||
|
удлиненнояйцевидная |
удлиненно- неравнобокая, неравнобокая, яйцевидная яйцевидная обратнояйцевидная |
удлиненная, неравнобокая, обратнояйцевидная |
|
Размеры семядолей: длина/ширина, мм |
||
|
5–6/3–4 |
5–6/4–5 7–8/4–5 6–8/ 4–6 |
9–11/4–6 |
|
Появление первых настоящих листьев, дни |
||
|
10–12 |
11–13 11–14 12–18 |
13–18 |
|
Форма первых настоящих листьев |
||
|
простая, сердцевидная |
простая, простая, удлиненно- тройчатая сердцевидная яйцевидная |
тройчатая |
|
Размеры первых настоящих листьев: длина/ ширина/ длина черешка, мм |
||
|
6–9/4–5/16–20 |
6–8/4–5/20–22 14–16/5–6/20–24 14–19/10–12/10–14 |
16–22/8–10/18–20 |
|
Размер дефинитивных листьев: длина/ширина, мм |
||
|
40–70/10–12 |
30–100/30–40 70–140/30–40 30–70/20–25 |
30–60/20–25 |
|
Количество листочков в одном сложном листе, шт |
||
|
9–13 листочковые |
7–9 листочковые 4–15 листочковые 3–5 (7) листочковые |
7(9) листочковые |
|
Форма листочков |
||
|
продолговатые, овальнопродолговатые или линейные |
линейные или продолговато- овальные или нитевидно- обратнояйцевидные эллиптические линейные или округло обратнояйцевидные |
продолговатоэллиптические |
|
Размеры листочков сложных дефинитивных листьев молодых растений: длина/ширина, мм |
||
|
6–8/3–4 |
8–14/6–8 6–25/3–4 8–17/5–6 |
6–13/3–5 |
Первые настоящие листья у всех изученных видов начинают формироваться через 10– 18 дней после появления проростков. В процессе индивидуального развития растений наблюдается морфологическая гидрофилия, которая выражается в виде смены простых листьев на сложные: в начале на тройчатые и, далее на непарноперистые листья по мере продвижения вверх по главному побегу. Первые листья у травянистых форм простые, сердцевидной и удлиненно-яйцевидной формы, у кустарниковых форм — тройчатые. Изученные виды по количественным показателям дины и ширины листовой пластинки четко подразделяются на две группы: они у однолетних трав заметно короткие и узкие, чем у многолетних трав и кустарников, где листовые пластинки почти на 2–3 раза длиннее и 2 раза шире, чем таковые у первой группы. У всех изученных видов черешки первых настоящих листьев довольно длинные, от 16–24 мм, за исключением A. dendroides , у которого они заметно короткие (10–14 мм).
У всех изученных видов второй лист главного побега формируется через 10–12 дней после появления первого настоящего листа. Далее синхронное появление третьих и последующих листьев главного побега составляет в пределах от 7 до 9 дней включительно.
У A. rhacodes листья главного побега до 5–6 (8) узла простые, далее вверх появляются непарноперистосложные листья с 3–5–7 листочками. У A. filicaulis и A. campylorrhynchus листья главного побега, начиная со второго узла сложные, тройчатые, у первого с третьего узла формируются непарноперистосложные листья с 6–7 парами листочков, а у второго тройчатые листья располагаются до 5–6 узла главного побега включительно. Далее вверх по главному побегу располагаются непарноперистосложные листья с 6–7 парами листочков. У третьей группы все настоящие листья с первого узла сложные: они у A. fedtschenkoanus со второго узла непарноперистосложные с 5–7 (9) листочками, а у A. dendroides тройчатосложные листья располагаются с 1 по 6 узла, далее вверх по главному побегу развиваются непарноперистосложные листья с 5 (7) листочками. Наши данные в той или иной мере согласуется с мнением некоторых исследователей, которые полагали, что у молодых по генезису астрагалов (например, виды-однолетники) часто наблюдаются сложные (перистые) первые листья, слабо охваченные редукцией. Это явление может найти свое объяснение в эфемерном образе жизни таких видов: развиваясь в условиях повышенного увлажнения (весною), они уходят от воздействия неблагоприятных (прежде всего температурных) условий [3].
