Ассоциация полиморфных вариантов гена CAPN1 с живой массой у овец калмыцкой курдючной породы и ее помесей с породами Шароле и Дорпер

Автор: Юлдашбаев Ю.А., Селионова М.И., Белая Е.В., Чылбак-оол С.О., Арилов А.Н., Пахомова Е.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Зоотехния и ветеринария
Статья в выпуске: 12, 2023 года.
Бесплатный доступ
Цель исследования - изучение фенотипического эффекта генотипов полиморфного гена кальпаина (CAPN1) на живую массу у овец пород калмыцкая курдючная и их помесей с породами шароле и дорпер для обоснования возможности генетического маркирования продуктивности в раннем возрасте. Задачи - проведение генотипирования и учета показателей продуктивности животных калмыцкой курдючной породы (КК), шароле (Ш), дорпер-калмыцких помесей овцематок (1/2Д×1/2КК), полученного чистопородного и помесного молодняка; установление влияния генотипов по гену CAPN1 на живую массу и среднесуточный прирост. Установлено, что у баранов-производителей калмыцкой курдючной породы и шароле в гене CAPN1 чаще выявлялись носители аллеля С, тогда как среди чистопородных и помесных овцематок встречаемость аллелей С и Т была равной. У чистопородного молодняка и помесей ½Ш×½КК преобладали особи с аллелем Т, у помесей 1/2Ш×1/4Д×1/4КК - носители аллеля С. Популяционно-генетический анализ ни в одной из групп не выявил достоверных отличий наблюдаемых частот генотипов от теоретически ожидаемых при равновесии Харди-Вайнберга. Независимо от породной принадлежности и пола живая масса молодняка при рождении разных генотипов по гену CAPN1 была схожей, тогда как в четырехмесячном возрасте,как баранчики, так и ярочки с генотипом СС имели достоверное превосходство над сверстниками других генотипов. У чистопородных баранчиков наибольшая разность наблюдалась между животными генотипов СС и СТ, у помесей 1/2Ш×1/2КК - животных генотипов СС и ТТ, которая составила 10,4 и 8,1 %; 9,94 и 10,95 % (p function show_abstract() { $('#abstract1').hide(); $('#abstract2').show(); $('#abstract_expand').hide(); }
Овцы, калмыцкая курдючная порода, шароле, живая масса, кальпаин, генотип
Короткий адрес: https://sciup.org/140304267
IDR: 140304267 | УДК: 57.032/636.033 | DOI: 10.36718/1819-4036-2023-12-215-224
Association of the CAPN1 gene polymorphic variants with live weight in the Kalmyk fat- tailed sheep and their crosses with Charolais and Dorper breeds
The purpose of research is to study the phenotypic effect of genotypes of the polymorphic calpain gene (CAPN1) on live weight in Kalmyk fat-tailed sheep and their crosses with the Charolais and Dorper breeds to substantiate the possibility of genetic marking of productivity at an early age. Objectives - carrying out genotyping and accounting of productivity indicators of animals of the Kalmyk fat-tailed breed (KK), Charolais (Sh), Dorper-Kalmyk crossbred ewes (1/2D×1/2KK), obtained purebred and crossbred young animals; establishing the influence of genotypes for the CAPN1 gene on live weight and average daily gain. It was found that in Kalmyk fat-tailed and Charolais breeding rams, carriers of the C allele in the CAPN1 gene were more often detected, while among purebred and crossbred ewes the occurrence of the C and T alleles was equal. In purebred young animals and 1/2Sh × 1/2KK crosses, individuals with the T allele predominated; in 1/2Sh × 1/4D × 1/4KK crossbreeds, the carriers of the C allele predominated. Population genetic analysis in none of the groups did not reveal significant differences in the observed genotype frequencies from those theoretically expected at Hardy-Weinberg equilibrium. Regardless of breed and sex, the live weight of young animals at birth of different genotypes for the CAPN1 gene was similar, while at four months of age, both rams and lambs with the CC genotype had a significant superiority over their peers of other genotypes. In purebred rams, the greatest difference was observed between animals of the CC and ST genotypes, in crossbreeds 1/2Sh × 1/2KK - animals of the CC and TT genotypes, which amounted to 10.4 and 8.1 %; 9.94 and 10.95 % (p function show_eabstract() { $('#eabstract1').hide(); $('#eabstract2').show(); $('#eabstract_expand').hide(); }
Текст научной статьи Ассоциация полиморфных вариантов гена CAPN1 с живой массой у овец калмыцкой курдючной породы и ее помесей с породами Шароле и Дорпер
Acknowledgments : this work has been carried out under the topic "Biotechnological methods of reproduction and genomic technologies in the breeding of farm animals and the preservation of the gene pool of small populations" as part of the "Scientific and Technological Frontiers" project of the "Priority 2030" program.
