Археозоологические материалы XI–XVII веков из раскопок Земляного городища в Старой Ладоге

Автор: Бачура О.П., Лобанова Т.В., Григорьева Н.В., Лапшин В.А.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Рубрика: Естественнонаучные методы в археологических исследованиях
Статья в выпуске: 278, 2025 года.
Бесплатный доступ
В работе описана остеологическая коллекция из раскопок 2007 и 2011 гг. Земляного городища в Старой Ладоге. Изучено более 28 тыс. костных остатков из культурных отложений XI–XVII вв. Проведен анализ видового состава и соотношения костей животных, описан анатомический состав остатков, а также возрастная структура забоя домашнего скота. Показано, что основой хозяйства у населения Ладоги было домашнее хозяйство. Наибольшее значение для населения города имел крупный рогатый скот и свинья. Структура домашнего хозяйства типична для русских городских поселений XI‒XVII вв. Охота и рыболовство имели место, но не играли большой роли в жизнеобеспечении населения. На протяжении XI– XVII вв. у жителей посада Ладоги наблюдается возрастание роли домашних животных в хозяйстве, а промысловая деятельность практически утрачивает свое значение. Кроме того, увеличивается молочное направление в скотоводстве.
Северо-Запад России, Старая Ладога, Земляное городище, древнерусское время, средние века, домашние животные, костные остатки
Короткий адрес: https://sciup.org/143184298
IDR: 143184298 | DOI: 10.25681/IARAS.0130-2620.278.396-412
Archaeozoological Materials of the 11th–17th Centuries from the Excavations of the Zemlyanoe Hillfort in Staraya Ladoga
The paper describes an osteological assemblage from the 2007 and 2011 excavations of the Zemlyanoe Hillfort in Staraya Ladoga. A total of more than 28 000 bone remains from the occupation layers dating to the 11th–17th centuries have been examined. The analysis of the animal species composition and percentages of animal bones has been performed. The anatomical composition of the remains and the age groups of the slaughtered domestic animals have been described. The paper shows that economy of the Ladoga population was based on household production. Cattle and pigs were most important animals for the urban population. The components of household production are typical of Russian urban settlements of the 11th–17th centuries. Local population was also engaged in hunting and fishing; however, these activities did not play a major role in subsistence economy. Throughout the 11th–17th centuries the role of domestic animals in the economy of the people who lived in the trading quarter around the city increased, while importance of fishing and hunting basically declined. Besides, the number of domestic animals raised for milk increased.
Текст научной статьи Археозоологические материалы XI–XVII веков из раскопок Земляного городища в Старой Ладоге
Земляное городище в Старой Ладоге – многослойный археологический памятник, историю которого можно разделить на два периода: существование неукрепленного поселения VIII‒XVI вв. и перекрывающее его укрепление бастионного типа конца XVI – XVII в. (отсюда и название Земляное городище, появившееся в XVII в.). Культурный слой поселения, в свою очередь, делится на верхние слои гумуса XI‒XVI вв. и нижние слои так называемого мокрого
1 Статья подготовлена в рамках гранта Российского научного фонда № 23-18-00515 «Формирование ранней городской структуры и культурного ландшафта Северной Руси на материале археологического комплекса Старой Ладоги».
слоя VIII‒X вв., сохранившего органику, в том числе остатки деревянных построек, что позволяет использовать метод дендродатирования ( Лапшин , 2019). Археологические исследования на Земляном городище ведутся уже более ста лет. Традиционно наибольший интерес привлекала нижняя часть отложений. Это относится и к остеологическим находкам. В обобщенном виде была описана коллекция костей, собранная в ходе раскопок 1949 г. в культурном слое IX‒X вв. В. И. Равдоникасом в центральной части городища ( Равдоникас , 1949; Цалкин , 1956). Также остеологическим материалам раннего Средневековья посвящена статья А. К. Каспарова по результатам раскопок в северной части Земляного городища ( Каспаров , 1997). В настоящей статье впервые дается описание остеологических материалов из верхней части культурного слоя XI–XVII вв. В начале этого периода Ладога начинает развиваться как городской центр древнерусского времени, превращаясь в типичный средневековый город Северо-Западного региона.
Хронология и стратиграфия
Представленная в данной публикации коллекция была собрана при археологических исследованиях раскопа 4 (2007 г.) и раскопа 5 (2011 г.) Староладожской археологической экспедицией ИИМК РАН под руководством А. Н. Кирпичникова. Последовательно исследованные площади располагаются в южной части Земляного городища, вдоль внутренней части деревоземляной стены. Культурный слой представлял собой сухой гумус черного и серого цвета с остатками деревянных конструкций разной степени сохранности.