Эпидермальные клетки семядольных листьев у изученных видов изодиаметрические, удлиненные и удлиненно-изодиаметрические. Они на верхней эпидерме (ВЭ) более или менее крупные, чем на нижней эпидерме (НЭ). Исключение составляет только кустарничек A. dendroides , у которого, наоборот, клетки нижней эпидермы почти в два раза крупнее, чем размеры верхней эпидермы. По размерам клеток верхней эпидермы изученные виды подразделяются на две группы: у однолетнего A. campylorrhynchus и кустарника A. fedtschenkoanus они очень крупные, а у остальных — относительно мелкие и 2,0–2,2 раза меньше, чем у первой группы. Что касается нижней эпидермы, то у однолетнего A. filicaulis они мелкие, у остальных видов более или менее крупные. В целом эпидермальные клетки семядолей у однолетнего A. campylorrhynchus очень крупные, а у другого однолетника A. filicaulis они относительно мелкие, остальные виды по этому показателю занимают промежуточное положение между этими таксонами.
Семядольные листья у изученных видов Astragalus L. амфистоматные. Устьичные аппараты аномоцитного и анизоцитного типов. У однолетних A. filicaulis, A. campylorrhynchus и кустарничка A. dendroides число устьиц на единицу площади на верхней эпидерме относительно больше, чем таковые на нижней эпидерме. Что касается многолетника A. rhacodes Bunge, то они на верхней и нижней эпидерме почти в одинаковом количестве, а у кустарника A. fedtschenkoanus, наоборот, их число на нижней эпидерме почти 2,5 раза больше, чем таковые на верхней эпидерме (Рисунок 1).
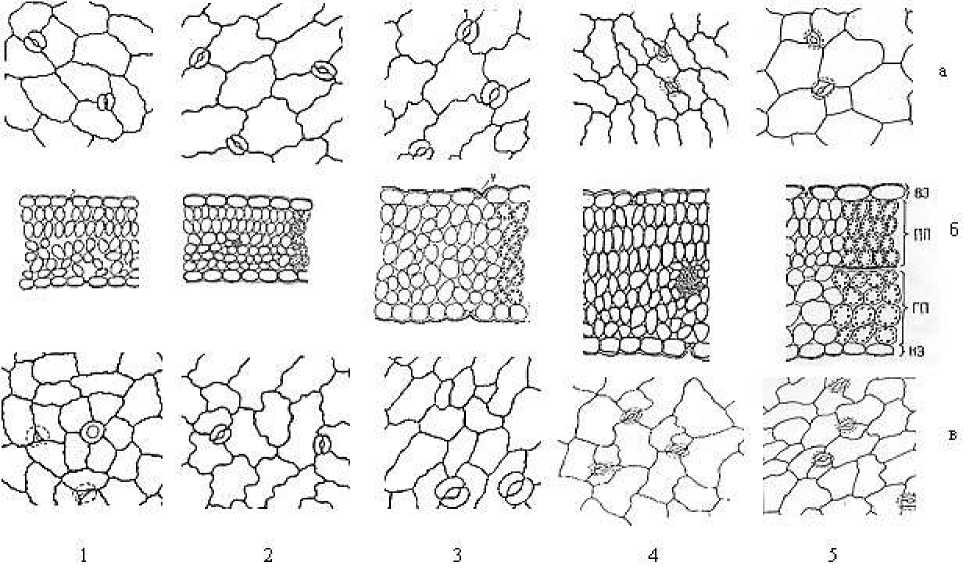
Рисунок 1. Анатомическое строение зрелых семядольных листьев некоторых видов рода Astragalus L. 1 — A. filicaulis , 2 — A. campylorrhynchus , 3 — A. rhacodes , 4 — A. fedtschenkoanus , 5 — A. dendroides . а — верхняя эпидерма, б — поперечный срез листовой пластинки, в — нижняя эпидерма
Изученные виды рода Astragalus по толщине пластинок семядольных листьев подразделяются на три группы. Первую группу составляют однолетники A. filicaulis, A. campylorrhynchus. Они имеют относительно тонкую пластинку. Многолетник A. rhacodes характеризуется довольно тонкой пластинкой и составляет отдельную, вторую группу. Растения с одревесневшими стеблями, имеющие