Введение. Основной тенденцией развития овцеводства в последние десятилетия во всем мире является устойчивый рост производства баранины, что определяет увеличение доли специализированных мясных пород и возрастающие требования к качеству баранины [1, 2]. Маркер-ассоциированная селекция может быть одним из инструментов ускорения этого процесса. Исследования генетико-биохимических основ фенотипического полиморфизма признаков, определяющих мясную продуктивность, ведутся уже многие десятилетия. Известно, что большинство показателей продуктивности находится под совместным контролем значительного числа генов. Однако применение генетических маркеров в повышении мясной продуктивности овец по сравнению с другими видами сельскохозяйственных животных до настоящего времени является менее разработанной областью. Тем не менее выявление генов, ассоциированных с мясной продуктивностью, может быть перспективным, поскольку признаки, определяющие рост костной, мышечной и жировой тканей, характеризуются невысокой наследуемостью [3].
Ген кальпаина является одним из генов, который может быть использован в качестве маркера мясной продуктивности овец. Белок каль-паин , кодируемый геном CAPN, принадлежит семейству цитоплазматических кальцийзависи-мых протеиназ с папаиноподобной активностью [4]. Клеточный протеолитический аппарат высокоселективен и строго регулируется, так как избыточная деструкция жизненно необходимых белков или замедленная деградация короткоживущих регуляторных белков могут существенно изменить клеточные функции. Этот механизм, по всей видимости, у многих прокариот -результат совместной деятельности кальпаи-нов, лизосомальных катепсинов и протеасом [5]. Высоковариабельная структура обнаруженных последовательностей кальпаинов и кальпаино-подобных молекул свидетельствует о широком разнообразии выполняемых ими клеточных функций. Кальпаины принимают участие в основных кальцийзависимых клеточных процессах - передаче сигнала, клеточном цикле, пролиферации, дифференцировке, слиянии мембран, формировании мышечных волокон [6, 7]. Разрушение молекулярных комплексов прикрепления цитоскелета к мембране является наиболее выраженной функцией кальпаинов и подтверждается тем, что более 30 белков цитоскелета чувствительны к кальпаинам [8]. Ряд исследований показал, что кальпаин-кальпаста-тиновая система регулирует рост скелетных мышц, ускорение которого может быть результатом уменьшения деградации мышечного белка за счет снижения уровня кальпаина и увеличения активности кальпастатина [9].
Таким образом, полиморфные варианты гена кальпаина могут оказывать фенотипические эффекты на прижизненные количественные признаки у сельскохозяйственных животных, являющиеся внешним проявлением внутренних процессов, контролируемых кальпаиновой системой. Это свойство определило интерес к изу- чению полиморфизма кальпаина у некоторых пород овец [10–12]. Однако в породах овец российской селекции, а также при использовании зарубежного генофонда выполнено недостаточно исследований полиморфизма гена кальпаина, что определило актуальность настоящей работы.
Цель исследования - изучение фенотипического эффекта генотипов полиморфного гена кальпаина ( CAPN1 ) на живую массу у овец пород калмыцкая курдючная и их помесей с породами шароле и дорпер для обоснования возможности генетического маркирования продуктивности в раннем возрасте.
