СО2-газообмен растений гороха посевного различных морфотипов под действием янтарной кислоты и эпина при ранней корневой засухе

Автор: Клочкова Н.М., Аканов Э.Н., Третьяков Н.Н.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Регуляторы роста растений
Статья в выпуске: 1 т.39, 2004 года.
Бесплатный доступ
В лабораторных и вегетационных опытах изучали влияние янтарной кислоты, эпина и смеси этих веществ на рост, СО2-газообмен, а также фотосинтетическую и дыхательную активность растений гороха двух морфотипов при дефиците влаги. Рассматривается возможность использования соотношения между интенсивностью фотосинтеза и дыхания для оценки адаптивного потенциала растений (устойчивость к засухе).
Короткий адрес: https://sciup.org/142132951
IDR: 142132951 | УДК: 636.656:581.1:58.03
CO2-gas exchange in Pisum sativum of different morphotypes during early root drought under the influence of succinic acid and
In laboratory and vegetation experiments the authors studied the influence of succinic acid, epin and its mixture on the growth, CO2-gas exchange and also photosynthetic and respiratory activity in Pisum sativum of two morphotypes (tendril and leaflet) during moisture deficiency. The possibility of usage of the correlation between photosynthesis and respiration activities for estimation of the plants adaptive potential (tolerance to drought). It was shown that succinic acid is the most effective compound positive affecting on CO2-gas exchange and productivity of pea plants as under optimal conditions as in the moisture deficiency. The pea plants of leaflet-type are more tolerant to early root drought in comparison with plants of tendril-type.
Текст научной статьи СО2-газообмен растений гороха посевного различных морфотипов под действием янтарной кислоты и эпина при ранней корневой засухе
В последние годы в практике растениеводства широкое распространение получили синтетические регуляторы роста растений, в том числе стероиды (1). Однако до сих пор не раскрыт механизм физиологического действия этих соединений (2). Как известно, при стрессовых факторах в тканях растений образуются специфические вещества, наиболее активным компонентом которых является янтарная кислота. Считается, что янтарная кислота не представляет опасности для человека и окружающей среды, так как относится к «биогенным» стимуляторам роста (3). Имеются сведения об аддитивном действии некоторых регуляторов роста на процессы фотосинтеза и дыхания (4).
Изучение защитно-приспособительных реакций фотосинтетического аппарата растений под воздействием неблагоприятных факторов среды — одна из центральных задач физиологии растений (5). Актуальной остается и проблема разработки физиологических экспресс-методов для отбора высокоурожайных сортов. Сведения об устойчивости баланса между энергетическими затратами на защитнокомпенсаторные реакции (дыхание) и способностью растений восполнять их в процессе фотосинтеза могут быть использованы для определения адаптивного потенциала растений (6).
Недостаточная засухоустойчивость растений гороха ограничивает возможность получения стабильных урожаев. При влажности почвы ниже 50 % ППВ нарушается водный обмен, резко снижается интенсивность фотосинтеза и дыхания растений (7). У возделываемых в настоящее время сортов гороха листочкового типа число и площадь листьев близки к оптимальным, поэтому дальнейшие исследования, очевидно, должны быть направлены на увеличение фотосинтетической активности не только листьев, но и других органов растений (8).
В связи с этим целью нашей работы было исследование влияния антистрессовых адаптогенов (янтарная кислота, эпин и смесь этих веществ) на газообмен и фотосинтетическую активность растений гороха посевного различных морфотипов в условиях водного дефицита, а также возможности использования соотношения между этими показателями для оценки засухоустойчивости.
Методика. Объектом исследований служили растения гороха посевного двух морфотипов — усатого (сорт Норд, незасухоустойчивый) и листочкового (сорт Ор-ловчанин, засухоустойчивый) (селекция ВНИИ зернобобовых культур, Орловская обл.). Семена замачивали в растворах янтарной кислоты, эпина и смеси янтарная кислота + эпин в концентрации соответственно 0,1 г/л, 10-15 М и 0,05 г/л + 10-15 М в течение 2 ч; в контроле использовали водопроводную воду. Затем семена раскладывали по чашкам Петри (по 15 шт. в каждую) и в течение 8 сут проращивали в растворе Кнопа (2-кратное разведение) в вегетационной камере при освещенности 18 клк, температуре 23-25 оС, влажности воздуха 60-70 % и фотопериоде 16 ч. Через 8 сут одну часть чашек Петри с растениями использовали для проведения лабораторного, другую — вегетационного опытов.