В 2007 г. мощность исследованных напластований составила 35–50 см ( Кирпичников , 2008а; 2008б). В 2011 г. была изучена толща около 1 м культурного слоя от самых поздних напластований до XI в. ( Кирпичников , 2012).
Традиционно в Ладоге для описания напластований культурного слоя используется схема горизонтов, сформулированная В. И. Равдоникасом. Он ввел в научный оборот семь горизонтов: от « а » до « ж » ( Равдоникас , 1949. С. 10–12), которые отражают культурно-хронологические изменения поселения. Поздний этап развития поселения и сооружение земляной крепости относятся к горизонтам а–г по схеме В. И. Равдоникаса. Впоследствии схема горизонтов была уточнена и дополнена в ходе продолжения исследовательских работ.
А. Н. Кирпичников при полевых и фиксационных работах также использовал литерные обозначения горизонтов. Его методика сводилась к тому, что все изученные за сезон слои и прослойки, пустые или содержащие строительные остатки, назывались горизонтами и получали буквенное обозначение, ежегодно начиная с литеры «А». Хотя, по сути, являлись просто уровнями полевой фиксации. Этот методический подход подробно изложен в одной из публикаций руководителя работ ( Кирпичников, Назаренко , 1997. С. 63). В настоящей работе «горизонты», по В. И. Равдоникасу и А. Н. Кирпичникову, синхронизированы (табл. 1).
Таблица 1. Таксономический состав костных остатков животных из раскопок Земляного городища
|
Раскоп (год) |
4 (2007) |
5 (2011) |
||||||
|
Горизонт, по А. Н. Кирпичникову |
В |
Б |
А |
Г низ |
Г верх |
В |
Б |
А |
|
Горизонт, по В. И. Равдоникасу |
Горизонт погребений раннего могильника при Климентовской церкви, г |
г |
б |
|||||
|
Хронология, вв. |
XI–XIII |
XIV– XV |
XV– XVI |
XVI– XVII |
||||
|
Крупный рогатый скот – Bos taurus |
740 |
276 |
761 |
2636 |
830 |
3648 |
2289 |
672 |
|
Свинья – Sus scrofa domecticus |
531 |
247 |
511 |
894 |
288 |
822 |
382 |
86 |
|
Мелкий рогатый скот – Capra et Ovis |
84 |
56 |
132 |
594 |
147 |
562 |
251 |
73 |
|
Лошадь – Equus caballus |
19 |
4 |
24 |
20 |
7 |
73 |
44 |
10 |
|
Собака – Canis familiaris |
12 |
8 |
8 |
29 |
5 |
19 |
3 |
5 |
|
Кошка – Felis catus |
– |
1 |
1 |
9 |
2 |
14 |
7 |
1 |
|
Заяц-беляк – Lepus timidus |
— |
— |
4 |
37 |
5 |
26 |
19 |
10 |
|
Белка – Sciurus vulgaris |
– |
– |
– |
2 |
1 |
– |
– |
– |
|
Бобр – Castor fiber |
18 |
2 |
18 |
72 |
8 |
9 |
7 |
1 |
|
Волк – Canis lupus |
1 |
– |
2 |
3 |
2 |
1 |
– |
– |
|
Лисица – Vulpes vulpes |
16 |
6 |
20 |
45 |
3 |
5 |
3 |
3 |
|
Куница – Martes martes |
– |
– |
– |
7 |
– |
– |
– |
– |
|
Медведь бурый – Ursus arctos |
– |
– |
1 |
4 |
– |
1 |
2 |
– |
|
Нерпа кольчатая – Phoca hispida ladogensis |
– |
– |
1 |
2 |
– |
3 |
1 |
– |
|
Кабан – Sus scrofa |
– |
– |
– |
1 |
– |
1 |
1 |
– |
|
Олень северный – Rangifer tarandus |
– |
– |
– |
1 |
– |
2 |
– |
– |
|
Лось – Alces alces |
8 |
— |
29 |
73 |
21 |
66 |
14 |
1 |
|
Млекопитающие, ближе неопределимые |
264 |
43 |
405 |
2731 |
660 |
2447 |
1209 |
243 |
|
Рыба |
80 |
110 |
217 |
397 |
63 |
172 |
131 |
20 |
|
Птица |
135 |
122 |
198 |
330 |
59 |
137 |
73 |
12 |
|
Итого |
1908 |
875 |
2332 |
7887 |
2101 |
8008 |
4436 |
1137 |
Материалы и методы
Общее количество исследованных остатков животных составило 28 595 экземпляров (табл. 1). Коллекция хранится в основных фондах музея Института экологии растений и животных УрО РАН под № 1852. Неопределимые остатки млекопитающих, а также кости птиц и рыб были идентифицированы до класса. Определимые кости млекопитающих были диагностированы до вида с помощью эталонной коллекции музея Института экологии растений и животных УрО РАН.