Объекты и методы. Исследование проведено на базе хозяйства «АРЛ» (Республика Калмыкия, Яшкульский район). В хозяйстве используется пастбищно-стойловая система содержания, при которой 285 дней - пастбищный период. Трава естественных пастбищ, представленная в основном полынью, несколькими видами дерно-видных злаков (ковыли, типчаки) и солянкой, занимает 75–80 % годового рациона овец. Дополнительно, в соответствии с физиологическим состоянием овцематок, используется около 7– 10 % концентрированных кормов и 10-17 % заготовленных грубых кормов. Таким образом, их основной рацион состоит из 3–4 кг травы злаково-полынного пастбища, 1,5 кг злаково-бобового сена, 0,25 кг концентрированного корма (50 % ячменя, 40 % кукурузы, 10 % шрота подсолнеч-никового) и 0,08 кг минеральной подкормки.
Отбор образцов для генотипирования и учет показателей продуктивности проводили от животных калмыцкой курдючной породы (КК) мясосального направления продуктивности, шароле (Ш) мясо-шерстного направления продуктивности, а также помесей с шароле и дорпер (Д) -мясного направления продуктивности. Схема скрещивания была следующей: бараны-производители КК (n = 6), возраст 4,5 года, живая масса 89,3±1,1 кг; бараны-производители Ш (n = 2), возраст 3,5 года, живая масса 80,3 ± 1,1 кг; овцематки КК (n = 40): 20 - для чистопородного скрещивания, 20 - с баранами Ш, возраст 3–4 года, живая масса 62,3 ± 0,32 кг; овцематки %Дх%КК (n = 40) возраст 3-4 года, живая масса 62,9 ± 0,31 кг .
Численность и распределение потомства по полу: группа 1 - КК (n = 26: J14 и?12); группа
2 – ½Ш×½КК (n = 32: ♂12 и ♀20), группа 3 – ½Ш×¼Д×¼КК (n = 50: ♂16 и ♀34).
Экстракцию ДНК проводили из цельной крови овец набором ДНК-Экстран-1 («Синтол», Москва) согласно инструкции, предоставленной фирмой-производителем. Образцы геномной ДНК животных анализировали с использованием технологии HRM-анализа на приборе CFX96 (BioRad, США). Использовали следующие праймеры: F 5’-AACATTCTCAACAAAGTGGTG-3’ и R 5’-ACATCCATTACAGCCACCAT-3’. Условия проведения амплификации и HRM-анализа были следующими: 1) 95 °С – 4 мин; 2) (94 °С – 45 с, 62 °С – 45 с, 72 °С – 45 с) × 45 циклов; 3) 72 °С – 7 мин [13, 14]. Анализ результатов проводили с помощью программного обеспечения для HRM-анализа Precision Melt Analysis™ software.
Живую массу животных разных генотипов устанавливали путем взвешивания. У молодняка учитывали данный показатель при рождении и в 4 месяца. По разности значений и периода учета определяли среднесуточный прирост. Полученный материал обрабатывали биометрически, используя программу MS Excel. Достоверность различий сравниваемых показателей по группам оценивали по критерию Стьюдента с уровнем значимости не ниже p < 0,05.
Результаты и их обсуждение. При генотипировании использовали HRM-анализ (High Resolution Melts, HRM), основанный на определении различий в кривых плавления (диссоциации ДНК) после проведения ПЦР в реальном времени с помощью специального программного обеспечения. При плавлении продукта ПЦР двойная спираль ДНК диссоциирует с высвобождением интеркалирующего красителя и снижением уровня флуоресценции. Скорость этого процесса в зависимости от температуры отслеживается с помощью специального программного обеспечения, которое преобразует полученные данные в график кривой плавления. Изменения в графике могут быть очень небольшие, в доли градуса, однако этого достаточно, чтобы выявить однонуклеотидные замены (SNP), небольшие инсерции, делеции и метилирование ДНК [15].
Визуализация результатов генотипирования представлена на рисунке.
