Арахидонил-липиды мононуклеарных клеток крови как биомаркеры для раннего обнаружения разных опухолей

Автор: Акопян Г.В., Торгомян Т.Р., Лазян М.П., Арутюнян К.С., Батикян Т.Б., Алексанян К.А., Тадевосян Ю.В.
Журнал: Juvenis scientia @jscientia
Статья в выпуске: 11, 2017 года.
Бесплатный доступ
В статье исследованы закономерности кратковременного (5 сек) включения и относительно длительного (60 мин) межфракционного перераспределения экзогенной [14C]арахидоновой кислоты (АК) в различные фракции фосфолипидов (ФЛ) и нейтральных липидов (НЛ) мононуклеарных клеток (МНК), выделенных из периферической крови практически здоровых доноров, больных раком молочной железы (РМЖ) и яичников (РЯ). Полученные данные свидетельствуют о достоверных и идентичных нарушениях в механизмах как быстрого, так и пролонгированного АК-модификаций липидов МНК периферической крови при РМЖ и РЯ. Делается заключение о вовлечении процессов АК-модификации жирнокислотного состава липидов клеточных мембран МНК крови в патогенез РМЖ и РЯ.
Мононуклеарные клетки, липиды, арахидоновая кислота, рак молочной железы, рак яичников
Короткий адрес: https://sciup.org/14110433
IDR: 14110433 | УДК: 577 | DOI: 10.15643/jscientia.2017.11.007
Arachidonil-lipids of blood mononuclear cells as a biomarker for early detection of different tumors
The regularities of quick (5 sec) incorporation of exogenous [14C]arachidonic acid (AA) into the various fractions of phospholipids (PL) and neutral lipids (NL) and it’s long-term (60 min) interfractional conversions in crude mononuclear cells (MNC) isolated from the peripheral blood of patients with breast (BC) and ovarian (OC) cancers in comparison with the healthy people was examined in the article. The data obtained provide evidence for reliable and identical disturbances in the MNC membrane lipids quick as well as long-term AA-modification mechanisms in BC and OC. We concluded that in the peripheral blood MNC some of the mechanisms responsible for fatty acid-content modification of lipids by AA are involved in the pathogenesis of BC and OC.
Текст научной статьи Арахидонил-липиды мононуклеарных клеток крови как биомаркеры для раннего обнаружения разных опухолей
Введение . Одним из наиболее важных факторов, влияющих на исход рака, является время обнаружения рака, как при первоначальном диагнозе, так и при рецидиве опухоли [1]. В настоящее время большинство случаев рака выявляются относительно поздно, что приводит к высокой смертности.
Основываясь на обнаружении многочисленных биомаркеров рака, предполагалось, что развитие биомаркеров периферической крови хорошо предназначено для улучшения диагностики, управления и лечения злокачественных опухолей [2-4]. Эта так называемая стратегия «основенная на исследовании крови», применима для обнаружения и мониторинга рака с использованием различные клеточные и молекулярные элементы периферической крови (белки, мРНК, циркулирующие опухолевые клетки, метаболиты и др.). Это очень привлекательная стратегия и может способствовать доступным скрининговым программам рака, но часто вытерпивает из-за проблем, возникающих в результате чувствительности и специфичности низких концентра- ций биомаркеров в крови [5], быстрой деградации маркерных молекул in vivo и ex vivo [6], гетерогенности опухолей и сильно варьируемой фоновой экспрессии биомаркеров в незлокачественных тканях [7]. Следовательно, современные диагностические тесты для обнаружение рака довольно недостаточны [8-9], поэтому существует настоятельная необходимость в новых, эффективных неинвазивных молекулярных тестах, основанных на исследования крови, и действующих на ранней стадии заболевания, когда лечебные вмешательства все еще возможны.