Для оценки раздробленности остатков было подсчитано количество целых и фрагментированных костей млекопитающих. К категории «целые» были отнесены действительно целые кости, а также кости от молодых животных, у которых отсутствует один или оба эпифиза, а также сами эпифизы. Наблюдаются некоторые различия в степени раздробленности костей млекопитающих между раскопами, даже в синхронных горизонтах. В раскопе 5 количество целых костей колеблется от 15 до 20 %, и в большинстве горизонтов высока доля неопределимых остатков (22–38 %). В раскопе 4 довольно много целых костей (26 %), а доля неопределимых не превышает 17 %. На наш взгляд, это обусловлено методикой сбора. В 2011 г. был собран весь остеологический материал, а в 2007 г. собирали преимущественно целые крупные кости. Об этом же свидетельствует малое общее количество костей, собранных с трех горизонтов в раскопе 4, а также малочисленность костей мелких видов диких животных, по сравнению с синхронными слоями раскопа 5 (табл. 1). Таким образом, объективную оценку раздробленности костей, соотношения элементов скелета и видового состава животных в данном случае нам дают только материалы 2011 г. из раскопа 5. В связи с этим при анализе состава элементов скелетов животных данные из раскопа 4 не были включены, т. к. дают искаженную картину.
Возрастная структура стада домашних животных определялась по степени стертости жевательной поверхности зубов по методике А. Грант ( Grant , 1982). Эта методика, в отличие от данных по прирастанию эпифизов трубчатых костей, позволяет довольно точно определять возраст от рождения до смерти животного. Для анализа использовались нижние челюсти и зубы с одной стороны тела. Следовательно, расчеты процентов проводились для количества особей. Структура забоя показана для объединенных выборок: XI–XIV вв. (А-Г 2007 г. и Г верх 2011 г.) и XV–XVII вв. (все остальные горизонты).
Результаты
Собранные в ходе раскопок костные остатки в массе представляют собой т. н. кухонные отходы – фрагментированные или поврежденные кости от животных, которых употребляли в пищу («пищевые» виды) ( Цалкин , 1956).
Кости домашних животных являются наиболее многочисленными среди млекопитающих во все временные периоды (табл. 1, 3).
Остатки крупного рогатого скота доминируют во всех горизонтах, их доля увеличивается от ранних периодов (XI–XIII вв.) к более поздним (XVI–XVII вв.) (табл. 2). Среди остатков представлены все элементы скелета (рис. 1). Можно отметить, что количество остатков парных костей головы (нижних челюстей) сопоставимо с количеством остатков парных костей тела (табл. 3). Это свидетельствует о разделке туш прямо на поселении. Различий в степени раздробленности костей крупного рогатого скота по горизонтам нет. Целыми являются наименее ценные части туши – зубы, мелкие кости запястья и заплюсны, фаланги пальцев и хвостовые позвонки, а практически все крупные кости сильно раздроблены. Кости черепа, нижних челюстей и все трубчатые кости верхних частей ног были порублены на множество фрагментов. Ребра рубили на три части. Большая часть позвонков разрублена вдоль тела позвонка на две-три части. У тазовых костей сохранились в основном подвздошная кость и суставная впадина. Вероятно, при разрубании суставная часть тазовой кости отделялась вместе с бедром, а подвздошная кость оставалась в туловищном отделе. Некоторые кости коровы были обработаны человеком (7 экз.), имели следы пребывания в огне (71 экз.), были погрызены собаками (193 экз.). Среди остатков наименьшее количество составляют рога, а преобладают кости туловищного отдела и верхних отделов ног (рис. 1). Это самые мясные части туши животного. Доля костей нижних отделов ног в два раза меньше, чем доля верхних отделов. Это связано с разной степенью их раздробленности. Доли целых костей и фрагментов у трубчатых костей верхних отделов конечностей соотносятся как 2 % : 98 %, а у нижних частей ног как 58 % : 42 %.
