Аргинин и оксид азота (NO) в эмбриогенезе сельскохозяйственной птицы (обзор)
 в эмбриогенезе сельскохозяйственной птицы (обзор)")
Автор: Титов В.Ю., Долгорукова А.М., Кочиш И.И., Мясникова О.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Эмбриональный период у бройлеров по продолжительности занимает практически треть от времени существования организма птицы, поэтому особенности развития на этом этапе очень значимы. Важным фактором, обусловливающим развитие, служат незаменимые аминокислоты, к которым относится аргинин. Аргинин — также субстрат синтеза оксида азота (NO), который служит медиатором тонуса гладкой мускулатуры, дифференциации тканей и апоптоза, экспрессии ряда генов. Возможность влиять на эти эффекты NO открывает широкие перспективы для регуляции различных физиологических процессов. В работах многих исследователей показано, что экзогенный ввод аргинина in ovo (даже в незначительном по сравнению с уже имеющимся в яйце количестве) оказывает эффект на эмбриональное и постэмбриональное развитие. Причем эти эффекты противоречивы, и нет убедительной гипотезы касательно их механизма (S. Subramaniyan с соавт., 2019; Y. Li с соавт., 2016). В настоящем обзоре рассмотрены пути метаболизма аргинина и его производного — NO в эмбриогенезе сельскохозяйственной птицы и связанные с ними физиологические эффекты. Показано, что аргинин в яйце содержится в количестве, превышающем необходимое для реализации физиологических процессов в эмбрионе (H. Zhu с соавт., 2022; В.Ю. Титов с соавт., 2022). В гомогенате развивающегося птичьего эмбриона, а также в плазме его крови концентрация свободного аргинина достигает нескольких сотен мкмоль/л, что многократно превышает концентрацию насыщения NO-синтазы — фермента, катализирующего синтез NO из аргиина. Тем не менее экзогенный ввод аргинина способствует увеличению синтеза NO (F. Dioguardi, 2011; S. Elms с соавт., 2013). Это явление называют «аргининовым парадоксом». Предполагают, что оно связано с наличием эндогенных конкурентных ингибиторов NO-синтазы, которые увеличивают концентрацию насыщения. Однако более конкретный анализ показывает, что ввод аргинина in ovo до 10-х сут инкубации не приводил к увеличению концентрации производных NO как в гомогенате, так и в плазме крови эмбриона. В то же время содержание аргинина в плазме и гомогенате повышалось. Введение аргинина в яйцо на заключительном этапе предплодного периода способствовало повышению концентрации производных NO, причем достоверно оно имело место в только в плазме крови, а в других тканях было незначительным. Следовательно, либо на этом этапе развития появляются конкурентные ингибиторы, либо аргинин, введенный в плазму, быстро теряет усвояемость. В пользу последнего говорит то, что при введение аргинина до закладки на инкубацию его содержание в плазме остается повышенным до заключительной стадии предплодного периода, но не происходит интенсификации синтеза NO (В.Ю. Титов с соавт., 2024). Оксид азота влияет на экспрессию ряда генов, ответственных за миогенез (D. Cazzato., с соавт., 2014). Практическое значение имеет то обстоятельство, что в мышечной ткани эмбрионов пород, линий и кроссов мясного направления продуктивности NO окисляется до нитрата, который транспортируется в аллантоис. В эмбрионах яичных форм NO депонируется в составе нитрозотиолов, нитрозильных комплексов железа и высокомолекулярных нитратов. Установлено, что окисление NO в эмбрионе — генетически детерминированный процесс, опосредованный как минимум двумя генами. Регулировать этот процесс искусственно проблематично, поскольку аргинин в мышечной ткани эмбриона находится в концентрации насыщения для NO-синтазы, а ее блокаторы действуют короткое время. Для соединений-доноров NO амниотическая оболочка непроницаема. Однако степень окисления эмбрионального NO до нитрата — очень чувствительный селекционный показатель (В.Ю. Титов с соавт., 2022). Мы предполагаем, что эффекты, наблюдаемые при экзогенном вводе аргинина in ovo в количествах, не превышающих 1,0 мкмоль/г массы яйца, обусловлены преимущественно увеличением концентрации соединений-доноров NO в плазме крови. Они могут влиять на тонус сосудов и, возможно, на тонус гладкой мускулатуры протоков пищеварительных желез.