В лабораторном опыте (3-кратная повторность) оценивали газообмен растений (на свету и в темноте) непосредственно во время действия корневой засухи и в последействии. Кратковременную засуху (3 сут) создавали посредством замены питательного раствора на раствор осмотика ПЭГ-6000 (10 атм.). При этом растения в контроле распределяли по двум вариантам: К — раствор ПЭГ-6000; АК (абсолютный контроль) — водопроводная вода. Через 3 сут во всех вариантах опыта и контроля осмотические растворы заменяли на исходный питательный раствор. СО 2 -газообмен регистрировали ежесуточно с 6- до 19-суточного возраста растений. Газообмен всего микроценоза оценивали, помещая поочередно чашки Петри в прозрачную герметичную камеру, соединенную по «замкнутому» контуру с ИК-газоанализатором ГОА-4 (концентрация СО2 0-0,05 %), что позволяло снизить влияние индивидуальных особенностей растений на показатели и учесть действие ценотического фактора на адаптивные реакции. При определении интенсивности фотосинтеза и дыхания в камере поддерживали те же параметры, что и в период подращивания растений; экспозиция составляла около 20 мин — 10 мин на свету и 10 мин в темноте.
В вегетационном опыте 8-суточные растения из чашек Петри пересаживали в сосуды емкостью 7 л с дерново-подзолистой тяжелосуглинистой почвой (по 9 шт. в каждый). Использовали те же варианты, что и в лабораторном опыте; повторность 3-кратная. Для создания водного дефицита прекращали полив растений. В течение 10 сут влажность почвы поддерживали на уровне 30 % ППВ, на 11-е сут — поднимали до 70 % ППВ и оставляли на этом уровне до конца опыта. СО2-газообмен оценивали на следующих фазах: семи-девяти листьев, ветвления, бутонизации, цветения и молочной спелости. О водообмене растений судили по интенсивности транспирации и водоудерживающей способности (9).
Результаты. При оценке в условиях вегетационного опыта высоты растений гороха листочкового и усатого морфотипов выявлено преимущество первых (в среднем на 3,3-3,5 см) (табл.). В оптимальных условиях высота растений сортов Норд и Орловчанин, начиная с фазы пяти-семи и до семи-девяти настоящих листьев увеличивалась на 7,7 см, а при засухе (фаза ветвления) — соответственно на 4,6 и 4,8 см. Максимальной высотой характеризовались растения гороха усатого морфотипа в варианте с обработкой семян янтарной кислотой. Высота растений листочкового морфотипа по вариантам опыта существенно не различалась. Аналогичные закономерности отмечены и в условиях лабораторного опыта.
По скорости образования очередных листьев не отмечено существенных различий как между растениями разных морфотипов, так и по вариантам обработки семян. Следует отметить, что растения листочкового морфотипа обладали более развитой корневой системой по сравнению с таковыми усатого морфотипа. В оптимальных условиях у растений обоих сортов после обработки семян янтарной кислотой не выявлено существенных преимуществ, однако при засухе у них продолжался рост корней, в то время как в контроле последний резко замедлялся. Так, в течение 4 сут длина корней у растений, полученных из семян, обработанных янтарной кислотой, увеличивалась на 1,33,0 см, а в других вариантах опыта — на 0,3-1,0 см.