Таблица 2. Соотношение остатков домашних и диких видов млекопитающих, птиц и рыб из раскопок Земляного городища, %
|
Категория |
Века |
|||
|
XI–XIII |
XIV–XV |
XV–XVI |
XVI–XVII |
|
|
Крупный рогатый скот |
58 |
65 |
73 |
80 |
|
Свинья |
29 |
23 |
16 |
10 |
|
Мелкий рогатый скот |
12 |
11 |
10 |
9 |
|
Лошадь |
1 |
1 |
1 |
1 |
|
Всего |
100 |
100 |
100 |
100 |
|
Домашние млекопитающие |
79 |
89 |
93 |
95 |
|
Дикие млекопитающие |
4 |
3 |
2 |
2 |
|
Птица |
9 |
4 |
3 |
2 |
|
Рыба |
8 |
4 |
2 |
1 |
|
Всего |
100 |
100 |
100 |
100 |
|
Количество, экз. |
9447 |
1461 |
8792 |
894 |
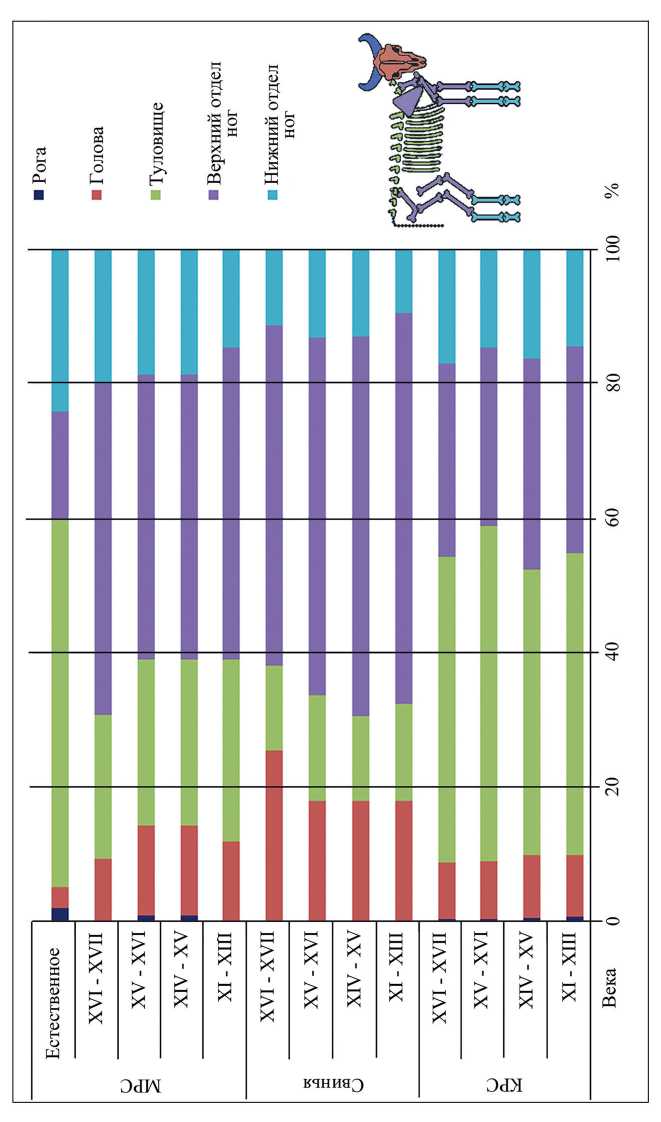
Рис. 1. Соотношение остатков различных отделов скелета домашних животных в материалах разных веков из раскопок Земляного городища и естественное соотношение в одном скелете
Таблица 3. Анатомический состав и количество остатков млекопитающих из раскопок Земляного городища в 2011 г.
|
Элемент скелета |
и Рн W |
w hQ И s И О |
Рн s |
l-Q 4 CS a |
ctf О О |
3 t§ |
hQ О |
СП |
ю о и |
ц m |
с$ S S |
С & о К |
|
Рог |
42 |
4 |
— |
— |
— |
58 |
— |
— |
— |
— |
||
|
Череп |
441 |
166 |
48 |
2 |
2 |
1 |
9 |
– |
3 |
– |
1 |
– |
|
Нижняя челюсть |
399 |
199 |
120 |
8 |
5 |
4 |
14 |
6 |
7 |
– |
– |
– |
|
Подъязычная |
29 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Изолированные зубы |
655 |
221 |
183 |
16 |
4 |
– |
20 |
2 |
24 |
1 |
– |
– |
|
Позвонки |
1137 |
95 |
141 |
9 |
5 |
6 |
4 |
4 |
3 |
1 |
3 |
– |
|
Грудина |
16 |
2 |
2 |
2 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Ребра |
3029 |
210 |
185 |
10 |
7 |
1 |
4 |
1 |
5 |
– |
– |
– |
|
Ключица |
– |
– |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
|
Лопатка |
538 |
202 |
79 |
9 |
2 |
1 |
2 |
6 |
2 |
— |
1 |
|
|
Плечевая |
517 |
221 |
134 |
9 |
4 |
6 |
9 |
8 |
13 |
– |
– |
1 |
|
Локтевая |
216 |
159 |
34 |
2 |
5 |
2 |
2 |
10 |
4 |
– |
3 |
1 |
|
Лучевая |
274 |
144 |
106 |
13 |
6 |
2 |
2 |
11 |
4 |
– |
1 |
– |
|
Таз |
427 |
156 |
72 |
7 |
1 |
4 |
– |
12 |
8 |
1 |
– |
2 |
|
Бедренная |
347 |
157 |
116 |
2 |
– |
3 |
5 |
7 |
7 |
– |
– |
– |
|
Большая берцовая |
463 |
194 |
142 |
9 |
6 |
2 |
3 |
9 |
11 |
– |
– |
1 |
|
Малая берцовая |
– |
38 |
– |
– |
– |
– |
– |
– |
2 |
– |
– |
– |
|
Коленная чашечка |
22 |
1 |
1 |
1 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Кости пясти |
147 |
– |
64 |
2 |
– |
– |
3 |
– |
– |
– |
– |
– |
|
Кости плюсны |
169 |
– |
66 |
3 |
– |
– |
4 |
– |
– |
– |
– |
– |
|
Метаподии |