Аргинин, оксид азота, NO, сельскохозяйственная птица, миогенез, экспрессия генов, живая масса
Короткий адрес: https://sciup.org/142247684
IDR: 142247684 | УДК: 636.5:591.392:57.044 | DOI: 10.15389/agrobiology.2026.2.209rus
Arginine and nitric oxide (NO) in the embryogenesis of poultry (review)
The embryonic period takes up almost a third of the broiler’s lifetime. Therefore, the developmental features at this stage are especially significant. Essential amino acids, in particular arginine, are an important factor contributing to body development. Arginine is also a substrate for the synthesis of nitric oxide (NO) which serves as a mediator of smooth muscle tone, tissue differentiation and apoptosis, and the expression of a number of genes. The ability to influence these NO effects opens up wide possibilities for regulating various physiological processes. Many researchers have shown that exogenous arginine injection in ovo has an effect on embryonic and postembryonic development, even upon injection of its small amount as compared to that already contained in the egg. Moreover, these effects are contradictory, and there is no convincing hypothesis regarding their mechanism (S. Subramaniyan et al., 2019; Y. Li et al., 2016). This review examines the metabolic pathways of arginine and its derivative NO during poultry embryogenesis and their associated physiological effects. It has been shown that egg arginine concentrations exceed those necessary for physiological processes in the embryo (H. Zhu et al., 2022; V. Titov et al., 2022). In the homogenate of the developing embryo, as well as in blood plasma, the concentration of free arginine reaches several hundred micromoles/l, which is many times higher than the saturation concentration for NO synthase, an enzyme that catalyzes the synthesis of NO from arginine. Nevertheless, exogenous arginine injection increases NO synthesis (F. Dioguardi, 2011; S. Elms et al., 2013). This phenomenon is called the ‘arginine paradox’. It is assumed that its cause is the presence of endogenous competitive inhibitors of NO synthase which increase the concentration of its saturation. However, a more specific analysis shows that the administration of arginine before day 10 of incubation did not lead to increase in the concentration of NO derivatives in both the homogenate and the blood plasma of the embryo. The exogenous arginine in the egg at the final stage of the prenatal period contributed to an increase in the concentration of NO derivatives. It reliably occurred only in blood plasma, and in other tissues, it was insignificant. Thereby, either the competitive inhibitors appear at this stage of development, or arginine in plasma quickly loses its digestibility. The latter is supported by the fact that when arginine is administered before incubation, its plasma content remains elevated until the final stage of the prenatal period. However, there is no intensification of NO synthesis. (V.Y. Titov et al., 2024). Nitric oxide affects the expression of a number of genes responsible for myogenesis (D. Cazzato, et al., 2014). Of practical importance is the fact that in the muscle tissue of embryos of meat type breeds, NO is oxidized to nitrate which is transported to allantois. In the embryos of egg-type breeds NO is accumulated in the nitrosothiols, nitrosyl iron complexes and high molecular weight nitrates. It has been established that NO oxidation in the embryo is genetically determined and mediated by at least two genes. It is difficult to artificially regulate this process, since arginine in the muscle tissue of the embryo is in a saturation concentration for NO-synthase, and blockers of this enzume act for a short time. The amniotic membrane is impenetrable for nitro- and nitroso compounds. However, the degree of oxidation of embryonic NO to nitrate is a very sensitive breeding indicator (V.Y. Titov et al., 2022). We assume that the effects observed with exogenous arginine injection in ovo in amounts not exceeding 1.0 µmol/g of egg weight are mainly due to an increase in the concentration of NO donor compounds in blood plasma. They can affect the tone of blood vessels and, possibly, the tone of the smooth muscles of the ducts of the digestive glands.
Текст обзорной статьи Аргинин и оксид азота (NO) в эмбриогенезе сельскохозяйственной птицы (обзор)
Для птиц аргинин — незаменимая аминокислота. Основная функция аргинина — участие в синтезе белков, а также различных биологически активных соединений и стимуляция секреции гормонов роста (1-3). Аргинин служит субстратом для синтеза оксида азота (4, 5).
Исследование выполнено за счет гранта Российского научного фонда ¹ 24-26-00148.
Установлено, что введение аргинина in ovo оказывает воздействие на постэмбриональное развитие даже если его количество более чем на порядок меньше изначально содержащегося в яйце (6-8). В ряде работ говорится, что добавки аргинина в корм влияют на рост и развитие цыплят (1, 9, 10). Это влияние пытались объяснить повышенной потребностью в аргинине в процессе эмбриогенеза и в 1-е сут постэмбрионального развития, а также эффектами оксида азота (NO), синтезируемого из аргинина (1, 9, 10). NO оказывает действие на тонус гладкой мускулатуры (11, 12), кровяное давление (13-15), дифференциацию тканей (16-18), их регенерацию и развитие (19, 20), на апоптоз (21-23) и экспрессию ряда генов, ответственных за рост и развитие (24-26).
Для более корректного определения причины наблюдаемых эффектов необходим контроль содержания аргинина и его метаболитов в тканях эмбриона и птицы после вывода. Необходимо установить, как изменяется концентрация аргинина в тканях в процессе развития эмбриона и при вводе экзогенного аргинина, а также определить влияние аргинина на содержание других аминокислот. Кроме того, важно выяснить, как изменяется концентрация производных аргинина и его физиологически значимых метаболитов в тканях в процессе развития эмбриона, как экзогенный аргинин влияет на эти показатели, на скорость эмбрионального и постэмбрионального роста и развития.
В представленной работе, обобщая данные разных авторов, в том числе свои собственные, мы попытались предложить механизмы развития эффектов, наблюдаемых в организме птицы при экзогенном введении аргинина в яйцо.
Содержание аргинина в курином яйце и изменение в процессе развития эмбриона. Общая концентрация аргинина в курином яйце составляет около 40 мкмоль/г (27, 28). Концентрация свободного аргинина в гомогенате яйца возрастает в процессе инкубации, достигая в конце 1,5-2,0 мкмоль/г против начальных 0,6 мкмоль/г (27, 29). Следовательно, пул свободного аргинина по мере развития эмбриона увеличивается, несмотря на его расход в синтетических процессах и незаменимость на эмбриональном этапе.
