Ассоциация аллельных вариантов гена лептина с показателями молочной продуктивности коров костромской, ярославской и черно-пестрой пород в Костромской области

Автор: Чаицкая К.Д., Лемякин А.Д., Тяжченко А.Н., Чаицкий А.А., Щеголев П.О., Никифорова А.И., Кофиади И.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика
Статья в выпуске: 6 т.59, 2024 года.
Бесплатный доступ
Ген лептина - один из ценных генетических маркеров, определяющих молочную продуктивность, технологические свойства молока и продолжительность хозяйственного использования крупного рогатого скота, что имеет практическое значение в селекционно-племенной работе. Ведение маркер-ориентированной селекции животных по конкретным показателям продуктивности и функционального долголетия с использованием ассоциированных с ними генотипов по гену лептина способствует сохранению и совершенствованию ценных племенных ресурсов отечественных пород, а также созданию референсных стад крупного рогатого скота. В настоящей работе впервые получены данные о наличии достоверной ассоциации локуса гена лептина LEP-A80V с хозяйственно полезными признаками у коров черно-пестрой породы на территории Костромской области, что доказывают значимость этих генотипов в характеристике племенных и продуктивных качеств животных региона. Цель работы заключалась в изучении ассоциативных связей различных аллельных вариантов гена лептина с показателями молочной продуктивности у коров из племенных хозяйств Костромской области. Объектом исследований был крупный рогатый скот (Bos taurus) трех пород (Костромская обл., 2019-2021 годы), всего 141 корова, в том числе костромской породы - 33, ярославской - 47, черно-пестрой - 61 животное. В исследования использовали данные племенного и зоотехнического учета. Оценка молочной продуктивности проводилась по результатам ежемесячных контрольных доек. Для выделения ДНК использовали цельную кровь. Биоматериал отбирали из хвостовой вены в вакуумные пробирки с антикоагулянтом EDTA. Выделение ДНК осуществляли с помощью набора DNeasy Blood & Tissue Kit («QIAGEN N.V.»», Германия). Полиморфные варианты гена лептина LEP определяли методом полимеразой цепной реакции в реальном времени (ПЦР-РВ) по трем локусам: LEP-R25C (аллели R, C; генотипы RR, RC, CC), LEP-A80V (аллели A, V ; генотипы AA, AV, VV) и LEP-Y7F (аллели Y , F ; генотипы YY, YF, FF) с использованием амплификатора DTprime (ООО «НПО ДНК-технология», Россия) и технологии HRM (high resolution melting). Установлено, что генное равновесие по локусу LEP-A80V у коров черно-пестрой породы было значимо смещено в сторону гомозиготных животных, преимущественно с генотипом АА (0,541±0,064), при этом полиморфизм оказывал статистически значимое влияние на удой и содержание жира в молоке, причем повышенным удоем обладали гомозиготные носители аллеля A (8856 кг с доверительным интервалом 8819,2-9394 кг), а наибольшей жирностью молока - гетерозиготные животные (4,05 % с доверительным интервалом 3,99-4,21 %). Генное равновесие по локусу LEP-Y7F у животных всех изученных групп оказалось достоверно смещено в сторону гомозигот YY (0,936-0,970), однако влияние этого полиморфизма на молочную продуктивность коров не обнаружили. Тем не менее для полиморфизма LEP-Y7F выявлена тенденция к большему удою и содержанию жира в молоке у первотелок черно-пестрой породы с генотипом YY (соответственно на 2275 кг и 0,075 %). По локусу LEP-R25C достоверное смещение генного равновесия наблюдалось только у коров черно-пестрой породы в сторону гомозигот, но статистически значимого влияния этого полиморфизма на молочную продуктивность мы не обнаружили, вероятно, в силу недостаточной численности генотипированных животных, что указывает на необходимость дальнейших исследований на более многочисленном поголовье.
Полиморфные варианты, молочная продуктивность, крупный рогатый скот, костромская порода, ярославская порода, черно-пестрая порода
Короткий адрес: https://sciup.org/142244123
IDR: 142244123 | УДК: 636.082.12:575.174.015.3 | DOI: 10.15389/agrobiology.2024.6.1091rus
Association of the leptin gene allelic variants with dairy productivity parameters in cows of Kostroma, Yaroslavl and Black-and-White breeds of the Kostroma region
The leptin gene is one of the most valuable genetic markers that determine milk productivity, technological features of milk production and productive longevity of cattle, which is of practical importance in breeding work. Conducting marker-associated breeding based on specific signs of productivity and functional longevity of animals using related leptin genotypes contributes to the preservation and multiplication of valuable breeding resources of local cattle breeds, as well as the creation of reference cattle herds. In this work, for the first time, data were obtained on the reliable association of the leptin gene LEP-A80V locus of with economically useful traits in Russian black-and-white cows in the Kostroma region, which proves the importance of these genotypes for characterizing the breeding and productive qualities of livestock in this area. The aim of the work was to study the associative relationships of various allelic variants of the leptin gene with signs of dairy productivity of cows from breeding farms in the Kostroma region. Kostroma, Russian black-and-white and Yaroslavl cows ( Bos taurus ) from breeding farms of the Kostroma region (2019-2021), a total of 141 cows, including 33, 47 and 61 animals of the corresponding breeds were tested. The data of breeding and zootechnical records were analyzed. Milk productivity was evaluated based on monthly control milking. Whole blood for DNA isolation was taken from the caudal vein into vacuum tubes with the anticoagulant EDTA. DNA was isolated using the DNeasy blood and tissue analysis kit (QIAGEN N.V, Germany). Polymorphic variants of the leptin LEP gene were determined by real-time polymerase chain reaction (RT-PCR) at three loci, the LEP-R25C (alleles R , C ; genotypes RR , RC , CC ), LEP-A80V (alleles A , V ; genotypes AA , AV , VV ), and LEP-Y7F (alleles Y , F , genotypes YY , YF , FF ) using a DTprime amplifier (DNA Technologies LLC, Russia) and HRM (high resolution melting) technology. It was found that the allele balance at the LEP-A80V locus in Russian black-and-white cows significantly shifts towards homozygous animals, mainly of the AA genotype (0.541±0.064). Homozygous carriers of the A allele have increased milk yield of 8856 kg (a confidence interval of 8819.2-9394 kg), while heterozygous animals had the highest milk fat content of 4.05 % (a confidence interval of 3.99-4.21 %). The allele balance at the LEP-Y7F locus in all the studied groups turned out to be significantly biased to YY homozygotes (0.936-0.970), however, effect of this polymorphism on dairy traits was not revealed. Nevertheless, for Y7F polymorphism, a tendency was revealed towards increased milk yield and fat content in the Russian black-and-white first-calf cows of the YY genotype, by 2275 kg and 0.075 %, respectively. For the LEP-R25C locus, a significant shift in the allele balance occurs only in Russian black-and-white cows towards homozygosity, but no statistically significant effect of this polymorphism on milk productivity was found, probably due to an insufficient number of genotyped animals, which indicates necessity of further research at a larger population.