жизненные формы кустарнички и кустарники A. dendroides и A. fedtschenkoanus обладают очень толстыми семядольными листьями, и составляют третью группу. Виды почти не отличаются друг от друга по толщине верхних и нижних эпидермальных клеток, и имеют показатели в пределах от 22,6±0,1 мкм до 32,84±1,8 мкм. У A. rhacodes мезофилл листовой пластинки не дифференцирован на палисадную и губчатую паренхиму, и имеет однородную структуру. У остальных видов мезофиллы листовых пластинок обладают дорсовентральной структурой. У однолетних видов ( A. filicaulis и A. campylorrhynchus ) верхняя палисадная паренхима (ВПП) состоит из одного слоя столбчатых клеток и имеет относительно небольшую толщину (50±0,2 мкм). А у A. dendroides и A. fedtschenkoanus палисадная паренхима чрезвычайно толстая (166,05±1,4 и 185,7±1,8 мкм соответственно). Показатели толщины губчатой паренхимы (ГП) у изученных видов имеют такую же закономерность, что верхняя палисадная паренхима (Рисунок 2).
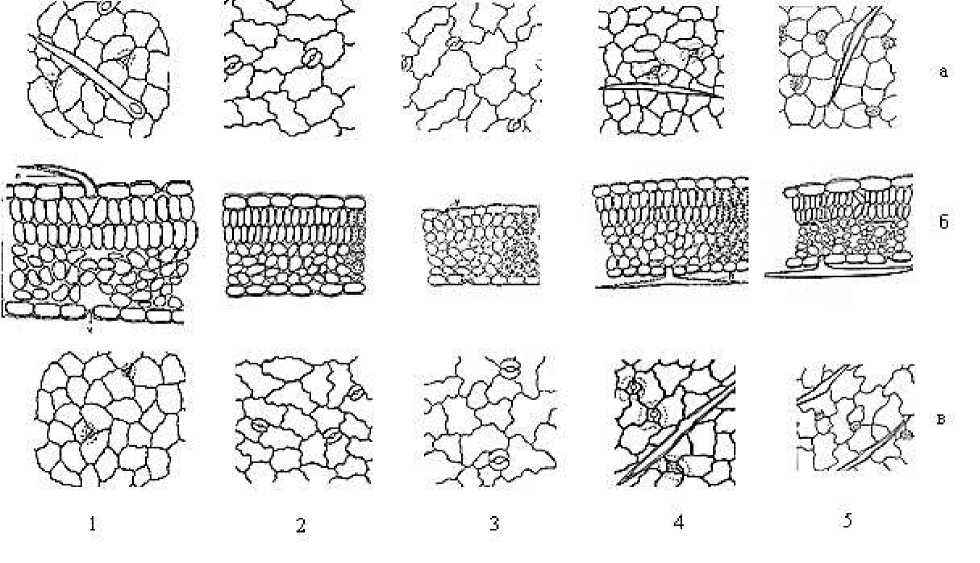
Рисунок 2. Анатомическое строение зрелых ювенильных листьев некоторых видов рода Astragalus L. 1 — A. filicaulis, 2 — A. campylorrhynchus, 3 — A. rhacodes, 4 — A. fedtschenkoanus, 5 — A. dendroides. а — верхняя эпидерма, б — поперечный срез листовой пластинки, в — нижняя эпидерма
Эпидермальные клетки ювенильных листьев у всех изученных видов более или менее мелкие, чем таковые у семядолей, с извилистыми тангенциальными стенками. Они у A. campylorrhynchus и A. rhacodes довольно крупные, чем у остальных видов (Таблица). По размерам эпидермальные клетки верхней и нижней эпидермы почти не отличаются. Устьичные аппараты встречаются на обеих сторонах листовой пластинки, амфистоматные. Они аномоцитного и анизоцитного типов, очень многочисленные на единицу площади листовой поверхности. По числу устьиц на единицу площади верхней эпидермы виды подразделяются на три группы: чрезвычайно многочисленные (154,4±0,2 и 167,4±1,2 на 1 мм2) у кустарничка и кустарника: A. dendroides и A. fedtschenkoanus ; довольно многочисленные (135±0,7 и 153±0,82 на 1 мм2) у однолетников ( A. filicaulis и A. campylorrhynchus ); более или менее малочисленные (90,67±0,27 на 1 мм2) у многолетнего A. rhacodes.