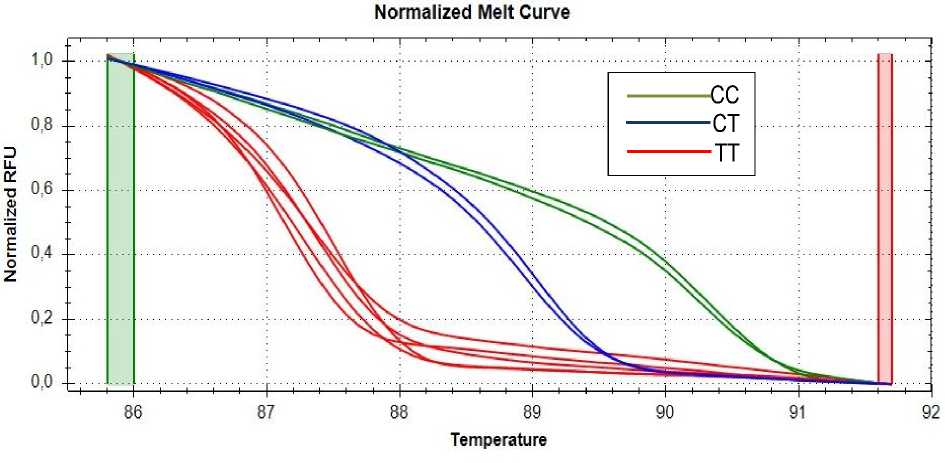
Результаты HRM-анализа в программе PrecisionMeltAnalysisТМsoftware
Характеристика популяционных частот аллелей С и Т CAPN1 среди чистопородного поголовья овец калмыцкой курдючной породы и ее помесей от скрещивания с породами шароле и дорпер приведена в таблице 1.
Частоты аллелей С и Т CAPN1 у овец калмыцкой курдючной породы и ее помесей от скрещивания с породами шароле и дорпер
Таблица 1
|
Аллель |
Бараны |
Овцематки |
Молодняк |
Баранчики |
Ярочки |
|
Породная принадлежность |
|||||
|
КК |
КК |
КК |
|||
|
С |
0,667±0,08 |
0,500±0,05 |
0,231±0,03 |
0,214±0,06 |
0,250±0,07 |
|
Т |
0,333±0,08 |
0,500±0,05 |
0,769±0,03 |
0,786±0,06 |
0,750±0,07 |
|
Ш |
КК |
½Шх×½КК |
|||
|
С |
0,750±0,22 |
0,500±0,05 |
0,344±0,03 |
0,417±0,08 |
0,300±0,05 |
|
Т |
0,250±0,22 |
0,500±0,05 |
0,656±0,03 |
0,583±0,08 |
0,700±0,05 |
|
Ш |
½ККх½Д |
½Ш×¼Д×¼КК |
|||
|
С |
0,750±0,22 |
0,550±0,02 |
0,540±0,02 |
0,500±0,06 |
0,559±0,03 |
|
Т |
0,250±0,22 |
0,450±0,02 |
0,460±0,02 |
0,500±0,06 |
0,441±0,03 |
Анализ данных показывает, что частоты аллелей С и Т различны у родителей и потомства как при чистопородном разведении, так и при скрещивании. Так, у баранов-производителей и овцематок калмыцкой курдючной частота аллеля С составила 0,667 и 0,500 соответственно, тогда как у чистопородного потомства наблюдалось повышение частоты аллеля Т до 0,769, при снижении встречаемости аллеля С до 0,231. Аналогичное изменение частоты встречаемости аллелей наблюдалось и в группе помесного молодняка при использовании породы шароле. Если у баранов и овцематок аллель Т выявлялся с частотой 0,250 и 0,500, то у потомства его концентрация повысилась до 0,656, тогда как аллель С у родителей выявлялся с частотой 0,750 и 0,500, а у потомства его распространение снизилось до 0,344. Меньшие изменения в частоте встречаемости аллелей отмечались в потомстве, полученном от помесных калмыцких курдючных с породой дорпер овцематок и баранов породы шароле: аллели С и Т выявлялись практически с той же частотой, что и у матерей. При этом следует отметить, что в сравнении с потомством других вариантов разведения в этой группе было несколько большее переопределение в пользу аллеля С, что определило его большую частоту.