57 |
82 |
19 |
7 |
12 |
– |
2 |
20 |
1 |
1 |
41 |
1 |
|
Запястье, предплюсна |
469 |
179 |
80 |
19 |
– |
1 |
4 |
– |
1 |
1 |
2 |
– |
|
Фаланги |
681 |
46 |
31 |
24 |
2 |
– |
30 |
1 |
1 |
1 |
7 |
– |
|
Скелет |
– |
– |
– |
– |
– |
4 |
– |
– |
– |
– |
– |
– |
|
Всего |
10075 |
2472 |
1627 |
154 |
61 |
37 |
175 |
97 |
97 |
6 |
59 |
6 |
Возрастной состав забитых особей показывает преобладание остатков от взрослых особей старше 3 лет во все периоды (более 55 %; рис. 2). Общая доля молодых животных до 3 лет в материалах двух периодов одинаковая. В оба периода среди особей до одного года представлены только новорожденные особи до 3 месяцев, а телята от полугода до 1,5 лет практически отсутствуют (рис. 2). В XI‒XIV вв. молодых животных забивали преимущественно в 2-летнем
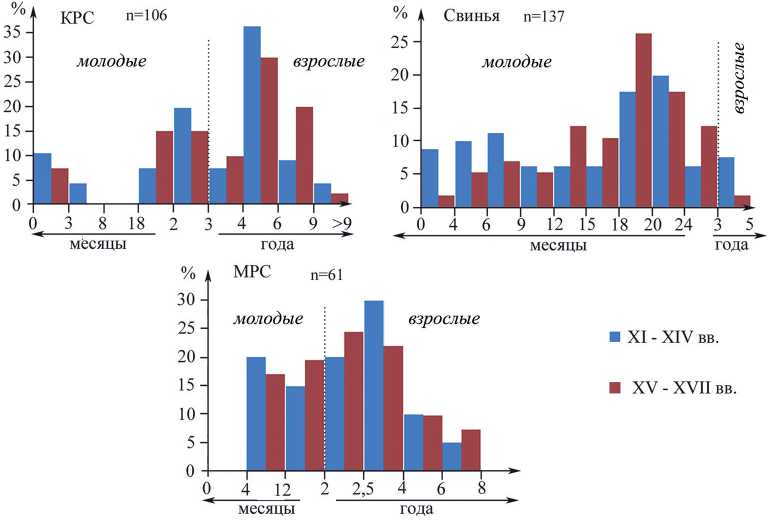
Рис. 2. Возрастная структура забитых на мясо домашних животных из раскопок Земляного городища, выявленная по степени стертости жевательной поверхности зубов нижней челюсти возрасте, в то время как в XV‒XVII вв. равномерно от 1,5 до 3 лет. Структура забоя взрослых животных в целом сходна в материалах двух периодов. Преобладают особи 4–6 лет (рис. 2), однако доля особей старше 6 лет в материалах позднего периода в два раза выше, чем в предыдущее время (рис. 2).
Свинья занимает второе место по количеству остатков в исследованной выборке. Доля костей свиньи постепенно уменьшается от ранних горизонтов к поздним (табл. 3). Характер раздробленности сходен с таковым у крупного рогатого скота. Найдено несколько костей со следами пребывания в огне (8 экз.) и обработки человеком (3 экз.). Есть кости, погрызенные собаками, а также со следами пребывания в желудочно-кишечном тракте (17 экз.). Среди остатков свиньи определены все элементы скелета, а преобладают кости верхних частей ног (табл. 3). Костей туловищного отдела почти в три раза меньше, чем у крупного рогатого скота (рис. 1). Это, вероятно, обусловлено меньшей степенью раздробленности мелких позвонков и ребер свиней по сравнению с коровой. В отличие от коров у свиней сохранилось почти в два раза больше фрагментов головы (рис. 1). Такое соотношение остатков характерно для этого вида во многих средневековых русских памятниках ( Бачура, Подопригора , 2000; Бобков-ская , 2010; Косинцев, Подопригора , 1998; Косинцев, Черная , 2000; Бачура и др ., 2020) и, возможно, связано с тем, что головы свиней чаще используют в пищу, чем у других домашних животных.