Текст научной статьи Ассоциация аллельных вариантов гена лептина с показателями молочной продуктивности коров костромской, ярославской и черно-пестрой пород в Костромской области
Генотипирование по ДНК-маркерам все чаще применяется в практике скотоводства для оценки племенной ценности животных в раннем возрасте. Также это позволит использовать маркер-ориентированную селекцию в стадах крупного рогатого скота (КРС) одновременно по всем интересующим хозяйственно полезным признакам (1). В настоящее время ведется поиск генов, связанных не только с высокой молочной продуктивностью, но и с сохранением функционального долголетия и здоровья коров, успешной реализацией их генетического потенциала, а также с высоким качеством молочной продукции.
Продукт экспрессии гена лептина ( LEP ) — гормон лептин — вырабатывается адипоцитами (клетками жировой ткани) и играет ключевую роль в энергетическом обмене. По механизму действия лептин относят к нейротропным гормонам. Повышение концентрации лептина в плазме крови подавляет аппетит, стимулирует энергетический обмен, активирует процесс утилизации жиров в жировой ткани и снижение живой массы. Лептин также воздействует на центр теплопродукции, участвует в регуляции функции щитовидной железы и влияет на функционирование иммунной системы (2). От его концентрации в плазме крови КРС зависит энергетический баланс организма, рост жировой ткани, а также потребление кормов (3). Накопление в крови телок достаточного количества лептина необходимо для полового созревания (4, 5), оно увеличивает секрецию гонадотропных гормонов (6), а также способствует развитию молочной железы (7). Есть мнение, что полиморфизм гена LEP существенно влияет на количественный и качественный состав молока и предположительно ассоциирован с функциональным долголетием коров, что имеет большую практическую значимость в селекционно-племенной работе (8).
Учитывая высокую полиморфность гена лептина и разнообразие физиологических функций, в которых он принимает участие, исследователи часто рассматривают его в качестве самостоятельного ДНК-маркера, способного оказывать существенное влияние на рост, развитие, репродуктивные функции и молочную продуктивность сельскохозяйственных животных (9).
Анализируя научные публикации, в которых фигурирует ген лептина, нельзя не отметить, что волна интереса к нему как к ДНК-маркеру молочной и мясной продуктивности пришлась на первое десятилетие XXI века. Затем внимание к этому маркеру заметно ослабло, и в качестве более перспективных генов-кандидатов стали рассматривать другие гены, например LEPR, DGAT1, TG . На наш взгляд, падение интереса к гену LEP связано с распространенной в ряде стран концепцией максимально интенсивного использования коров в течении 2-3 лактаций с последующим выбытием. Подобная концепция молочного скотоводства заставляет исследователей отдавать предпочтение ДНК-маркерам, обеспечивающим максимальную молочную продуктивность в течении короткого срока хозяйственного использования. Воздействие полиморфизма гена лептина на продуктивность молочного скота оценивается как слабое и несистемное, поэтому не вписывается в концепцию «одноразовой коровы». Тем не менее влияние полиморфизма гена лептина на продуктивное долголетие и пожизненную продуктивность коров обнаружено, что привлекает внимание как ученых из стран Латинской Америки, Ближнего Востока и ЮгоВосточной Азии (10-12), так и российских исследователей, придерживающихся концепции долговременного хозяйственного использования молочного скота (13).
У КРС ген лептина локализован в 4-й хромосоме и состоит из промоторной области, трех экзонов, двух интронов и 3´UTR-области (9, 14, 15). Ген лептина высокополиморфный. На сегодняшний день в базе данных Ensembl зарегистрировано 1385 однонуклеотидных полиморфизмов (SNPs) гена LEP, причем большая часть расположена в некодирующих областях гена, поэтому изучение их влияния на фенотип животных сопряжено с определенными трудностями. Более доступны для изучения однонуклеотидные 1092
полиморфизмы, расположенные в экзонах 2 и 3, поскольку изменения кодирующей ДНК напрямую влияют на структуру и функциональные свойства синтезируемого белка. Среди этих SNPs наибольший интерес вызывают полиморфизмы Y7F и A80V, локализованные в экзоне 2, и R25C, расположенный в экзоне 3.
Полиморфизм Y7F, хоть и находится в кодирующей части гена LEP , не пользуется популярностью ввиду того, что генотипы, отличные от YY , встречаются в популяциях крайне редко. А.В. Перчун с соавт. (16) приводят распределение аллелей Y : F для популяции коров костромской и бурой швицкой пород: соответственно 0,968:0,032 и 0,915:0,085. По данным Н.В. Ковалюк с соавт. (17), соотношение частот аллелей Y : F в голштинизи-рованной популяции коров черно-пестрой породы составляло 0,97:0,03.
L. Giblin с соавт. (18) удалось показать, что полиморфизм гена лептина Y7F у КРС ассоциирован с накоплением энергии в виде отложений жировой ткани. Вместе с тем ирландскими учеными было установлено, что аллель F SNP Y7F ассоциирован со снижением удоя и выхода молочного белка (18). А.А. Шевцова с соавт. (19) также связывают полиморфизм Y7F со снижением величины удоя коров.
Данные о распределении генотипов и аллелей полиморфизма LEP A80V для кашмирской популяции коров джерсейской породы и для местного помесного скота были получены S.A. Bhat с соавт. (20), причем для чистопородных животных соотношение аллелей A : V составляло 0,775:0,225, а среди помесей — 0,829:0,171. В свою очередь, M.R. Dar с соавт. (21) установили соотношение аллелей A и V 0,9:0,1 у помесных коров, полученных от скрещивания Bos taurus и B. indicus . Среди отечественных ученых актуальные данные по распределению генотипов и аллелей полиморфизма A80V представили Л.Н. Чижова с соавт. (22), причем для коров ярославской породы соотношение аллелей A : V составило 0,78:0,22, а для животных черно-пестрой и голштинской пород (в обоих случаях) — 0,86:0,14.
Ряд зарубежных авторов отмечают ассоциацию полиморфизма A80V гена лептина с удоем, длиной тела и возрастом первого плодотворного осеменения (23-25). В более поздних исследованиях M.A. Ca n izares-Mar-t i nez с соавт. (26) получили сходные результаты: телки голштинской породы с генотипом ÀÀ оплодотворялись и телились значительно раньше сверстниц, а также имели более короткий сервис-период. На ассоциацию этого полиморфизма с удоем также указывают M.R. Dar с соавт. (21).