Сравнивая частоты аллелей среди всех групп, можно заключить, что среди баранов-производителей обеих пород в гене CAPN1 чаще выявлялись носители аллеля С, тогда как среди чистопородных и помесных маток носительство аллелей С и Т было практически равным; среди молодняка – у чистопородных и двухпородных помесей преобладали особи с аллелем Т, тогда как трех породных – носители аллеля С.
Анализ распределения генотипов в исследованных вариантах скрещивания и чистопородного разведения включал оценку соответствия наблюдаемого распределения генотипов теоретически ожидаемому согласно уравнению Харди-Вайнберга (табл. 2).
Таблица 2
Распределение генотипов и оценка генетического равновесия (χ2) в CAPN1 у овец калмыцкой курдючной породы и ее помесей от скрещивания с породами шароле и дорпер, %
|
Генотип |
Бараны |
Овцематки |
Молодняк |
Баранчики |
Ярочки |
|||||
|
Породная принадлежность |
||||||||||
|
КК |
КК |
КК |
||||||||
|
Н |
О |
Н |
О |
Н |
О |
Н |
О |
Н |
О |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
|
СС |
33,3 |
44,4 |
40,0 |
25,0 |
15,4 |
5,3 |
14,3 |
4,6 |
16,7 |
6,3 |
|
СТ |
66,7 |
44,4 |
20,0 |
50,0 |
15,4 |
35,5 |
14,3 |
33,7 |
16,7 |
37,5 |
|
ТТ |
0,0 |
11,1 |
40,0 |
25,0 |
69,2 |
59,2 |
71,4 |
61,7 |
66,7 |
56,3 |
Окончание табл. 2
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
|
χ 2 |
0,31 |
2,05 |
2,00 |
0,42 |
0,30 |
|||||
|
Ш |
КК |
½Ш×½КК |
||||||||
|
СС |
50,0 |
56,3 |
40,0 |
25,0 |
25,0 |
11,8 |
33,3 |
17,4 |
20,0 |
9,0 |
|
СТ |
50,0 |
37,5 |
20,0 |
50,0 |
18,8 |
45,1 |
16,7 |
48,6 |
20,0 |
42,0 |
|
ТТ |
0,0 |
6,3 |
40,0 |
25,0 |
56,3 |
43,1 |
50,0 |
34,0 |
60,0 |
49,0 |
|
χ 2 |
1,33 |
2,05 |
3,66 |
0,99 |
1,16 |
|||||
|
Ш |
½ Д×½ КК |
½Ш×¼Д×¼КК |
||||||||
|
СС |
50,0 |
56,3 |
35,0 |
30,3 |
24,0 |
29,2 |
12,5 |
25,0 |
29,4 |
31,2 |
|
СТ |
50,0 |
37,5 |
40,0 |
49,5 |
60,0 |
49,7 |
75,0 |
50,0 |
52,9 |
49,3 |
|
ТТ |
0,0 |
6,3 |
25,0 |
20,3 |
16,0 |
21,2 |
12,5 |
25,0 |
17,6 |
19,5 |
|
χ2 |
1,33 |
0,28 |
0,55 |
0,81 |
0,02 |
|||||
Примечание : Н – наблюдаемые; О – теоретически ожидаемые значения; отклонение Н от О по закону Харди-Вайнберга значимо при χ2 ≥ 3,84.
Полученные данные не выявили ни в одной из групп достоверно значимого отклонения наблюдаемых генотипов от теоретически ожидаемых. Однако у баранов-производителей калмыцкой курдючной отмечалось некоторое увеличения доли гетерозигот (66,7 % наблюдаемой против 44,4 % ожидаемой), тогда как у овцематок это соотношение имело противоположный характер – снижение доли гетерозигот (20,0 % наблюдаемой против 50,0 % ожидаемой). У потомства отмечалось увеличение числа гетерозигот СТ и гомозигот ТТ (15,4 и 69,2 % против 35,5 и 59,2 % соответственно).