Высота (см) растений гороха сортов Орловчанин и Норд в зависимости от воздействия различных физиологически активных веществ в оптимальных условиях и при дефиците влаги (вегетационный опыт)
|
Вариант опыта |
Фаза развития растений |
||
|
пять-семь настоящих листьев |
семь-девять настоящих листьев |
ветвление |
|
|
70 % ППВ |
30 % ППВ |
||
|
С о р т О р л о в ч а н и н (листочковый морфотип) Янтарная кислота 19,7 26,7 31,0 Эпин 19,7 27,0 32,4 Янтарная кислота + эпин 18,0 27,0 32,1 Контроль 18,7 27,3 32,0 Абсолютный контроль 17,0 23,6 28,0 Среднее 18,6 26,3 31,1 С о р т Н о р д (усатый морфотип) Янтарная кислота 16,2 24,1 28,9 Эпин 15,0 22,9 27,5 |
|||
Янтарная кислота + эпин14,9
22,5
22,0
23,7
23,0
27,0
26,4
28,4
27,6
Контроль15,0
Абсолютный контроль15,4
Среднее15,3
П р и м е ч а н и е. Описание вариантов опыта см. в разделе «Методика».
У растений листочкового морфотипа под воздействием засухи в условиях лабораторного опыта снижалась скорость образования сухого вещества; после обработки семян янтарной кислотой характер этого процесса соответствовал таковому при оптимальных условиях. У растений сорта Орловчанин в условиях вегетационного опыта отмечена б о льшая интенсивность транспирации, чем у таковых сорта Норд. Максимальная интенсивность транспирации у растений обоих морфотипов наблюдалась при оптимальных условиях водообеспечения. В условиях засухи интенсивность транспирации наиболее существенно снижалась в случае обработки семян эпином — до 120 мг/г сырой массы ⋅ ч-1, что, возможно, связано с максимальным закрытием устьиц или увеличением содержания в клетках коллоидно-связанной воды.
Листья растений гороха листочкового морфотипа характеризовались наибольшей водоудерживающей способностью. У растений усатого морфотипа при обработке семян янтарной кислотой водоудерживающая способность составляла 360 мг/г сырой массы ⋅ ч-1, листочкового морфотипа — 330 (под воздействием эпина) и 350 (контроль) мг/г сырой массы ⋅ ч-1. Увеличение водоудерживающей способности обусловлено более высоким содержанием в клетках осмотически активных веществ и свидетельствует об адаптации растений к недостатку влаги. По-видимому, при обработке семян янтарной кислотой и эпином увеличивается также количество коллоидно-связанной воды.
Нами отмечены значительные различия по составляющим СО 2 -газообмена растений в разных вариантах опыта. По данным литературы, минимальное снижение соотношения между интенсивностью фотосинтеза и дыхания у устойчивых сортов гороха под воздействием стрессоров обусловливает более стабильные физиологические процессы (10).
В условиях лабораторного опыта в течение первых 12 сут у проростков гороха во всех вариантах опыта не отмечено видимого фотосинтеза. В этот период рост растений осуществлялся за счет метаболитов, поступающих из семядолей. Поэтому суммарный газообмен отражал интенсивность дыхания. Выявлены различия по газообмену при оптимальных условиях, действии засухи и репарации. В оптимальных условиях в начальный период развития растений наблюдалось постепенное увеличение интенсивности фотосинтеза (рис. А).
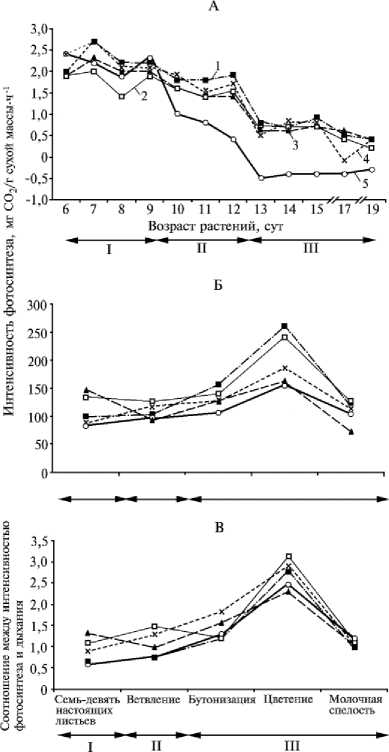
Интенсивность фотосинтеза и соотношение между интенсивностью фотосинтеза и дыхания у растений гороха посевного сорта Орловчанин (листочковый морфотип) по фазам онтогенеза в условиях засухи под влиянием различных физиологически активных веществ : А — лабораторный, Б и В — вегетационный опыты; I — до засухи, II — период засухи, III — период репарации; 1, 2 и 3 — обработка семян соответственно эпином, янтарной кислотой и смесью эпин + янтарная кислота, 4 и 5 — соответственно контроль (водный раствор ПЭГ-6000, 10 атм.) и абсолютный контроль (водопроводная вода).