В современной науке общепринято, что наиболее жизненно важные функции клеток происходят внутри или вокруг клеточной плазматической мембраны (ПМ), состоящей из белков и липидов. В этом контексте, при воздействии на клетку любого внешнего (физиологического или патогенного) сигнала, в зависимости от его длительности и интенсивности, происходит обратимое либо необратимое, кратковременное или хроническое смещение исходного и установление нового, так называемого изме-
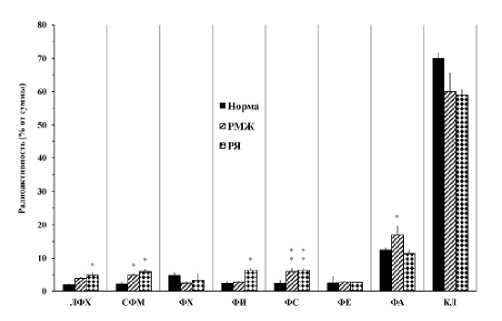
Рисунок 1. Быстрое (5 сек) включение экзогенной [14C]арахидоновой кислоты в фосфолипиды мононуклеарных клеток крови в норме, при раке молочной железы и раке яичников.
Примечание: Здесь и на следующих рисунках данные представлены в процентах от суммы радиоактивности, включенной в мембранные липиды.
На всех рисунках – * – P < 0.5; ** – P < 0.001
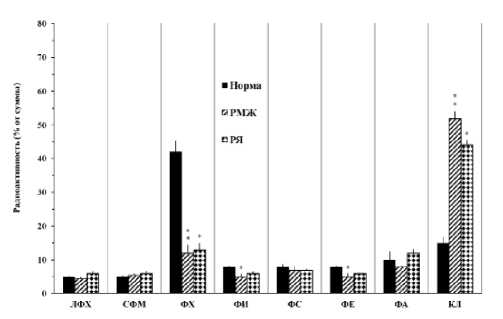
Рисунок 2. Пролонгированное (60 мин) межфракционное перераспределение экзогенной [14C]арахидоновой кислоты в фосфолипиды мононуклеарных клеток крови в норме, при раке молочной железы и раке яичников
ненного «метаболического статуса покоя» ПМ. В этих процессах липидный компонент липопротеинового бислоя ПМ участвует быстрыми и обратимыми сдвигами и активными реакциями модификации. Исходя из результатов ранее проведенных нами исследований [10-13], а также данных литературы [14-15], известно, что подобные сдвиги в модификации жирнокислотного (ЖК) состава мембранных липидов выявляются уже с первых секунд действия на клетку внешних сигналов.
Как известно, по своей функциональной значимости в жизнедеятельности клеток особое место занимают липиды, содержащие арахидоновую кислоту (АК), их метаболиты, а также свободная АК.
Согласно имеющимся данным, АК вовлекается в процессы сигнальной трансдукции [16] или стабилизации мембранных структур [17] в качестве регулятора текучести липидного бислоя. Однако, в научной литературе отсутствуют данные о возможных нарушениях в механизмах АК-модификации липидов мембран мононуклеарных клеток (МНК) периферической крови больных раком.
МНК состоят из лимфоцитов (включая Т и В-клетки, клетки естественного киллера), моноцитов, макрофагов и дендритных клеток и играют разнообразную и важную роль в мониторинге иммунного гомеостаза, в противоопухолевом ответе, а также в хеморезистентности опухлевых клеток. МНК – как легкодоступный клеточный пул крови, в последнее время используются для изучения механизмов развития рака и для открытия новых, неинвазивных биомаркеров [18-19].
В настоящей работе представлены результаты сравнительных исследований процессов кратковременного (5 сек) включения экзогенной [14C]АК в различные фракции фосфолипидов (ФЛ) и нейтральных липидов (НЛ) и ее относительно более длительного (60 мин) межфракционного перераспределения в ПМ МНК, выделенных из периферической крови практически здоровых доноров и больных раком молочной железы (РМЖ) и яичников (РЯ).