Повышенная концентрация свободного аргинина, вызванная экзогенным вводом аминокислоты в яйцо перед закладкой на инкубацию, сохраняется по крайней мере до 14-х сут (29). По-видимому, в яйце аргинин содержится в достаточном и даже избыточном количестве для обеспечения всех физиологических процессов в эмбрионе.
Тем не менее существуют публикации о влиянии аргинина в корме несушек на биохимические параметры плазмы, содержание иммуноглобулинов в плазме, активность NO-синтазы в тканях эмбрионов (30-32), массу эмбрионов и их мышц (33-35). Поскольку в этих работах не контролировалось содержание связанного и свободного аргинина в эмбрионе, есть основание предположить, что наблюдаемые эффекты были связаны с процессами, происходящими не в самом эмбрионе, а при формировании яйцеклетки.
В ряде работ говорится об эффектах введения аргинина in ovo на разных сроках инкубации (6, 36, 37). S. Subramaniyan с соавт. (6) вводили в яйца кур кросса Ross аргинин в дозах от 100 до 2500 мкг/100 мкл стерильного физиологического раствора на 8-е, 14-е и 18-е сут инкубации. Такие дозы, исходя из массы яиц 60 г, должны обеспечить увеличение концентрации аргинина в расчете на яйцо в пределах от 10 до 230 нмоль/г. Это меньше исходно содержащегося в яйце количества свободного аргинина и много- кратно меньше его общей концентрации (27-29). Тем не менее, согласно полученным результатам, при определенных дозах вводимого аргинина повышались выводимость, выживаемость, масса суточных цыплят, содержание фактора дифференциации миобластов (myoD1) и миогенина (myog) в грудных мышцах. При этом эффекты сильно и несистемно менялись в зависимости от дозы и срока ввода. Объяснения этим феноменам в статье нет.
Согласно результатам, полученным Y. Li с соавт. (7), введение аргинина в амнион у кур кросса Ross на 16-е сут в дозах, увеличивающих общую концентрацию аргинина на величину от 37,5 до 600 нмоль/г, оказывало отрицательный эффект на живую массу и массу мышц.
Аргининовый парадокс. Для того чтобы иметь хотя бы гипотезу, объясняющую сущность рассмотренных выше эффектов, нужен количественный анализ содержания аргинина и физиологически значимых продуктов его метаболизма (рис. 1). Один из них — оксид азота (NO), который синтезируется из аргинина под воздействием фермента NO-синтазы (3840).
Аргинин
Синтез белковых
NO-синтаза
молекул
Аргиназа
(у птиц отсутствует) аргининдекарбоксилаза, аргининллицинамидотрансфераза
Белок
NO, цитруллин
Полиамины, пролин, глутамат, мочевина, креатин
Рис. 1. Основные пути метаболизма аргинина (1-3) .
Здесь мы сталкиваемся с феноменом, известным как аргининовый парадокс. Концентрация субстратного насыщения для эндотелиальной NO-синтазы составляет несколько мкмоль/л. В то же время плазма крови и другие ткани организма содержат свободный аргинин в концентрации десятков и сотен мкмоль/л (41). Тем не менее экзогенный ввод аргинина способствовал проявлению таких эффектов, как снижение артериального давления, которое может быть связано с NO (12, 41, 42).
Одно из предположений о механизме этих явлений связано с локализацией NO-синтазы в клетке. Например, NO-синтаза может быть локализована совместно с другим ферментом, использующим аргинин как субстрат, особенно если у этого фермента концентрация насыщения на несколько порядков выше, чем у NO-синтазы (42, 43). Также может иметь место неодинаковая концентрация аргинина в различных компартментах клетки (42, 44), наличие и состояние мембранного транспортера аргинина в клетки и конкурентов аргинина за этот транспортер (45). Однако эти гипотезы не удалось подтвердить экспериментально (42, 45).
Существует предположение об эндогенных конкурентных ингибиторах NO-синтазы. Таковыми служат производные аргинина диметиларгинин симметричный (СДМА) и асимметричный (АДМА). Это не протеиногенные аминокислоты, но, конкурируя с аргинином, они способны увеличить концентрацию субстратного насыщения фермента (46, 47). Считается, что эти соединения образуются посредством метилирования аргинина метилтранс-феразами (46). На модельных системах показано, что СДМА и АДМА могут повысить концентрацию насыщения NO-синтазы до величин, которые регистрируют в условиях in vivo (48). Наблюдается увеличение концентрации этих соединений при почечной недостаточности (49). Концентрация АДМА в плазме взрослого человека равна 0,4-0,6 мкмоль/л (46, 49). В плазме но- ворожденных концентрация АДМА значительно выше — около 1 мкмоль/л, но уже на 2-е сут после рождения показатель нормализуется (46). АДМА и СДМА выводятся из организма с мочой, АДМА также претерпевает ферментативное разложение (50, 51). Эти соединения есть и у птиц (47).
Таким образом, образование метилпроизводных аргинина и их вывод из организма могут быть регуляторным механизмом в синтезе NO. При этом интерес представляет не синтез сам по себе, а физиологические эффекты NO в организме.
Оксид азота и его метаболизм в живых тканях. Поскольку NO представляет собой химически активную и короткоживущую в свободном состоянии молекулу, для успешной реализации его физиологических эффектов необходимо выполнение двух условий: предотвращение окисления NO до токсического нитрита и взаимодействия с физиологически не значимыми мишенями.