По состоянию зубов свиней получено соотношение возрастных групп, среди которых в оба периода доминируют (85–86 %) молодые животные до 2 лет (рис. 2). Остатков свиней старше пяти лет в кухонных отбросах не обнаружено. Максимальную долю в оба периода составляет группа особей в возрасте 18‒24 месяцев (рис. 2). В горизонтах XI‒XIV вв. почти в два раза выше доля поросят в возрасте до одного года, чем в более поздних горизонтах (36 % и 19 % соответственно), но при этом заметно меньше особей от года до 2 лет (50 % и 67 % соответственно).
Мелкий рогатый скот занимает третье место среди млекопитающих по количеству остатков (табл. 1; 3). Доля этого вида оставалась неизменной на протяжении всего времени. Среди остатков представлены все элементы скелета (табл. 3). Характер раздробленности сходен с таковым у крупного рогатого скота (рис. 1). На основании морфологических особенностей нижних челюстей, плечевых, лучевых, таранных, пястных и плюсневых костей, а также первых фаланг проведена видовая диагностика костей козы и овцы ( Boessneck , 1970; Zeder, Lapham , 2010; Zeder, Pilaar , 2010). Кости обоих видов были найдены во всех горизонтах, однако во все периоды доля козы не превышала 8 %.
Возрастная структура забоя овец на протяжении всего периода была одинаковая. Среди животных нет новорожденных и полугодовалых особей (рис. 2). Возрастные группы от 0,5 до 4 лет представлены в равном количестве остатков. Особей из более старших возрастных групп в 2–3 раза меньше (рис. 2). Отсутствуют животные старше 8 лет. Две челюсти козы принадлежали животным в возрасте 2,5–4 года.
Кости лошади составляют 1 % среди остатков домашних животных и представлены всеми элементами скелета (табл. 2; 3). Найдена одна фаланга от новорожденной особи, два фрагмента нижней челюсти животных, не достигших одного года, и трубчатые кости ног от полувзрослых особей (4 экз.), эпифизы у которых не приросли.
Собаке принадлежат отдельные целые кости от разных частей тела взрослых животных (табл. 3). Остатки собак немногочисленны во всех горизонтах памятника, однако их количество уменьшается от ранних слоев к поздним (табл. 1).
Почти все одиночные целые кости, а также фрагменты 4 скелетов кошек были найдены в раскопе 5 (табл. 1). Части трех скелетов найдены в межжилищном пространстве горизонта В. Фрагмент скелета задней лапы кошки был найден в юго-западной части постройки 5 горизонта Г. Все остатки принадлежали животным старше одного года, за исключением лучевой кости от молодой особи.
Промысловые млекопитающие в Ладоге представлены 11 видами (табл. 1). Общая доля их остатков крайне мала (2 %).
Среди диких животных наибольшее количество костей принадлежит лосю , однако почти 37 % всех его остатков – это обработанные фрагменты рогов, вероятно, срезанные в процессе изготовления изделий. А 73 % костей тела приходится на элементы головы и нижних частей ног (табл. 3), т. е. не мясные части туши.
Единичные кости головы кабана (два клыка и нижняя челюсть) были определены по размерам зубов (Кисагулов, 2018). Еще в начале XX в. были известны заходы кабанов в районы Финского залива и юго-западного побережья Ладожского озера (Гептнер и др., 1961).
Северному оленю принадлежали две кости ног (пяточная и первая фаланга) и фрагмент черепа с основанием рога. Рог зашлифован человеком. До второй половины XIX в. в районе Ладожского озера проходила южная граница ареала северного оленя (Там же).
Единичные находки кольчатой нерпы , скорее всего, обусловлены случайным выловом во время промысла сига и судака в Волхове, куда тюлень поднимается в конце осени ( Труханова и др ., 2012).
Все остальные виды диких млекопитающих в той или иной мере можно отнести к «пушной» добыче. Большая часть костей этих видов происходит из нижних горизонтов памятника (табл. 1). В пищу могли использовать тушки бобра, а также зайца-беляка. Остатки этих видов наиболее многочисленны и в значительной мере представлены костями тела и верхних отделов конечностей (табл. 3). Вероятно, зайцев добывали в основном ради меха. На это указывает малое количество его остатков на поселении. Кроме того, у восточных славян в XIX в. существовали определенные запреты на употребление мяса зайцев в пищу ( Зеленин , 1991. С. 147). Возможно, такие традиции существовали и в более ранние периоды.