При обработке различными физиологически активными веществами (ФАВ) интенсивность газообмена существенно не менялась, а фотосинтеза — возрастала под воздействием янтарной кислоты до 1,7 мг СО 2 /г сухой массы ⋅ ч-1. В условиях засухи интенсивность фотосинтеза снижалась, особенно при обработке семян эпином. В этот период различия по вариантам обработки были более существенными, в период репарации они вновь нивелировались. При засухе интенсивность фотосинтеза растений возрастала медленнее, чем при оптимальной водообеспе-ченности. Интенсивность дыхания в начале засухи резко увеличивалась — в среднем с 2,5 до 4,0 мг СО 2 /г сухой массы ⋅ ч-1, а затем плавно снижалась. Наиболее стабильным этот показатель был у растений, полученных из семян, обработанных янтарной кислотой.
В условиях вегетационного опыта мы оценивали динамику газообмена микроценоза в зависимости от возраста растений. Интенсивность фотосинтеза у растений сорта Орловчанин с возрастом постепенно возрастала, а затем плавно снижалась (см. рис. Б). У растений сорта Норд этот показатель резко возрастал в фазе семь-девять настоящих листьев—ветвление, а в фазе бутонизация—молочная спелость также резко снижался.
У растений листочкового мор-фотипа интенсивность фотосинтеза под воздействием ФАВ увеличивалась, усатого морфотипа — уменьшалась. Интенсивность темнового дыхания растений сорта Орловчанин к концу периода вегетации увеличивалась; у растений сорта Норд наблюдалась обратная закономерность. После обработки семян эпином у растений листочкового мор- фотипа интенсивность дыхания повышалась до 140 мг СО2/(раст⋅ч), при засухе, но без обработки регуляторами роста — снижалась до 70 мг СО2/(раст⋅ч). В период засухи интенсивность дыхания растений в варианте обработки семян янтарной кислотой резко увеличивалась с 80 до 110 мг СО2/(раст⋅ч), а в последействии (фаза бутонизация—цветение) — снижалась; к концу периода вегетации — возрастала до 100 мг СО2/(раст⋅ч). Следует отметить, что на начальных стадиях развития интенсивность дыхания растений усатого морфотипа в контроле была меньше, чем при обработке семян ФАВ — соответственно 40 и 70 мг СО2/(раст⋅ч), однако в период бутонизации резко возрастала до 140 мг СО2/(раст⋅ч). Под влиянием регуляторов роста этот показатель увеличивался при оптимальных условиях.
Максимальное соотношение между интенсивностью фотосинтеза и дыхания наблюдалось в период цветения, что свидетельствует об эффективном использовании растениями фотоассимилятов (см. рис. В). В фазе семь-девять настоящих листьев— ветвление соотношение между интенсивностью фотосинтеза и дыхания у растений усатого морфотипа было выше, чем у таковых листочкового морфотипа, а на более поздних фазах (бутонизация—молочно-восковая спелость) — наоборот. У растений сорта Орловчанин это соотношение плавно возрастало до фазы цветения, а затем снижалось; сорта Норд — резко повышалось в период семь-девять настоящих листьев—ветвление, а затем снижалось. У растений сорта Орловчанин под воздействием ФАВ этот показатель в фазе ветвление—бутонизация (период засухи) увеличивался во всех вариантах опыта и был наибольшим (3,2) под воздействием янтарной кислоты, наименьшим (2,4) — при обработке семян смесью янтарная кислота + эпин. У растений усатого морфотипа различия между вариантами при обработке семян ФАВ были более существенными, чем у таковых листочкового морфотипа, что свидетельствует об их меньшей устойчивости. Максимальное соотношение между интенсивностью фотосинтеза и дыхания (1,6) также отмечено при обработке семян янтарной кислотой на более поздних этапах вегетации растений. Начиная с фазы семь-девять настоящих листьев—ветвление до фазы бутонизации, этот показатель увеличивался, а ближе к фазе молочной спелости — постепенно снижался. Аналогичная закономерность выявлена в варианте янтарная кислота + эпин, но соотношение в этом случае было наименьшим. У растений, полученных из семян, не обработанных ФАВ, в условиях засухи отмечено резкое снижение этого показателя, которое сохранялось на протяжении всего вегетационного периода.