Ювенильные листья у изученных видов отличаются от семядольных листьев с опушением из числа простых волосков за исключением A. campylorrhynchus, у которого опушение отсутствует. Опушение листьев у кустарничка A. dendroides и кустарника A. fedtschenkoanus — редкое, у многолетнего A. rhacodes — густое, а у однолетника A. filicaulis — очень густое. Число волосков на единицу площади верхней эпидермы у A. filicaulis почти в два раза больше, чем таковое у нижней эпидермы. Что касается остальных видов, где имеет место опушение листовой пластинки, то у них, наоборот, их больше на нижней эпидерме.
Листовая пластинка ювенильных листьев имеет дорсовентральное строение. Они у однолетних видов (A. filicaulis и A. campylorrhynchus) более или менее толще, чем пластинки семядолей. Что касается остальных видов, то у них, наоборот, они почти в два и более раза тоньше, чем у семядольных листьев. По толщине верхняя и нижняя эпидерма ювенильных листьев почти не отличается. Однако по этому показателю виды довольно четко отличаются друг от друга и подразделяются на три группы: 1 — толстые у однолетников (A. filicaulis и A. campylorrhynchus); 2 — средней толщины у кустарничка A. dendroides и кустарника A. fedtschenkoanus; 3 — очень тонкие у многолетнего A. rhacodes. Мезофилл состоит из верхних палисадных и нижних губчатых паренхим. Палисадная паренхима однослойная. Она у A. filicaulis и A. fedtschenkoanus очень толстая (67±0,27 и 66,3±1,2 мкм), а у многолетнего A. rhacodes — тонкая (30,06±0,68 мкм), у других — средней толщины. Губчатая паренхима состоит из 4–6 слоев округлых, продолговатых клеток. Она у A. filicaulis самая толстая (104±0,43 мкм), а у A. rhacodes — самая тонкая (53,6±0,23 мкм), у других видов — промежуточная. Таким образом, л истовые пластинки ювенильных листьев у однолетних видов (A. filicaulis и A. campylorrhynchus) очень толстые (234±0,33 и 198,7±0,21 мкм), а у многолетнего A. rhacodes — чрезвычайно тонкие (114,03±0,16 мкм). Что касается остальных видов, то они по этому показателю занимают промежуточное положение между этими видами. Эпидермальные клетки дефинитивных листьев у всех изученных видов более или менее мелкие, чем таковые у семядолей и ювенильных листьев, с более или менее прямыми или в той или иной степени извилистыми радиальными стенками. Они у А. campylorrhynchus и А. rhacodes довольно крупные, чем у остальных видов, как таковые ювенильные листья (Рисунок 3).