В исследовании Ф.Ф. Зиннатовой с соавт. (27) установлено, что коровы с гомозиготным генотипом AA статистически значимо отличались от сверстниц с генотипом VV более высоким удоем, а у гетерозиготных животных было выявлено наибольшее содержание жира в молоке, и по выходу молочного белка они превосходили сверстниц с генотипом VV . Похожие результаты описаны в других публикациях, хотя при этом наибольшее содержание белка в молоке отмечалось у гетерозигот AV (28-30). Противоположные данные получили S.A. Bhat с соавт. (20): полиморфизм A80V не оказал статистически значимого влияния на удой и содержание белка, но существенно повлиял на жиромолочность коров, при этом носительницы генотипа ÀÀ по этому показателю значительно превосходили гетерозиготных сверстниц.
Распределение генотипов и аллелей локуса LEP-R25C рассматривалось в работе M. Ahani Azari с соавт. (31), была определена частота аллеля R на уровне 0,4. При этом сводные данные о распределении генотипов и аллелей по локусу LEP-R25C, представленные в работе Н.Ю. Сафиной (32), показывают, что в большинстве исследованных популяций КРС частота встречаемости аллеля C существенно выше, чем аллеля R, и для распространенных пород молочного скота (голштинская, черно-пестрая и айршир-ская) в среднем соотношение аллелей R:C составляет 0,42:0,58. При этом Л.Н. Чижова с соавт. (22) сообщают о соотношении аллелей R:C 0,16:0,84.
Отмечено существенное влияние полиморфизма R25C на некоторые морфологические особенности молочного скота (глубина тела, ширина таза и строение задних конечностей), а также на величину удоя коров (25). В исследованиях R.C. Chebel с соавт. (33) наблюдалась тенденция к увеличению количества получаемого молока и показателей его качественного состава у генотипов RC и CC по локусу LEP-R25C . Так, коровы голштинской породы, гомозиготные по аллелю R , были менее продуктивными (12248 кг), чем гетерозиготные (12530 кг) и гомозиготные сверстницы CC (12304 кг). При этом наименее жирным оказалось молоко коров RR (3,45 %) по сравнению с коровами RC (3,47 %) и CC (3,51 %). По белковомолочности существенной разницы выявлено не было, но наивысшие показатели при этом были зафиксированы у животных с генотипом RC (2,94 %) и RR (2,93 %) (33). В работе Ф.Ф. Зиннатова с соавт. (34) самым высоким удоем выделялись коровы черно-пестрой породы с генотипом RR , но сверстницы с генотипом CC характеризовались наибольшим содержанием жира и белка в молоке. По данным А.А. Ярыш-кина с соавт. (35), носительницы генотипа RR имели более высокий удой и содержание жира в молоке, однако по продолжительности хозяйственного использования они (как и гомозиготы CC ) статистически значимо уступали гетерозиготным сверстницам.
Однонуклеотидные полиморфизмы гена лептина активно исследуются как маркеры мясной продуктивности КРС. Так, F. Kawaguchi с со-авт. (36) выявили ассоциацию полиморфизма Y7F с признаками развития скелетной мускулатуры у черного японского скота, T.D. Carvalho с соавт. (37) — влияние локуса LEP-R25C на толщину подкожного жира помесных мясных коров, полученных при скрещивании различных пород, в том числе красных ангусов.
Зарубежные исследователи проявляют большой интерес к SNPs в некодирующих участках гена LEP . A. Mandefro с соавт. (38) использовали как экзонные, так интронные SNPs для обнаружения филогенетических связей между различными популяциями крупного рогатого скота. В работе J. Metin Kiyici с соавт. (39) выявилась связь полиморфизма LEP - Sau 3A1, локализованного в интроне 2, с показателями электропроводности, pH и числом соматических клеток в молоке, причем наибольшими значениями выделялись животные с генотипом BB . M. Glantz с соавт. (40) экспериментально определили ряд локусов гена LEP , влияющих на технологические свойства молока: A1457G ( G > A ) и С963Т ( C > T ) оказались ассоциированы с содержанием белка в молоке, а полиморфизмы A1457G, A59V, C963T и T945M — с содержанием кальция в молоке и его активной кислотностью (40).
Несмотря на достаточно большое число отечественных работ по обсуждаемой проблеме, подавляющее большинство из них касается только черно-пестрой породы. Малочисленные российские породы, такие как костромская и ярославская, упоминаются в единичных публикациях (16, 22), актуальность которых быстро снижается. Поэтому изучение полиморфизмов гена лептина и их влияния на хозяйственно полезные признаки молочного скота отечественных малочисленных пород по-прежнему остается актуальным.
В настоящей работе впервые получены данные о наличии достоверной ассоциации локуса гена лептина LEP - A80V с хозяйственно полезными признаками у коров черно-пестрой породы на территории Костромской области, что доказывает значимость этих генотипов в характеристике племенных и продуктивных качеств животных региона.
Цель работы заключалась в изучении ассоциативных связей различных аллельных вариантов гена лептина с показателями молочной продуктивности у коров из племенных хозяйств Костромской области.
Ìåòîäèêà. Объектом исследований был крупный рогатый скот ( Bos taurus ) костромской, черно-пестрой и ярославской пород из трех племенных хозяйств (Костромская обл., 2019-2021 годы): СПК «Гридино», СПК «Расловское» и ООО «Ладыгино». Выборка ( n = 141) включала 33 коровы костромской породы, 47 — ярославской и 61 — черно-пестрой. Содержание круглогодичное стойловое, привязное. Кормление животных осуществлялось согласно внутрихозяйственным рационам, сбалансированным по основным питательным веществам, витаминам, макро- и микроэлементам, в соответствии с молочной продуктивностью и физиологическим состоянием.
Среди коров костромской породы особи с долей кровности < 50 % по улучшающей швицкой породе составляли 75,5 %, с долей кровности 50 % и > 50 % — по 12,1 % изучаемой выборки. В группе ярославской породы распределение было следующим: животные с долей кровности < 50 % по голштинской породе составили 89,4 % всей выборки, 50 % — 6,4 %, > 50 % — 4,2 %. В выборке животных черно-пестрой породы 88,5 % особей имели долю крови > 50 % по улучшающей породе, 11,5 % — кров-ность 50 %, коровы с долей кровности < 50 % отсутствовали.
Полиморфные варианты гена лептина LEP определяли методом полимеразой цепной реакции в реальном времени (ПЦР-РВ) по трем локусам: LEP-R25C (rs29004488 С > T, аллели R , C ; генотипы RR , RC , CC ), LEP-A80V (rs29004508 С > T, аллели A , V ; генотипы AA , AV , VV ) и LEP-Y7F (rs29004487 A > T), аллели Y , F ; генотипы YY , YF , FF ) с использованием амплификатора DTprime (ООО «НПО ДНК-технология», Россия) и технологии HRM (high resolution melting, плавление с высоким разрешением).