Следовательно, у более устойчивых растений листочкового морфотипа газообмен нормализуется на более ранних стадиях развития, соотношение между интенсивностью фотосинтеза и дыхания снижается в меньшей степени, что, возможно, позволяет им оптимизировать энергоресурсы для обеспечения метаболических процессов в отличие от менее устойчивых растений.
Общая зерновая продуктивность микроценозов была обусловлена особенностями СО 2 -газообмена растений разных морфотипов в различных вариантах опыта. Наибольшей продуктивностью характеризовались растения в варианте с янтарной кислотой (на 30 % выше, чем в контроле).
Таким образом, растения гороха посевного листочкового морфотипа (сорт Орловчанин) характеризуются минимальной амплитудой изменчивости по соотношению между интенсивностью фотосинтеза и дыхания при действии ранней корневой засухи. Показано, что обработка семян гороха янтарной кислотой оказывает положительное влияние на СО 2 -газообмен и продуктивность растений как при нормальном увлажнении, так и при дефиците влаги, особенно у сорта Орловчанин (лис-точковый морфотип). Сопоставление характера СО 2 -газообмена в условиях лабораторного (до 19-суточного возраста) и вегетационного (до фазы молочной спелости) опытов свидетельствует о возможности диагностики устойчивости растений гороха посевного к засухе в возрасте 6-7 сут на средах с ПЭГ-6000 при использовании вышеприведенных методических подходов.
Л И Т Е Р А Т У Р А
-
1. Т р е т ь я к о в Н.Н., Я к о в л е в А.Ф., Г о р я ч е в а О.В и др. Влияние эпибрассинолида на интенсивность фотосинтеза и продуктивность яровой пшеницы в условиях засухи. Мат. Междунар. на-уч.-метод. конф. «Продукционный процесс сельскохозяйственных культур». Орел, 2001, 1.
-
2. Е р ш о в а А.Н., В и н о к у р о в а Н.В. Действие кинетина и эпибрассинолида на содержание гликозида, свободных аминокислот, белка и семенную продуктивность растений гороха. Тез. докл. V Междунар. конф. «Регуляторы роста и развития растений». М., 1999: 91.
-
3. К о ф Э.М., Б о р и с о в а Т.А. Антистрессовое действие янтарной кислоты на проростки гороха. Агрохимия, 1999, 1: 55-59.
-
4. П у з и н а Т.И., К о р а б л е в а Н.П., К и р и л л о в а И.Г. и др. Действие синтетических регуляторов роста на некоторые физиологические показатели и продуктивность растений картофеля. Мат. Междунар. науч.-метод. конф. «Продукционный процесс сельскохозяйственных культур». Орел, 2001, 1.
-
5. У д о в е н к о Г.В. Физиолого-генетические аспекты адаптации растений к экстремальным условиям среды. В сб.: Физиолого-биохимические механизмы регуляции адаптивных реакций растений и агрофитоценозов. Кишинев, 1984: 118.
-
6. Р а х м а н к у л о в а З.Ф. Соотношение фотосинтеза и дыхания как энергетическая основа адаптации растений к неблагоприятным внешним условиям. Автореф. докт. дис. М., 2002.
-
7. Б а х т е н к о Е.Ю. Аутэкологический подход к физиологическому ответу растений на затопление и засуху. Автореф. докт. дис. М., 2001.
-
8. А м е л и н А.В. Морфофизиологические основы повышения эффективности селекции гороха. Авто-реф. докт. дис. М., 2001.
-
9. Г у с е в М.В. Малый практикум по физиологии растений. М., 1982.
-
10. Н о в и к о в а Н.Е. Физиологическое обоснование роли морфотипа растений в формировании урожайности сортов гороха. Автореф. докт. дис. Орел, 2002.
Московская сельскохозяйственная академия им. К.А. Тимирязева, 127550, Москва, ул. Тимирязевская, 49