Методы исследования. В работе использованы пробы перифрической крови 10-ти практически здоровых доноров, 14-и первичных больных с РМЖ (стадии Т1N0М0 и Т2N1М0) и 9-и больных с РЯ (стадия Т3NхМ1) по классификации ТNМ. Пробы крови были предоставлены Национальным центром онкологии имени В.Фанарджяна МЗ РА. МНК из цельной крови выделяли методом Иннеса и др. [20] в градиенте фиколл-верографина (1 часть 32.8% верографина и 2.4 части 8% водного раствора фиколла 400 000, d =1.076-1.077). Интактность клеток, определяемая окрашиванием трипановым синим, составляла более 90%. При чрезмерном загрязнении МНК эритроцитами последние лизировали гипотоническим шоком. МНК осаждали при 650g в течение 15 мин и ресуспендировали (106 клеток/мл) в минимальной основной среде Игла (фирмы «Sigma», pH 7.4).
Включение экзогенной [14С]АК в липиды ПМ интактных клеток осуществляли ранее описанным методом [10]. Для этого при 37 оС к клеточной супензии добавляли 0.2 мкКю [14С]АК (сп. акт. 56 мКю/ммоль, «Amersham International», Великобритания), в среде Игла (pH 7.4). На 5 сек и 60 мин инкубации реакции останавливали приливанием холодной смеси хлороформ-метанола (1:2, об/об).
Экстракцию липидов осуществляли по методу Блай и Дайер [21]. Фракционирование ФЛ проводили одномерной ТСХ на на фирменных («Merck», Германия) ТСХ пластинках в системе растворителей хлороформ-ацетон-метанол-уксус-ная кислота-вода (6:8:2:2:1, об/об). НЛ фракционировали в смеси растворителей петролейный эфир-диэтиловый эфирмуравьиная кислота (30:10:1, об/об).
Распределение радиоактивности в идентифицированных соответствующими химически чистыми стандартами («Sigma», США) и проявленных в парах йода липидных фракциях определяли сканированием ТСХ пластинок на радиосканере фирмы «Berthold» (Германия). Степень радиоактивности проб определяли в жидкости Брея на сцинтилляционном спектрометре «Roche-Bioelectronique Kontron», модель SL-4221 (Франция). Статистический анализ проводили согласно методу Стьюдента.
Результаты и обсуждение . Результаты исследования процессов быстрого (5 сек) включения АК в мембранные ФЛ МНК практически здоровых доноров (далее будет отмечен как норма) обнаружили (Рис. 1) преимущественного накопления радиоактивности (более 60% от суммы включенной радиоактивности) во фракции кардиолипинов (КЛ). Во фракции фосфатидных кислот (ФК) относительное содержание радиоактивности составило около 13%, а в остальных фракциях колебалось в пределах 2-5%. Обнаруженный нами факт специфичности КЛ, как предпочитаемого акцептора для быстрых процессов эстерификации АК, дополняют литературные данные о ключевой функциональной значимости в жизнедеятельности клеток ЖК состав КЛ фракции [22-23].
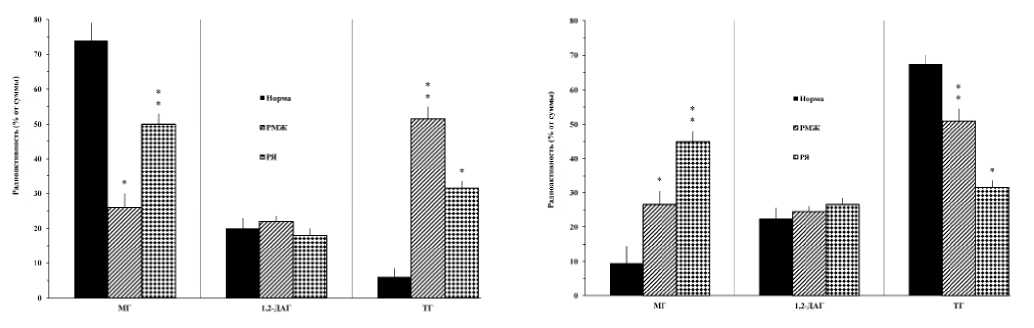
Рисунок 3. Быстрое (5 сек) включение экзогенной [14C]арахидоновой кислоты в нейтральные липиды мононуклеарных клеток крови в норме, при раке молочной железы и раке яичников
Рисунок 4. Пролонгированное (60 мин) межфракционное перераспределение экзогенной [14C]арахидоновой кислоты в нейтральные липиды мононуклеарных клеток крови и в норме, при раке молочной железы и раке яичников
В МНК больных с РМЖ и РЯ в идентичных условиях эксперимента были выявлены (рис. 1), в основном, схожие нарушения в процессах быстрого включения экзогенной АК в исследованные ФЛ фракции. В частности, при двух формах патологии наблюдалось уменьшение относительного содержания КЛ и некоторое повышение всех остальных АК-содержащих ФЛ фракций (лизофосфатидилхолины – ЛФХ, сфингомиелины – СФМ, фосфатидилсерины – ФС и фосфа-тидилинозиты – ФИ и фосфатидные кислоты – ФК). В случае фосфатидилхолинов (ФХ), уровень включенной АК понижена от нормы при РМЖ и РЯ, а во фракции фосфатидилэтано-ламинов (ФЭ) – равномерна норме.