Для амплификации целевых фрагментов гена LEP применяли праймеры, исключающие образование неспецифических ПЦР-продуктов. Размер целевых ПЦР-продуктов составил 70 п.н. По полиморфизму LEP-R25C праймеры R25C_forw TCGATTCCTGTGGCTTTGG и R25C_rev CCGTGT-GTGAGATGTCATTG, зонды типирующие (флуоресцентные) 25C_C GC-CCATCCGCAAGGTCC-FAM и R25C_T GCCCATCTGCAAGGTCC-VIC. По полиморфизму LEP-A80V праймеры A80V_forw GGGTCACTGGTTTG-GACTTC и A80V_rev GAGGTTCTCCAGGTCAYTGG, зонды типирующие (флуоресцентные) A80V_C CAGACATTGGCGATCTACC-FAM и A80V_T CAGACATTGGTGATCTACC-VIC. По полиморфизму LEP-Y7F праймеры Y7F_for GTGGTTTCTTCTGTTTTCAGGИ и Y7F_rev GGTGTCATCCT-GGACCTTG, зонды типирующие (флуоресцентные) Y7F_A GACCCCT-GTATCGATTCCT-FAM и Y7F_T GACCCCTGTTTCGATTCCT-VIC.
В качестве флуоресцентного ДНК-интеркалирующего красителя в
|
составе ПЦР-смеси использовали EvaGreen® («Biotium», США). Условия амплификации и плавления (диссоциации) продуктов реакции оптимизировали для получения наиболее стабильных результатов HRM-исследования. Амплификацию проводили по следующей программе: |
|||||
|
Блок Температура, ° С Продолжительность, мин:с |
Регистрация Число циклов флуоресценции |
Тип блока |
|||
|
1 |
94 |
5:00 |
1 |
Цикл |
|
|
2 |
94 |
0:30 |
5 |
Цикл |
|
|
64 |
0:15 |
+ |
|||
|
3 |
94 |
0:05 |
30 |
Цикл |
|
|
64 |
0:15 |
+ |
|||
|
4 |
94 |
1:00 |
1 |
Цикл |
|
|
5 |
45 |
0:15 |
4 |
+ |
Цикл |
|
6 |
70 ^ 90 |
0:10 |
100 |
+ |
Плавление |
Плавление продуктов амплификации проводили в диапазоне от 70 до 90 ° С с увеличением температуры на 0,2 ° С каждые 10 с. Учет и анализ результатов осуществляли с помощью ПО RealTime_PCR с применением модуля HRM-анализа. Учитывали данные равномерности нагрева термоблока амплификатора (то есть к результатам кластеризации дополнительно применяли опционную настройку температурной коррекции). Результаты предварительных HRM-протоколов верифицировали методом прямого секвенирования по Сэнгеру с применением пар праймеров, позволявших амплифицировать более длинные фрагменты целевых последовательностей LEP , чем праймеры для HRM-исследований. Для секвенирования экзона 2 c полиморфизмами LEP-Y7F и LEP-R25C использовались праймеры for GTGGTTTCTTCTGTTTTCAGG и rev CCGTGTGTGAGATGTCATTG, для начального участка экзона 3 с полиморфизмом LEP-A80V — праймеры for GGGTCACTGGTTTGGACTTC и rev GAGGTTCTCCAGGTCATTGG.
Сведения о молочной продуктивности животных за 1-ю и 3-ю лактации (удой за 305 сут лактации, процентное содержание молочного жира и молочного белка) были получены на основании ежемесячных контрольных доек согласно ГОСТ Р 57878-2017 (М., 2020). При анализе параметров продуктивности использовали данные зоотехнического учета племенных хозяйств СПК «Гридино», СПК «Расловское» и ООО «Ладыгино» за 20192021 годы из базы данных ИАС «СЕЛЭКС».
Полученные результаты обрабатывали методами популяционно-генетического анализа с использованием программных возможностей Мic-rosoft Excel. Статистическую обработку показателей молочной продуктивности осуществляли при помощи языка статистического программирования R (версия 4.0.6). Для определения соответствия распределения вариантов в выборках нормальному типу применяли тест Шапиро-Уилка (функция shapiro.test), при этом нулевая гипотеза состояла в том, что тестируемая выборка соответствует нормальному типу распределения. Выборки, прошедшие shapiro.test со значением p-value < 0,95, считались не соответствующими нормальному типу распределения. По результатам теста Шапиро-Уилка при обработке сведений о молочной продуктивности по всем выборкам было принято решение использовать непараметрические методы статистики.
По каждой группе животных определенного генотипа в таблицах данные приводятся в виде медианы (Me) с указанием ее 95 % доверительного интервала в виде диапазона из минимального и максимального значений (ДИ1-ДИ2), рассчитанных при помощи рангового одностороннего теста Манна-Уитни в модификации Уилкоксона (wilcox.test). Нулевая гипотеза этого теста состояла в том, что вычисленное значение не было истинной медианой выборки. В случае когда значение медианы, вычислен- ное при помощи функции wilcox.test, имело p-value > 0,05 и расчет ее 95 % доверительного интервала оказывался невозможен, для вычисления использовалась функция median(), возвращающая значение второго квартиля выборки.
Для выявления влияния изучаемых полиморфизмов гена LEP на мо- лочную продуктивность коров оценивали статистическую значимость разницы между групповыми медианами посредством нахождения U-критерия, рассчитанного при помощи рангового двустороннего теста Манна-Уитни в модификации Уилкоксона (wilcox.test) при уровне статистической значимости p < 0,05. Нулевая гипотеза теста состояла в том, что разница между истинными медианами двух тестируемых групп равна 0.
С целью уточнения результатов попарного сравнения медианных показателей молочной продуктивности выборок также рассчитывали H-критерий с помощью рангового теста Краскела-Уоллиса (kruskal.test) для трех выборок, соответствующих разным генотипам одного локуса, при уровне статистической значимости p < 0,05. Нулевая гипотеза теста состояла в том, что разница между медианами всех трех групп равна 0, альтернативная — что медиана хотя бы одной группы отличается от двух остальных.
Для расчетов использовали следующие формулы:
-
f = m / N , где f — частота генотипа, m — число особей, имеющих определенный генотип, N — общее число особей;
f p = (2 n AA + n ab )/2 N и f q = (2 n BB + n ab )/2 N , где f p — частота аллеля A , f q — частота аллеля B , n aa , n ab , n bb — число особей с генотипом соответственно АА , АВ и BB , N — общее число особей;
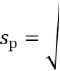
fP-fq
2-N где sp — ошибка частоты аллелей, fp — частота аллеля A, fq — частота аллеля B, N — общее число особей;
, где spp — ошибка частоты генотипа АА, /pp, /pq, /qq — частоты генотипов соответственно АА, АВ и BB, N — общее число особей.
Результаты. При анализе частоты встречаемости генотипов полиморфизма A80V по гену лептина было установлено, что генотип АА с наибольшей частотой регистрировался в популяции КРС черно-пестрой породы — 0,541 (табл. 1).