В настоящее время нет единого представления об этих механизмах, поскольку для формулировки гипотез нужен оперативный контроль всего состава нитро- и нитрозосоединений в тканях и его изменений. Однако это представляет некоторую техническую трудность: используемые методы имеют либо недостаточную избирательность, либо малую чувствительность. Все они основаны на нитровании или нитрозировании специфических реагентов и требуют кислой среды, в которой нитрозильные комплексы железа могут разрушаться и окисляться до нитрита. S-нитрозотиолы (RSNO) и динитрозильные комплексы железа (ДНКЖ) также частично определяются как нитрит (52, 53). Эти методы не дают представления об особенностях синтеза и метаболизма NO (53-55).
Живые ткани содержат так называемые соединения-доноры NO, к которым относятся S-нитрозотиолы (56-58) и различные нитрозильные комплексы железа (59, 60). Моноядерный динитрозильный комплекс железа, содержащий тиолатные лиганды, парамагнитен. Однако преобладающий в тканях биядерный комплекс диамагнитен и не может быть обнаружен методом электронного парамагнитного резонанса (ЭПР) (61-63). О синтезе и метаболизме NO судят по суммарному содержанию нитрата и нитрита, исходя из представления о том, что NO — активная молекула, которая быстро метаболизируется, а соединения-доноры NO нестабильны и быстро распадаются (54, 55). При этом в некоторых тканях концентрация доноров NO сравнима с концентрацией нитрата или даже выше нее (53, 64).
В связи с описанными трудностями до сих пор нет ясного представления о механизмах взаимодействия NO с физиологическими мишенями и о том, как обеспечивается специфичность этого воздействия, а также о метаболизме NO во многих тканях и о характере его изменения при некоторых патологиях. Непонятно, снижается ли синтез NO при заболеваниях, связанных с увеличением кровяного давления, или происходит инактивация NO до момента взаимодействия с мишенью (65-67).
Разработанный в Российском научно-исследовательском медицинском университете им. Н.И. Пирогова совместно с Институтом химической физики РАН ферментный сенсор основан на обратимом ингибировании фермента каталазы всеми нитрозосоединениями с примерно одинаковой эффективностью, которая возрастала на два порядка в присутствии хлорида в плазменной концентрации, а также бромида и тиоцианата (53, 68, 69).
Различные нитрозосоединения теряют ингибирующий эффект под воздействием веществ, специфичных для каждой их группы (53, 70, 71). Такой сенсор не может определить детали строения тех или иных веществ, но способен с высокой точностью (50 нмоль/л) и без подготовки образца оце-212
нить содержание нитрозильных комплексов железа как соединений, теряющих способность ингибировать в системе, содержащей ловушку NO и хелатор железа, который разрушает комплекс.
S-нитрозотиолы определялись как соединения, теряющие ингибирующий эффект под воздействием солей ртути либо как приобретающие свойства нитрозильных комплексов железа под воздействием закисного железа (70, 71). Высокомолекулярные нитросоединения, способные трансформироваться в нитрозильные комплексы железа, определялись как соединения, приобретающие ингибирующий эффект под воздействием закисного железа и тиолов, а также под воздействием треххлористого ванадия, восстанавливающего эти соединения до нитрозосостояния. Нитрит и нетиолатные нитрозосоединения не теряли ингибирующий эффект под воздействием солей ртути и системы ловушка NO—хелатор (70, 71).
При помощи сенсора показано, что нитрит и нетиолатные нитрозосоединения в норме в большинстве тканей находятся в следовых концентрациях — менее 100 нмоль/л (53, 72). Похожие данные получали другие исследователи при помощи различных модификаций хроматографического метода (73, 74). При этом концентрация соединений-доноров NO может достигать десятков и даже сотен мкмоль/л (53, 64). То есть механизмы, препятствующие окислению NO до нитрита, в норме существуют. Мы предполагаем, что вновь синтезированная молекула NO сразу включается в состав соединений-доноров (RSNO, нитрозильных комплексов железа), а последние непосредственно взаимодействуют с физиологической мишенью (53) (рис. 2).
t RSNO _______ Взаимодействие ______ Окисление до нитрата
NO ММВ^^ ДНКЖ ■■■■■► с физиологической ЯММ^
RNO, мишенью Вывод из организма
Рис. 2. Предполагаемый механизм предотвращения окисления NO до нитрита. Синтезированная в тканях молекула NO включается в состав так называемых соединений-доноров: S-нитрозотиолов (RSNO), динитрозильных комплексов железа (ДНКЖ), некоторых высокомолекулярных нитратов (RNO 2 ). Соединения-доноры продлевают физиологическое время жизни NO и непосредственно взаимодействуют с физиологическими мишенями (53).
Сами соединения-доноры не претерпевают спонтанного распада с высвобождением NO. Высвобождение происходит при наличии мишени, имеющей химическое сродство к NO, и агента, разрушающего соединение-донор. Предположительно таким агентом может быть часть апофермента мишени. В роли донора, непосредственно взаимодействующего с мишенью, как полагают некоторые исследователи, выступает динитрозильный комплекс железа (53, 75) (рис. 3).