На основании представленных экспериментальных данных, можно заключить, что при РМЖ и РЯ выявленные нарушения АК ацилирования ФЛ на быстрой (5 сек) мембраносвязанной фазе процессов липидных модификации являются характерным признаком для патологических МНК.
Как известно, в быстропротекающих процессах изменения липопротеинового бислоя ПМ участвует, в основном, внутримембранный потенциал модификационных механизмов. В частности, в процессах реконструирования ЖК состава мембранных липидов вовлечен так называемый «цикл Ландса» [24] и включающий ферменты деацилирования (фосфолипазы типа А) и реацилирования (ацилтранс-феразы, трансацилазы) [25-26].
При относительно длительном воздействии внешних факторов, в модификационные процессы липидов вовлекаются также внутриклеточныe кооперативныe механизмы ПМ. Изучение процессов включения и перераспределения АК в мембранные ФЛ в течение 60 мин в норме показало (рис. 2) преимущественное накопление метки (около 45%) в количественно преобладающей во внешнем монослое мембран МНК фракции ФХ. Во фракции КЛ накопление АК составляло около 20%, а в остальных ФЛ фракциях варьировало в пределах 5-10%.
Выявленные закономерности в относительно длительных процессах реацилирования/деацилирования мембранных ФЛ нарушены при РМЖ и РЯ. Так, избирательное накопление ЖК обнаруживаются во фракциях КЛ (до 4050%), а не ФХ (около 15%). В остальных исследованных ФЛ фракциях уровни АК идентифицируются в пределах нормы. На наш взгляд, в процессах как быстрого (5 сек) ацилирования, так и более пролонгированных (60 мин) перераспределения ЖК в мембранных ФЛ выявляются идентичные дефекты в ПМ патологических МНК. Превалирующий субстрат для накопления АК является КЛ при РМЖ и РЯ, что может быть вследствии аналогично активированных ЖК-модифицирующих ферментов – ацилтрансферазов, транс-ацилазов или фосфолипазов.
В следующей серии экспериментов были проведены сравнительные исследования по изучению закономерностей включения АК во фракции моноацилглицеринов (МГ), 1,2-диацилглицеринов (1,2-ДАГ) и триацилглицеринов (ТГ) в мембранах нормальных и патологических МНК.
При 5 сек инкубации МНК с экзогенной АК в норме выявило (рис. 3) включение ЖК во фракции МГ, 1,2-ДАГ и ТГ в соотношении 75, 20 и 5%, соответственно. В экспериментальных сериях с патологическими МНК обнаруживалось достоверное понижение АК во фракции МГ (при РМЖ 25%, при РЯ 45%) и повышение во фракции ТГ (при РМЖ 50%, при РЯ 30%, соответственно) по сравнению с нормой.
При этом уровень АК во фракции 1,2-ДАГ практически остается в пределах нормы. Следовательно, по сравнению с нормой при исследованных двух формах рака обнаруживаются схожие нарушения в механизмах быстрого включения АК во фракции НЛ МНК. Эти сдвиги, вероятно, обусловлены повышением активности ферментных систем межфракционного перераспределения АК по схеме МГ → ДАГ → ТГ, приводящих к быстрому накоплению метки во фракции ТГ, которые выполняют функцию «временного депо» [27] для избытка свободной АК.