1. Частота встречаемости генотипов и аллелей локуса LEP-A80V у коров разных пород из племенных хозяйств Костромской области (f ± s , 2021 год)
|
Порода |
n |
Генотипы |
Аллели |
Гетерозиготность |
χ 2 |
|
АА \ АУ \ VV |
A | V |
H o H e |
Костромская 33 0,424±0,085 0,515±0,087 0,061±0,042 0,682±0,057 0,318±0,057 0,515 0,434 1,16 Черно-пестрая 61 0,541±0,064 0,410±0,063 0,049±0,028 0,746±0,039 0,254±0,039 0,419 0,383 17,9* Ярославская 47 0,447±0,072 0,447±0,072 0,106±0,045 0,670±0,048 0,330±0,048 0,447 0,414 0,14 Примечание. H o — наблюдаемая, H e — теоретически ожидаемая гетерозиготность.
* p < 0,01.
У коров костромской породы чаще встречался генотип AV (0,515). У животных ярославской породы генотипы АА и AV определялись с одинаковой частотой — 0,447, а генотип VV имел большую частоту встречаемости относительно костромской и черно-пестрой пород. При этом гено- тип VV характеризовался наименьшей частотой встречаемости у всех животных. У всех трех пород коров по полиморфизму A80V чаще всего встречался аллель À (см. табл. 1).
Распределение аллелей и генотипов локуса LEP - A80V в популяциях костромской и ярославской породы соответствовало уравнению Харди-Вайнберга, что свидетельствовало о соблюдении генного равновесия. В то же время у коров черно-пестрой породы наблюдалось статистически значимое превышение наблюдаемой гетерозиготности над теоретически ожидаемой (p < 0,01), что указывало на преимущество гетерозиготных животных при размножении в этой популяции.
По результатам анализа частоты встречаемости генотипов полиморфизма Y7F было установлено, что среди трех пород КРС наиболее часто встречался аллель Y (0,893-0,970), при этом среди генотипов преобладал гомозиготный вариант YY (табл. 2).
2. Частота встречаемости генотипов и аллелей локуса LEP-Y7F у коров разных пород из племенных хозяйств Костромской области ( f ± s , 2021 год)
|
Порода |
n |
Генотипы |
Аллели |
Гетерозиготность |
χ 2 |
||
|
FF |
YF |
YY |
F Y |
H o 1 H e |
|||
|
Костромская |
33 |
0,030±0,030 |
0 |
0,970±0,030 0,030±0,021 0,970±0,021 |
0 0,059 |
33,0* |
|
|
Черно-пестрая |
61 |
0,098±0,038 |
0,016±0,016 |
0,885±0,041 0,107±0,028 0,893±0,028 0,016 0,191 |
49,6* |
||
|
Ярославская 47 Примечание. H o * p < 0,01. |
0,064±0,036 0,021±0,021 0,936±0,034 0,074±0,027 0,947±0,027 0,021 0,141 — наблюдаемая, H e — теоретически ожидаемая гетерозиготность. |
33,6* |
|||||
Частота встречаемости генотипа YY у животных костромской породы оказалась наибольшей (0,970), при этом гетерозиготные животные YF в выборке отсутствовали. Схожие особенности распределения частот встречаемости генотипов были выявлены у коров черно-пестрой и ярославской пород. Среди них наиболее высокая доля животных с гомозиготным генотипом FF была зарегистрирована среди черно-пестрой породы — 0,098 против 0,064 у ярославской породы.
Во всех трех группах коров по локусу Y7F наблюдалось статистически значимое (p < 0,01) нарушение генного равновесия в сторону недостатка гетерозигот. Это указывает на наличие отбора, направленного на элиминацию аллеля F , связанного с пониженной жизнеспособностью животных, из популяций (8, 9).
Данные по полиморфизму R25C показали, что аллель R имел наибольшую частоту встречаемости, при этом у коров ярославской породы его регистрировали чаще (0,840) (табл. 3). Также у ярославской породы чаще остальных встречался гомозиготный вариант RR (0,702) и гетерозиготный вариант RC (0,277), а встречаемость ÑÑ генотипа была наименьшей.
-
3. Частота встречаемости генотипов и аллелей локуса LEP-R25C у коров разных пород из племенных хозяйств Костромской области ( f ± s , 2021 год)
Порода
n
Костромская Черно-пестрая Ярославская
Генотипы
RR RC □ CC
Аллели
R C
Гетерозиготность
H o □ H e
χ 2
0,424±0,086 0,485±0,087 0,091±0,05 0,667±0,058 0,333±0,058 0,485 0,4440,27
0,475±0,045 0,344±0,061 0,180±0,049 0,648±0,043 0,352±0,043 0,339 0,4533,95*
0,702±0,067 0,277±0,065 0,021±0,021 0,840±0,038 0,160±0,038 0,277 0,2680,05
Примечание. H o — наблюдаемая, H e — теоретически ожидаемая гетерозиготность.
*p < 0,05.
4. Показатели продуктивности коров разных пород и генотипов локуса
LEP-A80V
из племенных хозяйств Костромской области в зависимости от лактации
(
Ìå
, ДИ1-ДИ2, 2019-2021 годы)
У коров черно-пестрой породы аллель Ñ и генотип CC имели наибольшую частоту встречаемости по сравнению с другими породами (0,352 и 0,180 соответственно). В то же время у коров костромской породы наиболее часто определяли генотипы RC и RR.
По локусу LEP-R25C в популяциях скота костромской и ярославской пород нарушение генного равновесия не было зафиксировано. При этом в группе коров черно-пестрой породы наблюдался статистически значимый избыток гетерозигот (p < 0,05). Это указывает на преимущество животных, имеющих гетерозиготный генотип RC , при размножении в указанной популяции.
Проверка характера распределения вариантов в группах коров при помощи теста Шапиро-Уилка показала несоответствие нормальному типу распределения как минимум одной из групп (генотипов) в каждой породе по каждому показателю. Поэтому для описания выборок животных с разными генотипами по генe LEP для каждого параметра молочной продуктивности вычисляли медиану и ее 95 % доверительный интервал, а сравнение медиан выборок выполняли при помощи U-критерия (тест Манна-Уитни в модификации Уилкоксона).