Во всех стратиграфических горизонтах были найдены остатки лисицы , но большая часть в материалах XI–XIII вв. (табл. 1). Подавляющее большинство ее остатков – это кости нижних частей лап (метаподии и фаланги). В нижнем горизонте найдены отдельные фрагменты костей волка (табл. 3) и единичные кости ног белки. Только в материалах XI–XIII вв. (табл. 1) были найдены кости куницы (фрагмент черепа, клык, фрагмент локтевой кости и 4 метаподии). Бурый медведь представлен фрагментом черепа и 6 целыми когтевыми фалангами, найденными в различных горизонтах.
Обсуждение
Изменение соотношения костных остатков групп животных в разные хронологические периоды характеризует изменения структуры питания его жителей, которая обусловлена, в свою очередь, сменой направления хозяйства. Соотношение остатков домашних копытных в кухонных отходах типично для русских средневековых городских поселений, с доминантой в виде крупного рогатого скота. Как правило, в средневековых селах среди остатков доминирует свинья (до 50 %), а в городах ‒ крупный рогатый скот (не менее 60 %) ( Антипина , 2005). В XI‒XIII вв. свинья играла существенную роль в хозяйстве населения Ладоги наряду с крупным рогатым скотом. На протяжении рассматриваемого периода поголовье свиней постепенно сокращалось, а поголовье крупного рогатого скота, напротив, увеличивалось. Поголовье мелкого рогатого скота оставалось практически неизменным на протяжении всего времени (табл. 2).
Основу мясного потребления в Ладоге составляло мясо крупного рогатого скота. Обращает на себя внимание минимальное количество телят среди забитых животных. Известно, что восточные славяне в XIX в. редко и неохотно употребляли в пищу телятину, что было обусловлено хозяйственными соображениями.
Во-первых, ее считали малопитательной и вообще вредной. Во-вторых, коровы перестают доиться без телят, а через 2–3 года из теленка выйдет «полнорослая скотина» ( Зеленин , 1991. С. 147; Даль , 2003. С. 591). Возможно, в Ладоге телятину также практически не употребляли в пищу. Наличие новорожденных и 3-месячных телят среди забитых животных может объясняться их гибелью в первые месяцы жизни от неблагоприятных погодных условий, бескормицы и т. п. Позже окрепшие животные откармливались до достижения ими значительного веса.
Обычно коров и быков, выращиваемых специально на мясо, забивают до 3 лет. В нашем случае доля особей этой возрастной категории не превышает 40 %, а максимальную долю составляют особи репродуктивного возраста от 3 до 6 лет (рис. 2), когда коровы наиболее эффективны для получения молока. Такой возрастной состав говорит в пользу мясо-молочного направления эксплуатации крупного рогатого скота ( Антипина , 2008. С. 72). Более высокая доля животных старше 6 лет в материалах XV‒XVII вв. может свидетельствовать об увеличении времени содержания коров и, как следствие, увеличении молочной направленности эксплуатации этих домашних животных. Какое-то количество молочных коров, вероятно, содержали прямо на территории города. Основное мясное поголовье крупного рогатого скота, вероятно, разводили в сельской округе, а в город поставляли в виде туш, которые разделывали уже на месте. Среди кухонных отходов наиболее многочисленными оказались фрагменты ребер. Размеры фрагментов в большинстве случаев укладываются в 10 см. Возможно, это связано со способом приготовления грудинки крупного рогатого скота в горшках, который можно считать традиционным для русского населения в XVII в. ( Антипина , 2011).
Вторым по значению пищевым видом была свинья. Свиней обычно держат 5‒6 лет, т. к. с 7 лет свиньи теряют способность к размножению. Возрастная группа от 3 до 6 лет у свиней обычно отражает смену маточного стада ( Плеханов , 1926; Дмитриев и др ., 1989). В условиях Ладоги, возможно, эти сроки были несколько смещены ‒ от 2 до 5 лет. Эта возрастная группа представлена здесь долей в 14 % в оба периода (рис. 2). Этого количества для свиньи – вида с коротким циклом воспроизводства и высокой плодовитостью – достаточно для устойчивого воспроизводства потребляемой на мясо части ее поголовья в рамках местного разведения. Судя по большому количеству особей всех возрастов от поросят до 5-летних животных, жители Ладоги в XI–XVII вв. сами разводили свиней для собственного пропитания. Вплоть до XIV в. чаще забивали поросят первого года жизни, а позже, в XV‒XVII вв., большую часть свиней откармливали дольше, а массовый забой начинался уже со второго года жизни, после первого опороса. Скорее всего, такие различия связаны с сокращением поголовья свиней в XV‒XVII вв. (табл. 2), что привело, в свою очередь, к сокращению их забоя в первый год жизни и/или к изменениям условий содержания.