Аналогичное исследование в условиях 60 мин инкубации в норме обнаружило (рис. 4) накопление АК во фракции ТГ (около 70%), 1,2-ДАГ (почти 20%) и МГ (10%). Характерно, что при РМЖ и РЯ по сравнению с нормой достоверно подавлялось пролонгированная эстирификация АК во фракции ТГ (50 и 30%, соответственно) на фоне многократного повышения уровня АК в МГ (25 и 45%, соответственно). В этом случае, как и при быстрых процессах ацилирования НЛ, уровни ДАГ оставались в пределах нормы. Эти результаты указывают на торможение при исследованных неоплазиях МГ → ДАГ → ТГ путей пролонгированных фракционных взаимопревращений, активированных при быстрых (5 сек) процессах ацилирования НЛ.
Обобщая вышеприведенные экспериментальные данные, можно заключить, что по сравнению с нормой в липидном бислое ПМ МНК периферической крови больных РМЖ и РЯ преобладают патологические изменения метаболического статуса покоя. В частности, МНК больных характеризуются наличием «дефектных» звеньев в механизмах АК-модификации отдельных фракций как ФЛ, так и НЛ. Некоторые значительно измененные фракции ФЛ (ФХ и КЛ)
и НЛ (МГ, ТГ), в PM МНК могут быть использованы в качестве перспективных биомаркеров, для раннего выявления и оценки злокачественности. Следует отметить, что необходимы дальнейшие исследования для выяснения корреляции между уровнями дефектных липидов и ферментами липидного метаболизма в патогенезе РМЖ и РЯ.
Список литературы Арахидонил-липиды мононуклеарных клеток крови как биомаркеры для раннего обнаружения разных опухолей
- Etzioni R., Urban N., Ramsey S., et al. The case for early detection//Nature Reviews Cancer. 2003. Vol. 3. No. 4. Pp. 243-252.
- Hanash S.M., Balk C.S., Kallioniemi O. Emerging molecular biomarkers-blood-based strategies to detect and monitor cancer//Nature Reviews Clinical Oncology. 2011. Vol. 8. Pp. 142-150.
- Bell D.W., Haber D.A. A blood-based test for epidermal growth factor receptor mutation in lung cancer//Clinical Cancer Research. 2006. Vol. 12. Pp. 3875-3877.
- Ronald J.A., Chuang Hui-Yen, Dragulescu-Andrasi A., et al. Detecting cancers through tumor-activatable minicircles that lead to a detectable blood biomarker//Proceedings of the National Academy of Sciences of the United States of America. 2015. Vol. 112. No. 10. Pp. 3068-3073.
- Nagrath S., Sequist L.V., Maheswaran S., et al. Isolation of rare circulating tumour cells in cancer patients by microchip technology//Nature. 2007. Vol. 450. No. 7173. Pp. 1235-1239.
- Haun J.B., Castro C.M., Wang R., et al. Micro-NMR for rapid molecular analysis of human tumor samples//Science Translational Medicine. 2011. Vol. 3. No. 71 Pp. 1-27.
- Diamandis E.P. Cancer biomarkers: Can we turn recent failures into success?//Journal of the National Cancer Institute. 2010. Vol. 102. No. 19. Pp. 1462-1467.
- Hori S.S., Gambhir S.S. Mathematical model identifies blood biomarker-based early cancer detection strategies and limitations//Science Translational Medicine. 2011. Vol. 3. No. 109. Pp. 1-19.
- Kern S.E. Why your new cancer biomarker may never work: Recurrent patterns and remarkable diversity in biomarker failures//Cancer Research. 2012. Vol. 72. No. 23. Pp. 6097-6101.
- Мелконян М.Г., Акопян Г.В., Тадевосян А.Ю. и др. Ацилирование липидов арахидиновой кислотой и образованое липидных вторичных посредников в анти-CD3/CD28 костимулированных лимфоцитах в норме и при различных формах лейкемии//Медицинская наука Армении. 2007. Т. XLVII. № 2. С. 56-63.