Сравнение продуктивности коров-первотелок разных генотипов по локусу LEP-A80V (табл. 4) при помощи U-критерия не позволило определить в группе животных костромской породы статистически значимую разницу между носительницами генотипов AA и AV . В группах первотелок других пород разница между гомозиготными и гетерозиготными животными оказалась более выраженной, но уровня статистической значимости не достигла. По содержанию компонентов в молоке также не было найдено статистически значимых различий.
|
Генотип |
1-я лактации |
3-я лактации и старше |
||||
|
n |
удой, кг |
МДЖ, % \ МДБ, % |
n \ удой, кг |
МДЖ, % 1 |
МДБ, % |
|
|
AA |
14 |
5011,0 |
Костромская 4,120 3,275 |
порода 11 5838,0 |
4,260 |
3,225 |
|
AV |
17 |
(4322,5-5612,0) 5034,0 |
(3,950-4,330) (3,190-3,320) 4,200 3,255 |
(5494,0-6329,0) 14 6226,5 |
(4,050-4,530) 4,250 |
(3,165-3,280) 3,245 |
|
VV |
2 |
(4387,5-5815,5) 7246,0 |
(3,970-4,370) (3,180-3,325) 4,160 3,295 |
(5807,0-6816,0) 1 7505,0 |
(4,060-4,410) 3,880 |
(3,200-3,285) 3,240 |
|
AA |
33 |
7477,5 |
Черно-пестрая порода 3,940 3,125 19 8856,0 |
3,970 |
3,120 |
|
|
AV |
25 |
(6833,5-8024,0) 7115,0 |
(3,835-4,060) (3,085-3,170) 3,980 3,175 |
(8819,2-9394,0)* 10 8563,6 |
(3,880-4,000)* 4,050 |
(3,075-3,145) 3,120 |
|
VV |
2 |
(6515,5-7604,5) 7439,0 |
(3,770-4,180) (3,130-3,220) 3,670 3,080 |
(8190,6-8819,2) 2 8819,2 |
(3,990-4,210) 3,980 |
(3,120-3,140) 3,130 |
|
AA |
21 |
4697,0 |
Ярославская 3,885 3,160 |
порода 20 5707,0 |
3,810 |
3,175 |
|
AV |
21 |
(4295,0-5105,0) 4381,0 |
(3,785-3,995) (3,110-3,225) 3,995 3,190 |
(5322,0-6294,5) 16 5604,3 |
(3,675-3,940) 3,870 |
(3,120-3,225) 3,215 |
|
VV |
5 |
(3999,5-4837,5) 4850,0 |
(3,865-4,085) (3,125-3,255) 3,940 3,130 |
(5338,5-5812,3) 2 5709,7 |
(3,830-3,920) 3,830 |
(3,180-3,245) 3,195 |
Примечание. МДЖ — массовая доля жира, МДБ — массовая доля белка.
* Различия между носителями генотипов AA и AV статистически значимы при p < 0,05.
К 3-й лактации разница по медианному удою между гомозиготными и гетерозиготными животными костромской породы стала более выраженной в сторону гетерозигот (с 23 кг по 1-й лактации до 388,5 кг по 3-й), но статистически значимого различия между группами не было выявлено. У коров черно-пестрой породы по 3-й лактации более высоким медианным удоем отличались гомозиготы AA по сравнению с гетерозиготами AV (на 292,4 кг), причем эта разница достигла уровня статистической значимости (p < 0,05). По массовой доле жира (МДЖ) в молоке среди полновозрастных коров черно-пестрой породы в наибольшую сторону выделя- лись носительницы гетерозиготного генотипа AV, значимо превосходя своих гомозиготных (AA) сверстниц на 0,08 % (p < 0,05).
Среди полновозрастных коров ярославской породы по удою и содержанию компонентов в молоке статистически значимых различий между генотипами не выявлено.
При сравнении показателей молочной продуктивности коров разных генотипов по локусу LEP-Y7F пришлось столкнуться с выраженным дефицитом гетерозиготных животных, что позволило провести сравнение только среди коров черно-пестрой породы между носительницами гомозиготных вариантов YY и FF гена лептина (табл. 5).
5. Показатели продуктивности коров разных пород и генотипов локуса LEP-Y7F из племенных хозяйств Костромской области в зависимости от лактации ( Ìå , ДИ1-ДИ2, 2019-2021 годы)
|
Генотип |
1-я лактации |
3-я лактации и старше |
||||
|
n |
удой, кг |
МДЖ, % □ МДБ, % |
n ] удой, кг \ |
МДЖ, % |
МДБ, % |
|
|
FF |
1 |
4833,0 |
Костромская 3,950 3,280 |
порода 1 5515,0 |
4,000 |
3,150 |
|
YF YY |
0 32 |
5171,0 |
4,180 3,260 |
25 6115,7 |
4,255 |
3,240 |
|
FF |
6 |
(4681,0-5650,5) 6297,5 |
(4,030-4,300) (3,210-3,310) (5827,5-6559,0) Черно-пестрая порода 3,720 3,170 2 8630,6 |
(4,130-4,375) 4,000 |
(3,210-3,270) 3,120 |
|
|
YF |
1 |
(4976,0-8030,0) 6601,0 |
(3,600-3,840) (3,080-3,250) 4,460 3,000 |
1 8893,0 |
4,310 |
3,230 |
|
YY |
54 |
7446,0 |
3,975 3,140 |
28 8819,2 |
3,995 |
3,120 |
|
FF |
3 |
(7011,5-7840,5) 4376,0 |
(3,870-4,085) (3,115-3,175) Ярославская 3,740 3,150 |
(8755,6-8856,6) порода 3 5804,2 |
(3,960-4,000) 3,720 |
(3,115-3,125) 3,190 |
|
YF |
1 |
4046,0 |
3,890 3,060 |
1 5830,0 |
4,370 |
3,260 |
|
YY |
43 |
4618,1 |
3,945 3,180 |
34 5646,7 |
3,840 |
3,195 |
|
(4352,5-4909,0) (3,680-3,890) (3,135-3,225) Примечание. МДЖ — массовая доля жира, МДБ — |
(5491,5-5815,0) массовая доля белка. |
(3,796-3,900) |
(3,180-3,225) |
|||
Анализ показателей молочной продуктивности коров-первотелок черно-пестрой породы показал тенденцию к наибольшему медианным удою и МДЖ в молоке у носительниц генотипа YY по сравнению с гомозиготными ( FF ) сверстницами — соответственно на 2275 кг и 0,075 %, однако по результатам расчета U-критерия отвергнуть нулевую гипотезу теста Манна-Уитни об отсутствии различий между группами не удалось (p > 0,05). В силу недостаточного числа полновозрастных коров с генотипом FF сравнить их со сверстницами с генотипом YY не представилось возможным.
Сравнение показателей молочной продуктивности коров-первотелок не позволило выявить статистически значимых различий между генотипами по локусу LEP R25C как по величине удоя, так и по содержанию компонентов молока (табл. 6).
Среди коров-первотелок костромской породы тенденцию к наибольшему медианному удою и МДЖ в молоке проявляли носительницы генотипа ÑÑ по сравнению с гетерозиготными сверстницами — соответственно на 883,3 кг и 0,13 %. Эта тенденция наблюдалась и у полновозрастных животных, но разница по показателям удоя и МДЖ между носительницами генотипов CC и RC была существенно меньше — соответственно 244,5 кг и 0,035 %. По массовой доле белка (МДБ) в молоке значимых различий между носительницами разных генотипов по локусу LEP-R25C не наблюдалось.