Фактор, вызывающий деструкцию комплекса
▼ NO
RSNO ^^^ ДНКЖ ^^^^ Мишень
Рис. 3. Предполагаемый механизм взаимодействия соединений-доноров NO с физиологической мишенью . RSNO — S-нитрозотиолы, ДНКЖ — динитрозильные комплексы железа. Соединения-доноры NO спонтанно не распадаются с высвобождением NO. Оксид азота высвобождается из них под влиянием различных факторов, вызывающих деструкцию соединения, при наличии веществ, имеющих химическое сродство к NO. Последние, предположительно, могут выступать и в роли деструкторов (53, 75).
Мы установили, что в ряде тканей встречаются нитрозильные комплексы железа, не имеющие тиолатных лигандов Fe(NO)n и приобретающие свойства динитрозильных комплексов железа, содержащих тиолатные лиганды (ДНКЖ/SH), после добавления в среду тиолов. Такие комплексы были получены и на модельных системах (53).
Взаимосвязь содержания аргинина и производных оксида азота (NO) в курином эмбрионе. Известно, что эмбриогенез сопровождается интенсивным синтезом оксида азота (26, 76, 77). У птиц эмбрион развивается отдельно от материнского организма и представляет сравнительно замкнутую систему. Из данных о концентрации аргинина в яйце (27-29) следует, что эта аминокислота содержится в избытке для физиологических процессов. О взаимосвязи содержания аргинина и интенсивности синтеза NO до сих пор не было сведений, поскольку для этого необходимо определение содержания всех основных метаболитов NO, что представляет техническую сложность.
Изначально в гомогенате снесенного куриного яйца содержится несколько мкмоль/л соединений-доноров NO и нитрата. По мере развития эмбриона доноры NO и нитрат накапливаются в тканях. К 3-м сут их концентрация в гомогенате достигает 120-150 мкмоль/л. Большая часть доноров NO (более 90 %) представлены соединениями, определяемыми как высокомолекулярные нитраты (RNO 2 ). Соединения-доноры NO находятся в амнионе, а нитрат за его пределами. После 11-х сут концентрация нитро- и нитрозосоединений в гомогенате яйца резко возрастает до нескольких сотен мкмоль/л к моменту вывода. Соединения-доноры NO на этом этапе представлены в основном динитрозильными комплексами железа, содержащими тиоловые лиганды (ДНКЖ/SH). Это связано с тем, что, начиная с 11-х сут, доноры NO накапливаются в клетках тканей зародыша, где преобладают ДНКЖ/SH. Концентрация RSNO, RNO 2 незначительна (29).
Из соотношения концентраций аргинина и производных NO в яйце следует, что на синтез NO тратится около 2 % аргинина (29). В эмбрионах одних пород, линий и кроссов NO накапливается в тканях эмбриона в составе соединений-доноров, в эмбрионах других преимущественно претерпевает окисление до нитрата. Последнее происходит в основном в эмбрионах птицы мясного направления продуктивности. В них окисляется более 90 % NO (29). При этом в любом случае NO в птичьем эмбрионе синтезируется в многократно больших количествах, чем необходимо для процессов жизнедеятельности.
Блокатор NO-синтазы N ω -нитро-L-аргинин, введенный в яйцо через воздушную камеру как до закладки на инкубацию, так и на 11-е или 12-е сут инкубации, способствовал снижению содержания доноров NO и нитрата в гомогенате куриного эмбриона (рис. 4), не влияя на их соотношение (29).
Аргинин, введенный вместе с нитроаргинином в соотношении 10:1, полностью предохранял пул производных NO, но при этом его введение не способствовало увеличению содержания этих веществ. Эти данные подтверждают тот факт, что аргинин в яйце находится в концентрации насыщения для NO-синтазы (см. рис. 4). По нашим данным, экзогенный ввод аргинина даже в количестве, удваивающем общий пул свободной аминокислоты, никак не влиял на содержание метаболитов оксида азота в гомогенате эмбриона (см. рис. 4). То есть после экзогенного ввода аргинина не происходит ни интенсификации синтеза NO, ни, по-видимому, значительной интенсификации других процессов, связанных с расходом аргинина, о чем можно судить по сохранению его повышенной концентрации после экзогенного ввода в яйцо (29).
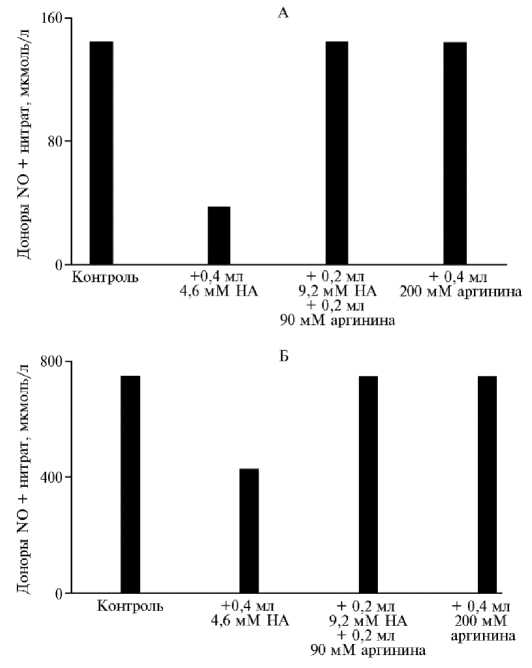
Рис. 4. Эффект аргинина и конкуретного блокатора NO-синтазы N ω -нитро-L-аргинина (НА) на содержание метаболитов NO в гомогенате куриного яйца при введении реагентов перед закладкой на инкубацию (А) и на 11-е сут инкубации (Б) . Измерения проводили на 7-е (А) и на 14-е сут инкубации (Б) (29, 78, 79) .