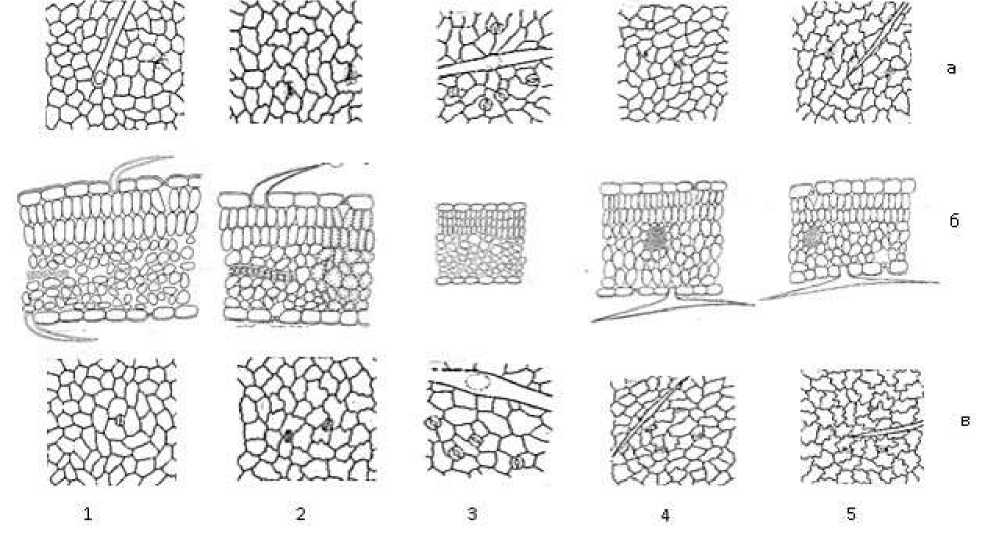
Рисунок 3. Анатомическое строение дефинитивных листьев некоторых видов рода Astragalus L., представляющие разные жизненные формы: 1 — A. filicaulis , 2 — A. campylorrhynchus , 3 — A. rhacodes , 4 — A. dendroides , 5 — A. fedtschenkoanus а — верхняя эпидерма, б — поперечный срез листовой пластинки, в — нижняя эпидерма
У большинства видов эпидермальные клетки верхней эпидермы крупнее чем нижних кроме А. dendroides , где нижние эпидермальные клетки крупнее. По данному показателю виды подразделяются на следующие группы: эпидермальные клетки средних размеров — A. campylorrhynchus , A. rhacodes, A. filicaulis , A. fedtschenkoanus (от 1111,6±7,15 до 1683,7±2,3 на 1 мм2) и эпидерма очень мелкоклеточная — A. dendroides (от 1909±3,0 до 2137±7,3 на 1 мм2).
Листовая пластинка дефинитивных листьев у изученных видов амфистоматные, т. е. устьичные аппараты встречаются на ее обеих сторонах. У А. campylorrhynchus и A. fedtschenkoanus число устьичных аппаратов на ВЭ в той или иной степени больше, чем таковые на НЭ. У остальных видов, наоборот, устьичные аппараты более часто встречаются на НЭ, чем на ВЭ.
Изученные виды более или менее четко отличаются друг от друга по числу устьичных аппаратов на единицу площади на ВЭ и НЭ и подразделяются на следующие группы: устьичные аппараты мало — A. filicaulis (95±0,5 на 1 мм2 на ВЭ), устьичные аппараты имеют средние числовые показатели — A. campylorrhynchus (181±0,96 на 1 мм2 на НЭ), A. dendroides (193,2±0,3 на 1 мм2 на ВЭ) и устьичные аппараты многочисленные — A. filicaulis (200±1,06 на 1 мм2 на НЭ), A. campylorrhynchus (219±1,2 на 1 мм2 на ВЭ), A. fedtschenkoanus (239,5±1,6 на 1 мм2 на ВЭ и 227,9±1,4 — на НЭ), А. dendroides (248,8±0,2 на 1 мм2 на НЭ), A. rhacodes (248,8±0,7 на 1 мм2 на ВЭ и 304,6±0,58 — на НЭ).
У всех изученных видов листовая пластинка дефинитивных листьев с обеих сторон опушена простыми одно-, двулучевыми волосками. Виды отличаются друг от друга числовыми показателями волосков на единицу площади поверхности листовой пластинки. Опушение листьев у A. filicaulis (109±0,32 на 1 мм2 на ВЭ) — очень густое; у A. filicaulis (39±0,05 на 1 мм2 на НЭ), A. rhacodes (37,2±0,16 на 1 мм2 на НЭ), A. campylorrhynchus (36±0,7 на 1 мм2 на ВЭ и 30±1,3 на 1 мм2 на НЭ) — умеренное, у A. rhacodes (23,5±0,22 на 1 мм2) — редкое, у A. fedtschenkoanus (3,27±0,8 на 1 мм2 на ВЭ и 8,93±0,2 на 1 мм2 на НЭ), А. dendroides (0,97±0,2 на 1 мм2 на ВЭ и 3,1±0,01 на 1 мм2 на НЭ) — единичные.