Среди первотелок ярославской породы проявилась аналогичная тенденция, но в гораздо меньшей степени. Так, коровы с генотипом RR за 1-ю лактацию имели более высокими медианными показателями удоя и 1100
МДЖ в молоке (на 127 кг и 0,035 %), чем их сверстницы RC , при этом к 3-й лактации эта разница практически нивелировалась. По МДБ существенных различий между носительницами разных генотипов по локусу LEP R25C отмечено не было.
6. Показатели продуктивности коров разных пород и генотипов локуса LEP-R25C из племенных хозяйств Костромской области в зависимости от лактации ( Ìå , ДИ1-ДИ2, 2019-2021 годы)
|
Гено- |
1- |
я лактации |
3-я лактации и старше |
||||
|
тип |
n ] удой, кг |
МДЖ, % |
МДБ, % |
n |
удой, кг \ |
МДЖ, % 1 |
МДБ, % |
|
Костромская порода |
|||||||
|
ÑÑ |
14 5504,0 |
4,180 |
3,280 |
12 |
6241,0 |
4,255 |
3,250 |
|
(4803,0-6143,5) |
(3,940-4,365) |
(3,205-3,355) |
(5823,0-6913,0) |
(4,045-4,460) |
(3,200-3,300) |
||
|
RC |
16 4620,7 |
4,050 |
3,230 |
12 |
5996,5 |
4,220 |
3,210 |
|
(3992,0-5322,0) |
(3,950-4,285) |
(3,165-3,295) |
(5547,5-6574,0) |
(4,040-4,410) |
(3,165-3,255) |
||
|
RR |
3 6175,0 |
4,690 |
3,390 |
2 |
6065,0 |
4,480 |
3,290 |
|
Черно-пестрая |
порода |
||||||
|
CC |
29 7138,5 |
4,035 |
3,155 |
15 |
87781,0 |
4,000 |
3,120 |
|
(6631,0-7675,0) |
(3,885-4,230) |
(3,110-3,205) |
(8474,5-8819,2) |
(3,990-4,165) |
(3,100-3,135) |
||
|
RC |
21 7184,5 |
3,900 |
3,140 |
10 |
8819,2 |
4,000 |
3,120 |
|
(6292,0-7949,0) |
(3,755-4,080) |
(3,095-3,185) |
(8630,6-8901,0) |
(3,910-4,120) |
(3,120-3,220) |
||
|
RR |
11 7838,0 |
3,815 |
3,100 |
5 |
8819,2 |
3,840 |
3,090 |
|
(6768,5-8920,0) |
(3,665 4,040) |
(3,025-3,180) |
(7877,6-10398,6) |
(3,540-4,000) |
(2,970-3,125) |
||
|
Ярославская порода |
|||||||
|
CC |
33 4615,0 |
3,925 |
3,165 |
26 |
5642,3 |
3,840 |
3,195 |
|
(4298,0-5010,0) |
(3,825-4,015) |
(3,120-3,220) |
(5491,8-5830,5) |
(3,780-3,910) |
(3,175-3,230) |
||
|
RC |
13 4488,0 |
3,890 |
3,170 |
12 |
5693,5 |
3,825 |
3,210 |
|
(4070,0-4936,5) |
(3,77-4,04) |
(3,095-3,265) |
(5064,0- 6260,0) |
(3,645-3,970) |
(3,155-3,260) |
||
|
RR |
1 4850,0 |
4,440 |
3,330 |
0 |
|||
|
Примечание. МДЖ — |
массовая доля жира, МДБ |
— массовая доля белка. |
|||||
В группе первотелок черно-пестрой породы тенденцию к наибольшему медианному удою и наименьшему медианному МДЖ в молоке проявляли животные с генотипом RR : они превосходили коров с RC и CC по удою на 653,5 кг и 699,5 кг, по МДЖ — уступали на 0,085 и 0,22 %. Аналогичная тенденция, но в существенно меньшем масштабе была характерна и для полновозрастных коров. При этом по МДБ в молоке животные разных генотипов по локусу LEP-R25C не имели достоверных различий.
Для оценки влияния генотипов гена лептина по локусам LEP-A80V , LEP-Y7F и LEP-R25C на основные признаки молочной продуктивности коров был рассчитан H-критерий при помощи рангового теста Краскела-Уоллиса, альтернативной гипотезой которого было то, что хотя бы одна из групп животных отличается от двух других.
В результате среди полновозрастных коров черно-пестрой породы были обнаружены значимые различия между генотипами по удою и содержанию жира в молоке, при которых удалось принять альтернативную гипотезу теста Краскела-Угллиса (p < 0,05). В то же время H-критерий для выборок коров костромской и ярославской пород не выявил статистически значимых различий между носительницами разных генотипов по локусу LEP-A80V (табл. 7). При этом по удою за 1-ю лактацию у коров костромской породы и по удою за 3-ю у черно-пестрой отмечалась существенная разница между носительницами разных генотипов этого локуса (p = 0,09 и p = 0,059).
Расчет H-критерия для выборок, сформированных согласно генотипу коров по локусу LEP-Y7F (табл. 8), показал отсутствие статистически значимых различий между носительницами разных генотипов (p > 0,05). Тем не менее у первотелок черно-пестрой породы отмечалась существенная тенденция к различиям между носительницами разных генотипов локуса LEP-Y7F по МДЖ в молоке (p = 0,095).
Вычисление H-критерия для выборок коров, сформированных со- гласно их генотипу по локусу LEP-R25C, не показало значимых различий между носительницами разных генотипов (табл. 9). Однако при этом наблюдалась выраженная тенденция к разнице групповых медиан по удою и МДЖ в молоке у коров-первотелок костромской породы (соответственно p = 0,055 и p = 0,099) и по МДЖ — среди полновозрастных коров черно-пестрой породы (p = 0,131).
-
7. Влияние генотипов по локусу LEP-A80V на показатели молочной продуктивности (χ2) у коров разных пород из племенных хозяйств Костромской области (H-критерий Краскела-Уоллиса, 2019-2021 годы)
Показатель
Порода
костромская черно-пестрая
ярославская
1-я лактация
Удой
4,80440
0,93286
3,68010
МДЖ
0,15723
0,19640
1,89260
МДБ
0,27790
3,73290
1,27840
3-я лактация
и старше
Удой
3,07620
5,65090
0,80283
МДЖ
0,61714
7,58910*
1,50810
МДБ
0,43705
1,52180
2,37430
Примечание. МДЖ — массовая доля жира, МДБ
— массовая доля белка.
* р < 0,05.
-
8. Влияние генотипов по локусу LEP-Y7F на показатели молочной продуктивности (χ2) у коров разных пород из племенных хозяйств Костромской области (H-критерий Краскела-Уоллиса, 2019-2021 годы)
Показатель
Порода
костромская
черно-пестрая
ярославская
Удой
1-я лакт 0,09928
ация
3,73610
1,09300
МДЖ
0,39739
4,70690
2,33560
МДБ
0,02487
2,25130
1,40210
Удой
3-я лактация 1,11370
и старше 2,14100
0,85337
МДЖ
0,40094
2,08300
3,95310
МДБ
1,11560
1,79810
0,93826
Примечание. МДЖ — массовая доля жира, МДБ
— массовая доля белка.