Мелкий рогатый скот занимал третье место в хозяйстве жителей Старой Ладоги. Здесь держали преимущественно овец и небольшое количество коз. Численность и тех, и других оставалась неизменной на протяжении всего рассматриваемого периода. Овец держали до 8 лет. Это максимальное время содержания этих животных и в современных хозяйствах (Дмитриев и др., 1989). Животных забивали равномерно, начиная примерно с 4-месячного возраста и до 4 лет. Затем интенсивность забоя снижалась в несколько раз. Самый высокий прирост мышечной массы наблюдается у животных до 6-месячного возраста, после чего их целесообразно забивать на мясо. Кроме того, лучшие овчины получают от молодняка 5–6-месячного возраста (Дмитриев и др., 1989). Обычно при содержании овец преимущественно для получения шерсти среди забитых животных преобладают особи старше 4 лет, доля которых должна составлять 75‒80 % (Ерохин и др., 2016). В случае Старой Ладоги доля таких особей не превышает 17 % (рис. 2). Очевидно, что овец использовали как для получения мяса, так и шкур и шерсти. Кроме того, возрастная группа этих копытных от 4 до 8 лет маркирует маточное стадо. Довольно большой процент особей данного возраста достаточен, чтобы обеспечить собственное разведение. Для получения молока содержать овец экономически невыгодно. А вот небольшое количество коз могли держать именно как дополнительный источник молока.
Лошади, вероятно, принадлежала более значительная роль в хозяйстве, чем это отражают немногочисленные костные остатки в материалах Земляного городища. Лошади, прежде всего, были ценны как транспортные животные. Половина остатков вида происходит от головы и нижних частей ног, т. е. не мясных частей тела (табл. 3), однако на трубчатых и костях осевого скелета есть следы порубов. Существовавшие запреты на потребление конины ( Зеленин , 1991. С. 147) могли нарушаться в неурожайные, голодные годы.
В Ладоге держали небольшое количество собак и кошек, о чем свидетельствует малое количество их остатков. Незначительное количество погрызенных костей (1 %) говорит о том, что собаки жили во дворах и свободно по городу не перемещались. Поскольку промысловая деятельность занимала несущественное место в жизни населения Ладоги, собак держали исключительно для охраны дворов, а не для охоты.
Промысел птиц и рыболовство в XI–XIII вв. еще играли какую-то заметную роль в хозяйстве населения Ладоги, но уже к XV в., а особенно позднее, утратили свое значение (табл. 2). Тоже можно сказать и об охоте на диких животных. Доля диких видов млекопитающих и их разнообразие неуклонно снижаются от XI в. к XVII в. (табл. 2). Горожане охотились в основном на лосей, зайцев, бобров и лисиц (табл. 1). В редких случаях добывали кабанов, северного оленя и нерпу. Максимальное количество видов диких млекопитающих было найдено в горизонтах XI–XIII вв., а в материалах XVI–XVII вв. найдены лишь единичные кости (табл. 1). Кости пушных зверей – бобра, волка, белки, куницы и бурого медведя (табл. 1) – почти исчезают уже к XV в. Это может быть связано с общим сокращением их численности в окрестностях города в результате антропогенной нагрузки (охоты, расширения пахотных земель и т. п.). Однако, судя по малочисленности остатков, на протяжении всего рассматриваемого периода охота не играла заметной роли в экономике города, а пушной промысел если и имел место, то в отдалении от города, где и производилась первичная обработка добытых животных. Оценить масштабы этой деятельности сейчас довольно трудно. А некоторые кости головы или лап диких животных могли храниться у жителей Ладоги в качестве ритуальных прикладов или охотничьих трофеев.
Заключение
Изучение сельскохозяйственных видов из остеологической коллекции Земляного городища показало сходные результаты, полученные для древнерусских городов XI–XVII вв. ( Цалкин , 1956; Антипина , 2005). Соотношение остатков домашних животных, где доминирует крупный рогатый скот, типично для городских поселений. На подворьях города для личных нужд содержали какое-то количество коров для получения молока, свиней, коз и овец, а также лошадей, кошек и собак. Большая часть говядины, скорее всего, поступала в город из близлежащих сельских поселений.
По мере развития хозяйства в период XI–XVII вв. у жителей посада Ладоги увеличивается роль домашних животных и сокращается промысловая деятельность. Начиная с конца XI в. к XVII в. происходит увеличение молочного направления в скотоводстве. Это приводит к постепенному сокращению поголовья свиней и увеличению поголовья крупного рогатого скота. Возможно, немаловажную роль в этом сыграло и увеличение роли сельского (пахотного) земледелия в экономике города (Человек и природная среда…, 2005. С. 77‒103). Все это могло способствовать сокращению потребности в мясных продуктах и частичной их замене на молоко и растительные продукты. Развитие сельского хозяйства и сведение лесов в окрестностях Ладоги могли послужить причиной значительного сокращения местообитаний диких животных, что, в свою очередь, привело к сокращению промысловой деятельности населения.