Листовая пластинка дефинитивных листьев у всех изученных видов рода Astragalus имеет дорсовентрального строения. По числу слоев мезофилла многослойные (7–10). У всех видов высота клеток ВЭ существенно не отличаются от таковых у НЭ (Рисунок 3.). По показателям толщины ВЭ и НЭ виды отличаются драг от друга: у A. fedtschenkoanus (38,7±0,8 мкм на ВЭ и 28,3±0,3 — на НЭ), A. filicaulis (31,1±0,05 мкм на ВЭ и 31,3±0,13 — на НЭ), A. dendroides (30,3±2,1 мкм на ВЭ и 29,9±1,04 – на НЭ) — толстые, у A. campylorrhynchus (22,3±0,1 мкм на ВЭ и 21±0,16 — на НЭ) — средние, у A. rhacodes (14,25±0,11 мкм на ВЭ и 16,56±0,03 мкм — на НЭ) — тонкие. У всех изученных видов рода Astragalus верхняя палисадная паренхима в той или иной степени развитая и состоит из 2–3 слоя столбчатых клеток, расположенных перпендикулярно к ВЭ. У A. filicaulis , A. campylorrhynchus , A. dendroides , A. fedtschenkoanus — двухслойные, а у A. rhacodes — трехслойные. Она у A. filicaulis (107±0,43 мкм), A. campylorrhynchus (92±0,38 мкм), A. dendroides (79,68±2,0 мкм) — толстые (высокие), у A. fedtschenkoanus (38,6±1,6 мкм) и A. rhacodes (37,4±0,07 мкм) — тонкие (невысокие). Губчатая паренхима у A. filicaulis , A. campylorrhynchus , A. rhacodes состоит из более 5 слоев, а у A. dendroides , A. fedtschenkoanus — менее 5 слоев. Таким образом, листовые пластинки дефинитивных листьев у однолетних видов ( A. filicaulis и A. campylorrhynchus ) очень толстые (288,5±0,3 и 278,3±0,31 мкм), а у многолетнего A. rhacodes — чрезвычайно тонкие (120,05±0,09 мкм). Что касается остальных видов, то они по этому показателю занимают промежуточное положение между этими видами, как таковые у ювенильных листьев (Рисунок 3).
Анализ полученных данных нашего исследования показал, что эпидермальные клетки семядолей у однолетнего A. campylorrhynchus очень крупные, а у другого однолетника A. filicaulis они относительно мелкие, другие виды по этому показателю занимают промежуточное положение между этими видами. Семядольные листья амфистоматные. Число устьиц на единицу площади на верхней эпидерме относительно больше, чем таковые на нижней эпидерме, за исключением кустарника A. fedtschenkoanus, у которого, наоборот, их число на нижней эпидерме почти 2,5 раза больше, чем таковые на верхней эпидерме. Однолетники A. filicaulis и A. campylorrhynchus имеют относительно тонкую пластинку, многолетник A. rhacodes характеризуется довольно тонкой пластинкой, а у кустарничка и кустарника (A. dendroides и A. fedtschenkoanus) обладают очень толстыми семядольными листьями. Мезофилл пластинки семядолей дифференцирован на палисадную и губчатую паренхиму и имеет дорсовентральную структуру, за исключением A. rhacodes, у которого он имеет только губчатую структуру. Виды друг от друга отличаются только по толщине семядолей.
Эпидермальные клетки ювенильных листьев у всех изученных видов более или менее мелкие с извилистыми тангенциальными стенками. По числу устьиц на единицу площади верхней эпидермы виды подразделяются на три группы: чрезвычайно многочисленные у кустарничка и кустарника — A. dendroides и A. fedtschenkoanus ; довольно многочисленные у однолетников ( A. filicaulis и A. campylorrhynchus ); более или менее малочисленные у многолетнего A. rhacodes. Ювенильные листья отличаются от семядольных листьев с опушением из числа простых волосков за исключением A. campylorrhynchus, у которого опушение отсутствует. Их пластинка имеет дорсовентральное строение. Они у однолетних видов ( A. filicaulis и A. campylorrhynchus ) более или менее толще, чем пластинки семядолей. Что касается остальных видов, то у них, наоборот, почти в два и более раза тоньше, чем у семядольных листьев. Мезофилл состоит из верхних палисадных и нижних губчатых паренхим. Палисадная паренхима однослойная. Она у A. filicaulis и A. fedtschenkoanus очень толстая, а у многолетнего A. rhacodes — тонкая, у других — средней толщины. Губчатая паренхима у A. filicaulis самая толстая, а у A. rhacodes — самая тонкая, у других видов — промежуточная. Таким образом, листовые пластинки ювенильных листьев у однолетних видов ( A. filicaulis и A. campylorrhynchus ) очень толстые, а у многолетнего A. rhacodes — чрезвычайно тонкие, другие виды по этому показателю занимают промежуточное положение между этими видами.