Сравнение полученных данных с результатами других исследований позволяет отметить, что расположенный между экзонами 5 и 6 полиморфизм, представляющий собой синонимичный переход Т/С в 44 нуклеотиде гена (AF309634.1) [12, 14], характеризуется разным
Таблица 3
Живая масса баранов-производителей и овцематок различных генотипов по гену CAPN1
|
Породная принадлежность |
По всем генотипам, кг |
Генотип |
|||
|
СС |
СТ |
ТТ |
|||
|
Бараны-производители |
КК |
89,3±1,1 |
91,0±0,6 |
88,0±1,4 |
– |
|
Ш |
80,3±1,1 |
– |
81,4 |
79,2 |
|
|
Овцематки |
КК |
62,3±0,32 |
62,3±1,2 |
62,4±1,3 |
62,0±1,1 |
|
½ Д ×½ КК |
62,9±0,31 |
62,8±0,5 |
63,0±0,7 |
62,8±0,6 |
|
Сопоставление живой массы разных генотипов показало, что бараны-производители калмыцкой курдючной породы с генотипом СС имели тенденцию к превышению над носителями генотипа СТ. Однако, ввиду небольшого соотношением генотипов СС, СТ, и ТТ у тех или иных пород. Так, генотип CC и аллель C являются наиболее частыми у трех египетских пород овец (барки, рахмани и оссими), при отсутствии генотипа ТТ у пород барки и оссими [15]. У овец породы бандур зарегистрированы два генотипа – СС и СТ с частотами 0,67 и 0,30 [16]. У курдских овец в этом локусе обнаружен незначительный избыток гетерозигот [17], тогда как у овец пород бандур [16] и зел [14], а также колумбийских креольских овец [18], наоборот, наблюдался их дефицит.
Анализ ассоциативной связи полиморфизма CAPN1 с живой массой и ее среднесуточным приростом у овец исследуемых групп был направлен на то, чтобы охарактеризовать особенности темпа роста чистопородного и помесного потомства разных генотипов (табл. 3).
размера выборки, для интерпретации данного наблюдения как закономерности требуется расширение исследования с привлечением большего числа животных. Среди овцематок разной породной принадлежности не установ- лено достоверных различий по живой массе между животными разных генотипов в гене CAPN1. По-видимому, значительно более жесткий отбор среди баранов калмыцкой курдючной породы привел к тому, что из разведения исключались носители генотипа ТТ, при этом животные с генотипом СТ уступали по живой массе носителям СС генотипа, что в определенной
степени свидетельствует о понижающем эффекте аллеля Т на живую массу баранов.
Показатели живой массы при рождении, в 4 месяца и среднесуточный прирост в этот период у чистопородного молодняка и от разных вариантов скрещивания, различных генотипов по гену CAPN1 представлены в таблице 4.