Содержание свободного аргинина и производных NO в крови и тканях куриного эмбриона. В плазме 18-суточного куриного эмбриона концентрация свободного аргинина составляла 460 мкмоль/л, 9-суточного — 200-300 мкмоль/л. В то же время содержание доноров NO в плазме возрастало за этот период в несколько раз (78).
В плазме эмбриона содержались соединения, определяемые как высокомолекулярные нитраты (RNO 2 ), в концентрации до нескольких ммоль/л. Также были обнаружены соединения с признаками нитрозильных комплексов железа, не содержащих тиолатные лиганды — Fe(NO) n в концентрации десятков и сотен мкмоль/л, нитрозотиолы (RSNO) и ДНКЖ/SH в концентрации нескольких мкмоль/л. Нитрата (NO 3- ) было многократно меньше, чем RNO 2 (табл. 1) (29).
Cразу после вывода концентрация производных NO резко снижалась (см. табл. 1). На 2-е-3-и сут в плазме было обнаружено 8-20 мкмоль/л Fe(NO) n , следовые количества RNO 2 (табл. 2), десятки мкмоль/л нитрата (78, 79). То есть в куриных эмбрионах имеет место высокая концентрация производных NO, обусловленная, по-видимому, интенсивным синтезом NO и менее интенсивным, чем у взрослых особей, выведением его из плазмы крови (29, 78).
Мы показали, что концентрация производных NO в плазме крови есть результат динамического равновесия между процессами синтеза, окисления и выведения из кровяного русла. Характерно, что содержание нит- рата, как и доноров NO, снижается после ввода блокатора NO-синтазы нитроаргинина и восстанавливается спустя несколько минут у взрослой курицы и в течение нескольких суток у эмбриона (29, 78).
1. Концентрация доноров NO и свободного аргинина в плазме крови у эмбрионов и цыплят породы мини-мясная в зависимости от времени развития (29, 78, 79)
Срок инкубации, сут ] Аргинин, мкмоль/л ] Fe(NO) n , мкмоль/л RNO 2 , ммоль/л
|
9 |
277,7±34,3 |
184,6±10,9* |
1,2±0,3* |
|
12 |
371,5±25,9* |
1650,0±230,4* |
12,3±0,9* |
|
14 |
495,8±35,8* |
1380,9±180,5* |
9,1±0,9* |
|
18 |
469,6±33,1* |
350,5±31,2* |
7,2±0,6* |
|
1-суточные цыплята |
299,4±29,4 |
38,2±4,7 |
0,037±0,005 |
Примечание. Fe(NO) n — нитрозильные комплексы железа, не содержащие тиоловые лиганды, RNO 2 — высокомолекулярные нитросоединения.
* p < 0,05 относительно 1-суточных цыплят.
2. Содержание свободного аргинина и доноров NO в плазме крови у эмбрионов и цыплят кросса Смена 9 при введении аргинина in ovo на 14-е сут инкубации (79)
|
Группа |
Аргинин, мкмоль/л |
Fe(NO) n , мкмоль/л |
RNO 2 , ммоль/л |
|
0,3 мл физраствора |
Эмбрионы, 18 387,5±22,4 |
-е сут 80,4±4,1 |
6,5±0,4 |
|
0,3 мл 3,0 ммоль/л аргинина |
408,1±27,4 |
275,8±15,6* |
19,8±1,8* |
|
0,3 мл 30,0 ммоль/л аргинина |
392,3±24,7 |
1225,6±51,5* |
24,5±2,2* |
|
0,3 мл 300,0 ммоль/л аргинина |
1102,1±64,3* |
7544,3±290,8* |
26,6±2,8* |
|
0,3 мл 300,0 ммоль/л аргинина перед закладкой на инкубацию |
807,6±35,5* |
71,8±3,3 |
6,3±0,4 |
|
Ц |
ыплята, 1- е сут после вывода |
||
|
0,3 мл физраствора |
358,8±28,3 |
37,2±3,3 |
< 0,1 |
|
0,3 мл 300,0 ммоль/л аргинина |
399,7±29,9 |
432,3±30,3* |
0,3±0,1* |
|
Ц |
ыплята, 3- и сут после вывода |
||
|
0,3 мл физраствора |
209,4±12,4 |
14,4±0,9 |
< 0,1 |
|
0,3 мл 300,0 ммоль/л аргинина |
261,5±13,3 |
44,8±2,0* |
< 0,1 |
* p < 0,05 относительно групп, в которых вводился физиологический раствор.
Очевидно, что рецепторы NO в эндотелии эмбриона менее чувствительны или менее доступны, чем у взрослой особи, поскольку у последней повышение концентрации доноров NO до нескольких десятков мкмоль/л вызывало обморочное состояние, связанное, по-видимому, с гипотензивным эффектом NO (29).
Высокая концентрация доноров NO в плазме эмбриона наблюдается и у крупного рогатого скота. После рождения она постепенно снижается, достигая значений, характерных для взрослых особей, через 7-10 сут (29). У птиц нормализация содержания депонированного NO в плазме наступает непосредственно перед выводом (29).