Эпидермальные клетки дефинитивных листьев у всех изученных видов более или менее мелкие, чем таковые у семядолей и ювенильных листьев, с более или менее прямыми или в той или иной степени извилистыми радиальными стенками. Они у A. campylorrhynchus и A. rhacodes довольно крупные, чем у остальных видов, как таковые ювенильные листья. У большинства видов эпидермальные клетки верхней эпидермы крупнее чем нижних, кроме A. dendroides , где нижние эпидермальные клетки крупнее. Листовая пластинка дефинитивных листьев у изученных видов амфистоматные. У A. campylorrhynchus и A. fedtschenkoanus число устьичных аппаратов на ВЭ в той или иной степени больше, чем таковые на НЭ. У остальных видов, наоборот, устьичные аппараты более часто встречаются на НЭ, чем на ВЭ. У всех изученных видов листовая пластинка дефинитивных листьев с обеих сторон опушена простыми одно-, двулучевыми волосками. Виды отличаются друг от друга числовыми показателями волосков на единицу площади поверхности листовой пластинки. Опушение листьев у A. filicaulis , A. rhacodes , A. campylorrhynchus — густое, у A. fedtschenkoanus и A. dendroides — единичные. Листовая пластинка дефинитивных листьев у всех изученных видов рода Astragalus имеет дорсовентрального строения. По числу слоев мезофилла многослойные (7–10). У всех видов высота клеток ВЭ существенно не отличаются от таковых у НЭ. У всех изученных видов рода Astragalus верхняя палисадная паренхима в той или иной степени развитая и состоит из 2–3 слоя столбчатых клеток, расположенных перпендикулярно к ВЭ. У A. filicaulis , A. campylorrhynchus , A. dendroides , A. fedtschenkoanus
— двухслойные, а у A. rhacodes — трехслойные. Она у A. filicaulis , A. campylorrhynchus , A. dendroides — толстые (высокие), у A. fedtschenkoanus и A. rhacodes — тонкие (невысокие). Губчатая паренхима у A. filicaulis , A. campylorrhynchus , A. rhacodes состоит из более 5 слоев, а у A. dendroides , A. fedtschenkoanus — менее 5 слоев. Таким образом, листовые пластинки дефинитивных листьев у однолетних видов ( A. filicaulis и A. campylorrhynchus ) очень толстые, а у многолетнего A. rhacodes — чрезвычайно тонкие, остальные виды занимают промежуточное положение между этими видами, как таковые у ювенильных листьев.
Выводы
-
1. В лабораторных условиях всходы у изученных видов появляются через 20–35 дней после посева. Прорастание семян у всех видов гипокотилярное. Семядольные листья проростков растут очень медленно в течение от 7–9 дней ( A. campylorrhynchus ) до 25–30 дней ( A. dendroides и A. fedtschenkoanus ), т. е. их размеры очень мало изменяются с момента распускание. Можно отметить наличие некоторой корреляции между формой и длиной листовой пластинки семядольных листьев: так, у однолетних они удлиненно-яйцевидные, довольно короткие, а у многолетних трав и кустарников — неравнобокие, яйцевидные, обратнояйцевидные, заметно длинные. Первые настоящие листья у всех изученных видов начинают формироваться через 10–18 дней после появления проростков.