-
9. Влияние генотипов по локусу LEP-R25C на показатели молочной продуктивности (χ2) у коров разных пород из племенных хозяйств Костромской области (H-критерий Краскела-Уоллиса, 2019-2021 годы)
Показатель
Порода
костромская 1
черно-пестрая
ярославская
Удой
1-я лакт 5,78470
ация
1,98060
0,40390
МДЖ
4,61490
2,07350
2,97570
МДБ
3,71910
1,35250
1,48530
Удой
3-я лактация 1,24840
и старше
2,23750
0,07228
МДЖ
2,41170
4,06450
0,27573
МДБ
3,50940
2,17900
0,13592
Примечание. МДЖ — массовая доля жира, МДБ
— массовая доля белка.
Генетическая структура изученных популяций коров костромской, ярославской и черно-пестрой пород из племенных хозяйств Костромской области по локусу LEP-A80V характеризовалась преобладанием аллеля A, в большей степени выраженным у черно-пестрого скота (A:V = 0,746:0,254). Доминирование аллеля À в группах животных костромской и ярославской пород было менее выражено, и соотношение аллелей A:V составляло соответственно 0,682:0,318 и 0,67:0,33. Наши результаты в целом соответствуют сведениям, полученным другими авторами, хотя у них для аллеля À харак- терна более высокая доля: для черно-пестрой породы — 0,86, для ярославской — 0,78, для швицкой породы, родственной с костромской, — 0,81 (8, 22). В имеющихся зарубежных исследованиях полиморфизма LEP-A80V частота аллеля À также заметно выше — от 0,775 до 0,900, однако эти работы проводились на животных джерсейской породы и ее помесей с местным кашмирским скотом (20, 21).
Подобный сдвиг в соотношении аллелей связан, по-видимому, с целенаправленной селекцией на повышение удоя коров и положительной ассоциацией аллеля LEP-A80VA с обильномолочностью. Эта связь установлена в большом числе зарубежных и отечественных публикаций (21, 24, 25, 27) и частично подтвердилась в нашем исследовании. Примечательно, что в черно-пестрой породе наибольшим удоем выделялись коровы с генотипом ÀÀ , как в исследованиях Ф.Ф. Зиннатовой (27), а в группах коров костромской и ярославской пород статистически значимая разница между генотипами полиморфизма LEP-A80V по удою мы не обнаружили.
В нашей работе показана значимая связь между локусом LEP-A80V и содержанием жира в молоке коров черно-пестрой породы, причем наибольшей МДЖ отличались гетерозиготные животные, что согласуется с результатами ряда авторов (27-30). При этом у коров костромской и ярославской пород мы не установили статистически значимого влияния полиморфизма A80V на содержание компонентов в молоке.
Генетическая структура изученных популяций по локусу LEP-Y7F характеризовалась доминированием по частоте встречаемости генотипа YY и аллеля Y . Наибольшей частотой встречаемости аллеля Y выделялись коровы костромской породы (0,97), что согласуется с данными А.В. Перчуна с соавт. (16). В группе животных черно-пестрой породы частота встречаемости аллеля F была наибольшей среди всех групп (0,104) и при этом существенно выше, чем частота этого же аллеля в аналогичных исследованиях Н.В. Ковалюк с соавт. (17).
Крайняя редкость генотипов YF и FF может быть связана с негативным влиянием аллеля F на энергетический обмен в организме, что отражается, в том числе, на росте и развитии животных (36). Кроме того, аллель F , по данным многих авторов, негативно влияет на показатели молочной продуктивности (18, 19). Нам не удалось установить статистически значимую разницу по показателям молочной продуктивности между носительницами разных генотипов по локусу LEP-Y7F , что согласуется с данными ряда авторов (18, 19). Отмечалась тенденция к различию между значениями показателя МДЖ в молоке у коров черно-пестрой породы разных генотипов по этому локусу, однако для получения статистически значимых результатов необходимо проведение экспериментов с более обширным объемом выборки.
В результате генотипирования животных по локусу LEP-R25C выяснилось, что во всех трех группах преобладал аллель Ñ , при этом наибольшая представленность аллеля R наблюдалась у черно-пестрой породы (0,352), наименьшая — в ярославской (0,16), а коровы костромской породы занимали промежуточное положение (0,333). В целом подобное распределение аллелей соответствует данным как зарубежных, так и российских ученых (8, 31, 32).
В нашем исследовании были отмечены тенденции к различию между носителями разных генотипов по локусу LEP-R25C по показателям молочной продуктивности, но порог статистической значимости не был преодолен. При этом результаты тестирования выборок по H-критерию методом
Краскела-Уоллиса показали тенденцию влияния полиморфизма LEP-R25C на удой и МДЖ в молоке у первотелок костромской породы, но статистической значимости не удалось достигнуть и в этом случае, хотя похожие данные были получены другими авторами (25, 34, 35). Это может быть связано с тем, что проанализированный в нашей работе размер поголовья не позволил использовать более чувствительные параметрические методы статистики и, следовательно, повысилась вероятность возникновения статистической ошибки второго рода при проверке гипотез.
Таким образом, во всех изученных нами группах коров костромской, черно-пестрой и ярославской пород из племенных хозяйств Костромской области по локусу LEP-A80V преобладал аллель A . При этом только в группе животных черно-пестрой породы наблюдался статистически значимый сдвиг генного равновесия по этому локусу в сторону гомозигот, в этой же группе полиморфизм LEP-A80V оказывал статистически значимое влияние на удой и содержание жира в молоке и аллель А был положительно ассоциирован с обильномолочностью и жирномолочностью. По локусу LEP-Y7F во всех изученных группах животных отмечалось статистически значимое смещение генного равновесия в сторону гомозигот FF. Однако установить влияние этого полиморфизма на показатели молочной продуктивности не удалось в силу крайне редкой встречаемости аллеля F и генотипов YF и FF. При изучении полиморфизма LEP-R25C только среди животных черно-пестрой породы обнаружился статистически значимый сдвиг генного равновесия в сторону гомозигот, однако и в этой группе не выявлено признаков достоверного влияния локуса на показатели молочной продуктивности. Полученные результаты отражают степень генетического разнообразия племенного крупного рогатого скота Костромской области, дополняя и актуализируя данные по основным отечественным малочисленным породам. Установлена достоверная ассоциация гена лептина с молочной продуктивностью и содержанием жира в молоке. Выявленные нами тенденции еще раз указывают на необходимость сохранения генетического разнообразия крупного рогатого скота. Дальнейшие исследования предполагают массовый скрининг высокоинтенсивных пород животных и значительное увеличение тестируемого поголовья.