Живая масса и среднесуточный прирост у чистопородного молодняка и от разных вариантов скрещивания, различных генотипов по гену CAPN1
Таблица 4
|
s 1— о ф 1________ |
Породная принадлежность |
||||||||
|
КК×КК |
½ Ш ×½КК |
½Ш×¼ Д ×¼ КК |
|||||||
|
Живая масса, кг |
о ОФ П 5: >= О £ |
Живая масса, кг |
h о Ф П О £ |
Живая масса, кг |
Is о о. Ф П О £ |
||||
|
4 О |
4 мес. |
4 О |
4 мес. |
4 О |
4 мес. |
||||
|
Ба |
ранчики |
||||||||
|
СС |
4,8±0,3 |
42,2±0,5* |
307±4,7* |
3,7±0,4 |
42,0±1,4* |
314±7,8* |
4,3±0,2 |
41,4±0,6 |
304±7,2 |
|
СТ |
3,6±0,2 |
38,2±0,4 |
284±3,6 |
4,0±0,4 |
40,2±1,6 |
297±6,9 |
3,7±0,1 |
40,8±0,6 |
305±5,3 |
|
ТТ |
4,3±0,3 |
41,9±0,4 |
308±2,7 |
3,7±0,2 |
38,2±1,2 |
283±9,1 |
5,2±0,3 |
41,6±0,5 |
298±5,7 |
|
Ярочки |
|||||||||
|
СС |
4,0±0,2 |
40,0±0,7* |
295±6,5* |
3,9±0,1 |
37,6±1,4 |
279±9,0 |
3,5±0,2 |
38,5±0,6 |
287±4,2 |
|
СТ |
3,3±0,1 |
36,3±0,5 |
271±7,1 |
3,6±0,3 |
35,1±0,5 |
256±4,7 |
3,7±0,1 |
37,5±0,5 |
278±4,4 |
|
ТТ |
3,7±0,1 |
34,6±0,6 |
253±5,8 |
3,5±0,1 |
36,0±0,8 |
266±7,0 |
4,0±0,1 |
37,2±1,3 |
272±8,7 |
*p < 0,05.
Установлено, что независимо от породной принадлежности и пола живая масса молодняка при рождении разных генотипов по гену CAPN1 была схожей, тогда как в 4-месячном возрасте как баранчики, так и ярочки с генотипом СС имели достоверное превосходство над сверстниками других генотипов по живой массе и среднесуточному приросту. Так, у чистопородных баранчиков наибольшая разность была между животными генотипов СС и СТ, у помесей ½Ш×½КК – животных генотипов СС и ТТ, которая составила 10,4 и 8,1 %; 9,94 и 10,95 % (p < 0,05) соответственно. Среди ярочек наибольшие различия наблюдались при чистопородном разведении между животными генотипов СС и ТТ – 15,6 и 16,6 % (p < 0,05) соответственно. Следует отметить, что у ярочек-носителей генотипа СС во всех вариантах отмечена тенденция к большему уровню среднесуточного прироста в сравнении с животными, имеющими генотипы СТ и ТТ.
Таким образом, полученные данные позволяют заключить, что у молодняка овец в ранний период роста и развития генотип СС в гене CAPN1 ассоциирован с большей живой массой, тогда как присутствие аллеля Т снижает ее уровень. Можно предположить, что положительное влияние на прирост живой массы аллеля С в гомозиготном состоянии будет прослеживаться и в более поздние возрастные периоды роста ягнят, что станет обоснованием целесообразности отбора носителей генотипа СС для увеличения численности животных с потенциально более высокой мясной продуктивностью. Однако для подтверждения этого суждения необходимо проведение дальнейших исследований на большей по численности выборке животных.
Заключение. Анализ результатов генотипирования полиморфизма в гене CAPN1 выявил, что среди баранов-производителей калмыцкой курдючной породы и шароле чаще выявлялись носители аллеля С, среди чистопородных и по- месных маток носительство аллелей С и Т было практически равным; среди молодняка у КК и помесей ½Ш×½КК преобладали особи с аллелем Т, у помесей½Ш×¼Д×¼КК– носители аллеля С.
Популяционно-генетический анализ не выявил достоверных отличий наблюдаемых частот генотипов от теоретически ожидаемых при равновесии Харди-Вайнберга.
В 4-месячном возрасте как баранчики, так и ярочки с генотипом СС имели превосходство над сверстниками других генотипов по живой массе и среднесуточному приросту. Наибольшее превосходство выявлено при сравнении с животными генотипа СТ у чистопородных баранчиков и с генотипом ТТ у помесных животных ½Ш×½КК, которое соответственно составило 10,4 и 8,1 %; 9,94 и 10,95 % (p < 0,05). Среди ярочек наибольшие различия наблюдались при сравнении с особями генотипа ТТ при чистопородном разведении и составили 15,6 и 16,6 % (p < 0,05) соответственно.