-
2. В процессе индивидуального развития растений наблюдается морфологическая гидрофилия, который выражается в виде смены простых листьев на тройчатые и, далее вверх по главному побегу на более сложные непарноперистые листья. У A. rhacodes листья главного побега до 5–6 (8) узла простые, далее вверх появляются непарноперистосложные листья 3–5–7 листочками. У A. filicaulis и A. campylorrhynchus листья главного побега начиная со второго узла сложные, тройчатые, у первого с третьего узла формируются непарноперистосложные листья с 6–7 парами листочков, а у второго тройчатые листья располагаются до 5–6 узла включительно. Далее вверх располагаются непарноперистосложные листья с 6–7 парами листочков. У третьей группы все настоящие листья с первого узла сложные. Они у A. fedtschenkoanus со второго узла непарноперистосложные с 5–7 (9) листочками, а у A. dendroides тройчатосложные листья с 1 по 6 узла, далее вверх по главному побегу располагаются непарноперистосложные листья с 5 (7) листочками.
-
3. Семядольные листья голые, эпидермальные клетки у всех изученных видов изодиаметрические, удлиненные и удлиненно-изодиаметрические, амфистоматные. Устьичные аппараты аномоцитного и анизоцитного типов, которые расположены на одинаковом уровне с собственными клетками эпидермы.
-
4. Ювенильные листья отличаются от семядольных листьев опушением из числа простых волосков за исключением A. campylorrhynchus, у которого опушение отсутствует.
-
5. В отличие от семядольных и ювенильных листьев дефинитивные листья изученных видов характеризуются мелкоклеточностью эпидермальных клеток, большой толщиной палисадной, губчатой паренхимы, и в целом листовой пластинки.
-
6. Изученные виды отличаются друг от друга формой ювенильных листьев, характером смены форм листовой пластинки от простого или тройчатосложного до непарноперистосложного в процессе формирования главного побега в онтогенезе видов, размерами эпидермальных клеток, числом устьичных аппаратов на единицу площади, высотой верхних и нижних эпидермальных клеток, числом слоев клеток мезофилла и общей толщиной листовой пластинки.
Список литературы Анатомо-морфологические особенности листовых органов некоторых видов рода Astragalus L. в их онтогенезе
- Бутник А. А., Нигманова Р. Н., Пайзиева С. А., Саидов Д. К. Экологическая анатомия пустынных растений Средней Азии. Т. 1: Деревья, кустарники, кустарнички. Т. 1. Ташкент: Фан, 1991. 145 с.
- Василевская В. К. Изучение онтогенеза как один из методов экологической анатомии // Проблемы ботаники. 1950. Т. 1. С. 264-281.
- Васильченко И. Т. О филогенетическом значении смены листовых форм в онтогенезе астрагалов // Ботанический журнал. 1961. Т. 46. №12. С. 1735-1739.
- Маматкулов О. И. Морфо-биологические особенности индивидуального развития Astragalus fedtschenkoanus Lipsky // Вестник Ошского государственного университета. 2006. №3. С. 215-217.
- Маматкулов О. И. Морфобиологические особенности индивидуального развития Astragalus campylorrhynchus Fisch. et Mey (Fabaceae L.) // Проблемы сохранения и восстановления особо охраняемых природных территорий Центральной Азии: Материалы конференции. Жалалабад, 2006. С. 104-106.
- Маматкулов О. И., Коланов О. Онтогенетические особенности Astragalus dendroides Kar. et Kir. (Fabaceae L.) // Ботаника, экология, охрана растений: Материалы международной научной конференции. Андижан, 2007. С. 80-82.
- Маматкулов О. И. Морфо-биологические особенности индивидуального развития Astragalus filicaulis Fisch. et Mey. (Fabaceae L.) // Вестник Ошского государственного университета. 2009. №3. С. 60-63.
- Маматкулов О. И. Онтогенетические и морфобиологические особенности Astragalus rhacodes Bunge (Fabaceae L.) // Известия вузов. 2014.. №7. С. 25-27.
- Лакин Г. Ф. Биометрия. М.: Высшая школа, 1990. 352 с.
- Пахомова М. Г. К методике просветления листьев хлопчатника для анатомических исследований // Доклады АН УзССР. 1963. №11. С. 45-50.
- Захаревич С. Ф. К методике описания эпидермиса листа // Вестник Ленинградского университета. 1954. Т. 4. С. 65-75.