В работах разных авторов аргинин вводился in ovo в разные сроки. Однако невозможность контролировать состав нитро- и нитрозосоединений плазмы не позволяла получить ясную информацию об эффектах экзогенного аргинина на синтез NO.
В наших экспериментах ввод аргинина в яйцо через воздушную камеру перед закладкой на инкубацию в количестве 0,3 мл 300 ммоль/л раствора не приводил к увеличению содержания нитро- и нитрозосоединений в плазме, регистрируемого на 18-е сут инкубации (см. табл. 2), а также на 7-е сут (78, 79). При этом содержание аргинина в плазме на 18-е сут более чем вдвое превышало контроль (см. табл. 2). Не наблюдалось увеличения содержания нитро- и нитрозосоединений и в мышечном каркасе эмбрионов по сравнению с контролем (29), и в других тканях (см. рис. 4). Таким образом, вероятно, подтверждается предположение о том, что аргинин в тканях эмбриона находится в концентрации насыщения для NO-синтазы.
Однако ввод аргинина in ovo на 14-е сут в такой же дозе приводил как к увеличению его содержания в плазме крови эмбриона, так и к повышению концентрации метаболитов NO. Содержание последних увеличивалось и при введении аргинина в меньших (на порядок и на два порядка) концентрациях, которые не приводили к существенному увеличению его концентрации в плазме (см. табл. 2). Можно предположить, что к этому времени появляются субстраты-конкуренты. Однако не ясно, почему аргинин, введенный до закладки на инкубацию, не способствовал продукции NO, находясь и на 18-е сут в плазме крови в повышенной по сравнению с контролем концентрации (см. табл. 2). То есть повышение концентрации аргинина в плазме на 18-е сут при его введении in ovo на 14-е сут и перед закладкой на инкубацию не есть эквивалентные процессы. Такую разницу можно объяснить либо гибким изменением концентрации субстратов-конкурентов в зависимости от срока инкубации и концентрации аргинина, либо тем, что введенный аргинин спустя какое-то время перестает быть доступным для NO-синтазы.
Повышенная концентрация доноров NO сохранялась до 3-х сут после вывода, в то время как концентрация аргинина нормализовалась уже к моменту вывода (см. табл. 2).
Таким образом, ввод аргинина на заключительной стадии предплод-ного периода способствует повышению содержания соединений-доноров NO в плазме крови. В мышечном каркасе, печени и тканях желудочно-кишечного тракта содержание депонированного NO не повышается (79). При этом конкурентный ингибитор NO-синтазы N ω -нитро-L-аргинин снижет концентрацию метаболитов NO в этих тканях, а введение аргинина одновременно с N ω -нитро-L-аргинином предотвращает снижение (29, 79). Следовательно, в этих тканях имеет место исходное содержание аргинина в концентрации насыщения для NO-синтазы.
Можно предположить, что физиологические эффекты разового ввода аргинина в яйцо в количествах, существенно не изменяющих его содержание в тканях, связаны с увеличением концентрации соединений-доноров NO в плазме крови. Повышенная концентрация доноров NO, вероятно, способствует снижению тонуса гладкой мускулатуры сосудов и пищеварительных протоков, стимулирует синтез пищеварительных ферментов (29). Так, в наших исследованиях на эмбрионах кроссов Ross 308 и Смена 9 введение аргинина на 14-е сут инкубации даже в количестве 0,3 мл 5,0 ммоль/л раствора приводило к достоверному увеличению скорости роста цыплят после вывода (до нескольких процентов), в то время как введение в несколько раз больших доз перед закладкой на инкубацию такого эффекта не имело (29, 79).
Влияние аргинина и NO на экспрессию генов, ответственных за миогенез. Мы установили, что в эмбрионах сельскохозяйственной птицы мясного направления продуктивности, начиная со 23-х сут эмбриогенеза, происходит окисление депонированного NO до нитрата, который накапливается за пределами амниона, а после формирования аллантоиса — в нем. Использование света при инкубации, способствующее повышению скорости постэмбрионального роста, также приводит к интенсификации окисления эмбрионального NO (29, 82).
В эмбрионах бройлеров, бойцовых кур, а также быстрорастущих пород и линий перепелов, цесарок, индеек, страусов более 90 % всего синтезированного в эмбрионе NO окисляется до нитрата. Напротив, в эмбрионах яичных форм окисление NO незначительное. Скрещивания различных пород и анализ потомства в первом и втором поколениях показали, что этот признак наследуется. Причем он определяется как минимум двумя генами (29).
Установлено, что окисление происходит в мышечной ткани. Состав нитро- и нитрозосоединений печени, плазмы крови и желудочно-кишечного тракта достоверно не различается у эмбрионов мясного и яичного направлений продуктивности (29, 82).
Известно немало публикаций о влиянии NO на экспрессию ряда генов (83-85). Исследованы, в частности, взаимосвязь между содержанием NO и экспрессией генов (86, 87), эффекты ингибиторов NO-синтазы, а также доноров NO. Как показано при помощи ПЦР-РВ и вестерн-блот-тинга, ингибиторы снижали экспрессию ряда генов, доноры NO — восстанавливали (26). Это гены миогенной дифференциации 1 ( myoD1 ), фактора миогенеза 5 ( Myf5 ), фактора пролиферации миоцитов 2с ( mef2c ), миогенина ( myog ), миозина ( mhy1 ) (26).