Список литературы Ассоциация полиморфных вариантов гена CAPN1 с живой массой у овец калмыцкой курдючной породы и ее помесей с породами Шароле и Дорпер
- Состояние и тенденции в производстве мяса домашних животных в мире и России / А.И. Ерохин [и др.] // Овцы, козы, шерстяное дело. 2021. № 2. С. 20–22.
- Малышева Е.С., Бессонова Н.М. Оценка качественных характеристик баранины // Вестник Алтайского государственного аграрного университета. 2016. № 4 (138). С. 124–127.
- Генетические маркеры мясной продуктивности овец (Ovis aries L.). Сообщение I. Миостатин, кальпаин, кальпастатин / В.И. Тру-хачев [и др.] // Сельскохозяйственная биология. 2018. Т. 53, № 6. С. 1107–1119.
- Немова Н.Н., Лысенко Л.А., Канцерова Н.П. Протеиназы семейства кальпаинов. Структура и функции // Онтогенез. 2010. Т. 41, № 5. С. 381–389.
- The calpain system / D.E Goll [et al.] // Physio Rev., vol. 83, 2003, P. 731–801.
- Glading A., Lauffenburger D.A., Wells A. Cut-ting to the chase: calpain proteases in cell mo-tility // Trends Cell Biol. 2002. V. 12. P. 46–54.
- Chondrogianni N., Fragoulis E.G., Gonos E.S. Protein degradation during aging: the lyso-some, the calpain and the proteasome dependent cellular proteolytic systems // Biogerontology. 2002. V. 3. P. 121–123.
- Dear T.N., Boehm T. Diverse mRNA expres-sion patterns of the mouse calpain genes Capn5, Capn6, and Capn11 during develop-ment // Mech. Devel. 1999. V. 89. P. 201–209.
- Barnoy S., Maki M., Kosower N.S. Overex-pression of calpastatin inhibits L8 myoblast fu-sion // Biochem. Biophys. Res. Comm. 2005. V. 332. P. 697–701.
- Genetic polymorphism at MTNR1A, CAST and CAPN loci in Iranian Karakul sheep / F.E. Shahroudi [et al.] // Iranian Journal of Biotechnology, 2006, 4(2): 117–122.
- Variation in exon 10 of the ovine calpain 3 gene (CAPN3) and its association with meat yield in New Zealand Romney sheep / Q. Fang [et al.] // Meat Sci., 2013, 94(3): 388–390. DOI: 10.1016/j.meatsci.2013.03.015.
- Association between single nucleotide poly-morphism in ovine Calpain gene and growth performance in three Egyptian sheep breeds / K. Mahrous [et al.] // J. Genet. Eng. Biotech-nol. 2016. 14, 233–240.
- Polymorphism of Calpastatin, Calpain and myostatin genes in native Dalagh sheep in Iran / M.A. Azari [et al.] // Slovak Journal of Animal Science, 2012, 45(1): 1–6.
- Genetic variability of calpastatin and calpain genes in iranianZel sheep using PCR-RFLP and PCR-SSCP methods. Iran / E. Dehnavi [et al.] // J. Biotechnol. 2012. 10, 136–139.
- Михайлова М.Е., Романишко Е.Л. Определение полиморфизма гена эстрогенового рецептора ESR1-Pvu II, маркера плодовитости свиней, с помощью HRM-анализа // Молекулярная и прикладная генетика. 2014. Т. 17. С. 91–96.
- Genetic polymorphism of ovine calpain gene in bandur sheep / N.S. Kumar [et al.] // Interna-tional Journal of Science, Environment and Technology, 2015, 4(3): 804–812.
- Calpastatin polymorphism and its association with daily gain in Kurdi sheep / M.R. Nassiry [et al.] // Iran. J. Biotecnol., 2006, 4(3): 188–192.
- Montes D., Lenis C., Hernández D. Polymor-phisms of the calpain and calpastatin genes in two populations of Colombian creole sheep // Rev. MVZ. 2015. Córdoba 24, 7113–7118.