- Тадевосян А.Ю., Батикян Т.Б., Асатрян Л.Ю. и др. Закономерности включения арахидоновой кислоты в липиды мембран нормальных и лейкемических лимфоцитов//Медицинская наука Армении. 1999. Т. 39, № 3. С. 33-40.
- Тадевосян Ю.В., Асатрян Л.Ю., Карагезян К.Г. Процессы деацилирования мембранных фосфолипидов при инициации фосфоинозитидного цикла в синаптосомах крыс и лимфоцитах человека//Нейрохимия. 1992. Т. 11. № 2. С. 194-205.
- Тадевосян Ю.В., Асатрян Л.Ю., Батикян Т.Б. и др. Катаболизм мембранных фосфатидилхолинов в лимфоцитах на ранних стадиях митогенной стимуляции фосфоинозитидного цикла//Биохимия. 1996. Т. 61, вып. 8. С. 1414-1421.
- Suzuki K.G., Fujiwara T.K., Sanematsu F., et al. GPI-anchored receptor clusters transiently recruits Lyn and Gα for temporary cluster immobilization and Lyn activation: single-molecule tracking study//Journal of Cell Biology. 2007. Vol. 177. No. 4. Pp. 717-730.
- Patterson A.D., Maurhofer O., Beyoglu D., et al. Aberrant lipid metabolism in hepatocellular carcinoma revealed by plasma metabolomics and lipid profiling//Cancer Research. 2011. Vol. 71. No. 21. Pp. 6590-6600.
- Gong Y., Zhang Z. Alternative signaling pathways: when, where and why?//FEBS Letters. 2005. Vol. 579, No. 24. Pp. 5265-5274.
- Leekumjorn S., Cho H.J., Wu Y., et al. The role of fatty acid unsaturation in minimizing biophysical changes on the structure and local effects of bilayer membranes//Biochimica et Biophysica Acta. 2009. Vol. 1788. Pp. 1508-1516.
- Koncarevic S., Lößner C., Kuhn K., et al. In-depth profiling of the peripheral blood mononuclear cells proteome for clinical blood proteomics//International Journal of Proteomics. 2014. Vol. 2014. Pp. 1-9
- Silva C., Santa C., Anjo S.I., et al. A reference library of peripheral blood mononuclear cells for SWATH-MS analysis//Proteomics -Clinical Applications. 2016. Vol. 10. No. 7. Pp. 760-764
- Innes J., Runtz M.M., Kim Y.T. et al. Induction of suppressor activity in the autologous mixed lymphocyte reaction and in cultures with ConA//Journal of Clinical Investigation. 1979. Vol. 64. Pp. 1608-1613.
- Bligh E.G., Dyer W.I. A rapid method for lipid extraction and purification//Canadian Journal of Biochemistry and Physiology. 1959. Vol. 37. Pp. 911-917.
- Li X.X., Tsoi B., Li Y.F., Kurihara H., et al. Cardiolipin and its different properties in mitophagy and apoptosis//Journal of Histochemistry & Cytochemistry. 2015. Vol. 63. No. 5. Pp. 301-311.
- Xu Y., Kelley R.I., Blanck T.J., et al. Remodeling of cardiolipin by phospholipid transacylation//Journal of Biological Chemistry. 2003. Vol. 278. No. 51. Pp. 51380-51385.
- Lands W.E.M. Stories about acyl chains//Biochimica et Biophysica Acta. 2000. Vol. 1483. Pp. 1-14.
- Perez R. Matabosch X., Llebaria A., et al. Blockade of arachidonic acid incorporation into phospholipids induces apoptosis in U937 promonocytic cells//Journal of Lipid Research. 2006. Vol. 47. Pp. 484-491.
- Yamashita A., Sugiura T., Waku K. Acyltransferases and transacylases involved in fatty acid remodeling of phospholipids and metabolism of bioactive lipids in mammalian cells//Journal of Biochemistry. 1997. Vol. 122. Pp. 1-16.
- Igal R.A., Whang S., Gonzalez-Baro M., et al. Mitochondrial glycerol phosphate acyltransferase directs the incorporation of exogenous fatty acids into triacylglycerol//Journal of Biological Chemistry. 2001. Vol. 276. Pp. 42205-42212.