Есть мнение, что оксид азота блокирует метилазу гистоновых белков, что оказывает влияние на экспрессию генов (83, 84, 88). Предположительно этот эффект может реализоваться тремя путями: через непосредственное воздействие на фермент, связывание железа, необходимого как кофактор, и регуляцию экспрессии ферментов, ответственных за метилирование (85, 89).
На эмбрионах кур и перепелов при помощи ферментного сенсора, позволяющего контролировать состав метаболитов NO в тканях эмбриона, и метода ПЦР-РВ мы исследовали взаимосвязь содержания NO в тканях и экспрессии семи генов, участвующих в процессе миогенеза: генов фактора пролиферации миоцитов 2с ( mef2c ), миогенной дифференциации 1 ( myoD1 ), фактора миогенеза 5 ( myf5 ), миозина ( mhy1 ), миогенина ( myog ), миостатина ( MSTN ), гонадотропного гормона.
Блокирование синтеза NO, вызывающее снижение содержания депонированного NO в тканях на 50-70 %, приводит к изменению экспрессии большинства исследованных генов в несколько раз. В основном наблюдалось снижение экспрессии генов, в частности гена, опосредующего синтез миостатина ( MSTN ), ответственного за подавление роста и дифференциации мышечной ткани. Блокирование синтеза NO перед закладкой яйца на инкубацию, а также на 11-е и 14-е сут приводило к снижению содержания 218
депонированного NO в тканях эмбриона и снижению экспрессии указанных генов (90, 91). Применение аргинина вместе с конкурентным ингибитором NO-синтазы нитроаргинином в соотношении 10:1 полностью снимало эффекты ингибитора на содержание депонированного NO и на экспрессию генов. Сам аргинин достоверных эффектов на эти показатели не оказывал (91).
Сравнительный анализ экспрессии вышеназванных генов у эмбрионов показал, что в эмбрионах мясных форм на 14-е сут инкубации экспрессия большинства этих генов была в среднем ниже, чем в яичных. При этом наблюдались существенные различия между экспрессией у эмбрионов разных пород, характеризующихся одинаковой степенью окисленности NO (91).
Таким образом, оксид азота в птичьем эмбрионе прежде всего может играть роль регулятора роста мышечной ткани, что важно для быстрорастущих форм, поскольку пролиферация миобластов происходит на эмбриональной стадии развития. По-видимому, имеет место совокупный эффект на экспрессию сразу нескольких генов.
Регуляция может осуществляться посредством активации механизмов окисления NO до нитрата, что происходит в эмбрионах быстрорастущих форм (рис. 5). Окисление начинается на 2-е-3-и сут эмбриогенеза и продолжается в течении всего эмбрионального развития. Данные исследований эмбрионов у гибридов различных пород позволяют предположить, что механизмы этого окисления обусловлены генетически и определяются несколькими генами (29, 82).
Генетически обусловленное окисление NO
Аргинин "■♦ NO ка^ Доноры NO ^^■^^■^■^ Нитрат
Экспрессия генов
Рис. 5. Предполагаемый механизм регуляции экспресии генов, ответственных за миогенез, в эмбрионах птиц (29, 82, 90) .
Следует упомянуть еще один возможный механизм регуляции содержания аргинина в тканях, синтеза NO и его эффектов. Увеличение содержания аргинина в рационе способствует снижению усвояемости лизина (9294), поскольку аргинин и лизин конкурируют за мембранный переносчик (95, 96). Эффект конкуренции аргинина с другими аминокислотами может иметь место при непрерывном вводе, обеспечивающем длительное повышение его концентрации.
Итак, аргинин в курином яйце исходно содержится в количестве, достаточном для формирования тканей зародыша и обеспечения физиологических процессов, протекающих в эмбрионе. Экзогенный ввод аргинина в яйцо в концентрациях, сравнимых с исходной, может способствовать увеличению содержания доноров NO. Причем это увеличение происходит только в плазме крови после ввода аргинина в яйцо на заключительной стадии предплодного периода. Оксид азота способствует расслаблению гладкой мускулатуры сосудов, пищеварительных протоков. Возможно, именно этот эффект приводит к некоторому увеличению скорости прироста живой массы. Исходя из данных, полученных разными авторами, оксид азота оказывает влияние на экспрессию генов, в частности ответственных за миогенез. В эмбрионах как минимум пяти видов птиц (кур, цесарок, перепелов, индеек и страусов) степень окисленности депонированного NO коррелирует с мясной продуктивностью взрослых особей. Однако возможность искусственно регулировать этот процесс ограничена рядом проблем. Амниотическая мембрана непроницаема для нитро- и нитрозосоединений, поэтому в ткани эмбриона могут попасть только доноры NO, введенные до закладки яиц на инкубацию. Аргинин в тканях эмбриона содержится в концентрации насыщения для NO-синтазы. Конкурентные блокаторы синтеза NO действуют непродолжительное время. В наших экспериментах введение нитроаргинина in ovo не оказывало достоверного влияния на скорость роста и инкубационные характеристики. При этом степень окисленности NO в тканях эмбриона служит важным селекционным показателем. Мы предполагаем, что все эффекты аргинина при его однократном введении в организм птицы на эмбриональной и постэмбриональной стадии в основном связаны с синтезом NO и его физиологическими эффектами.
1 ФГБНУ ФНЦ Всероссийский научно-исследовательский и технологический институт птицеводства,
