Биохимический механизм автолитических процессов мышечной ткани рыб

Автор: Антипова Л.В., Дворянинова О.П., Черкесов А.З.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Пищевая биотехнология
Статья в выпуске: 2 (64), 2015 года.
Бесплатный доступ
Проведенные исследования позволили установить, что интенсивный распад мышечного гликогена приводит к резкому снижению величины рН мышечной ткани в кислую сторону, что в свою очередь сказывается на химическом составе и физико-коллоидной структуре белков, в результате чего: увеличивается устойчивость мяса рыбы к действию гнилостных микроорганизмов; снижается растворимость мышечных белков, уровень их гидратации, водосвязывающей способности; происходит набухание коллагена соединительной ткани; повышается активность катепсинов (оптимум рН 5,3), вызывающих гидролиз белков на более поздних стадиях автолиза; разрушается бикарбонатная система мышечной ткани с выделением углекислого газа; формируются предшественники вкуса и аромата мяса; активизируется процесс окисления липидов. В результате накопления молочной, фосфорной и других кислот в мясе рыбы увеличивается концентрация водородных ионов, результатом чего является снижение рН. Резко проявляющаяся кислая среда и наличие неорганического фосфора считается причиной распада актомиозинового комплекса на актин и миозин, который начинается после 8 часов хранения, т.е. наступает период расслабления мышечных волокон и период разрешения окоченения, а затем последняя стадия созревания мяса - глубокий автолиз. Таким образом, на основе классических представлений о биохимических изменениях мяса наземных животных и суммируя полученные данные по посмертным изменениям в мышечной ткани рыб, можно сделать вывод, что они имеют аналогичный характер закономерности по сравнению с мышечной тканью наземных животных, но главным их отличием является более высокая скорость протекания автолитических превращений. Это в свою очередь приводит к более быстрому изменению ФТС мяса рыб, которые являются определяющими показателями при разработке ассортиментных групп продуктов с учетом стадий автолиза в мясе.
Мышечная ткань, актомиозиновый комплекс, автолиз, созревание, прудовые рыбы, гликоген, актин, миозин
Короткий адрес: https://sciup.org/14040444
IDR: 14040444 | УДК: 597.554
Biochemical mechanism of autolytic processes of muscular tissue of fishes
The conducted researches allowed to establish that intensive disintegration of a muscular glycogen leads to sharp decrease in size рН muscular tissue in the sour party that in turn affects a chemical composition and physic-colloidal structure of proteins therefore: resistance of meat of fish to action of putrefactive microorganisms increases; solubility of muscle proteins, level of their hydration which is water connecting abilities decreases; there is a swelling of collagen of connecting fabric; activity of the cathepsin (an optimum рН 5,3) causing hydrolysis of proteins at later stages of an autolysis increases; the bicarbonate system of muscular tissue with release of carbon dioxide collapses; predecessors of taste and aroma of meat are formed; process of oxidation of lipids becomes more active. As a result of accumulation dairy, phosphoric and other acids in meat of fish concentration of hydrogen ions of that decrease рН is result increases. Sharply shown sour environment and availability of inorganic phosphorus is considered the reason of disintegration of an actin-myosin complex on actin and a myosin which begins after 8 hours of storage, i.e. there comes the period of relaxation of muscle fibers and the period of permission of an numbness, and then the last stage of maturing of meat - deep autolysis. Thus, on the basis of classical ideas of biochemical changes of meat of land animals and summarizing the obtained data on posthumous changes in muscular tissue of fishes, it is possible to draw a conclusion that they have similar nature of regularity in comparison with muscular tissue of land animals, but their main difference is higher speed of course of autolytic transformations. It in turn leads to faster change of FTS of meat of fishes who are the defining indicators when developing assortment groups of products taking into account stages of an autolysis in meat.
Текст научной статьи Биохимический механизм автолитических процессов мышечной ткани рыб
В основе посмертного окоченения мышечной ткани прудовых рыб лежат сложные физико-химические процессы, протекающие главным образом в мышечном волокне. Главную роль в мышечном сокращении играют миофибриллы, которые из веществ, входящих в состав саркоплазмы, используют необходимую энергию для выполнения своих функций. Источником энергии в данном случае служат только биохимические процессы ферментативного распада (гидролиз, фосфоролиз) некоторых веществ, в частности, нуклеозидфосфатов гликогена, находящихся главным образом в саркоплазме.
В мышечной ткани рыбы основная часть энергии, необходимой для сокращения мышц, выделяется в результате расщепления АТФ. По внешнему проявлению и химизму процессов посмертное окоченение практически идентично двигательному сокращению живой мышцы. Молекулярный механизм сокращения мышц основан на взаимодействии миозиновых и актиновых филламентов.
Наступление посмертного окоченения связано с тем, что в мышцах живой и совершенно свежей уснувшей рыбы отмечается достаточно высокое содержание АТФ, которая удерживает актин и миозин в диссоциированном состоянии. При работе живого организма, так же как и после его смерти, происходит распад находящихся в мышцах АТФ с образованием АДФ и фосфорной кислоты. Это происходит под влиянием АТФазной активности миозина. Однако в живом организме полного распада АТФ не происходит и образующийся АДФ снова восстанавливается до АТФ.
В мертвом организме, в отличие от живого, процесс идет преимущественно в направлении автолитического распада энергетических веществ и по мере расходования АТФ, креатин фосфата, гликогена происходит ассоциация актина и миозина с образованием вязкого актомиозина, придающего мышце жесткость (окоченение) (Антипова, Жеребцов, 1991).
В основе автолитических превращений мяса рыбы лежат изменения углеводной системы, системы ресинтеза АТФ и состояния миофибриллярных белков, входящих в систему сокращения.
Следует отметить, что пусковым механизмом посмертного окоченения является распад мышечного гликогена, который катализируют 11 ферментов, при этом распад идет по пути фосфоролиза и дальнейшего анаэробного окисления до образования пировиноградной и молочной кислот, поскольку аэробный путь превращения затухает, вследствие прекращения доступа кислорода в клетки. Затем начинается самораспад тканей под действием гидролитических ферментов. Наступает автолиз.
Уменьшение количества АТФ в мышцах сопровождается активизацией функциональных групп актина и миозина, в результате чего происходит перемещение нитей актина по поверхности миозина с образованием нерастворимого АКМ, придающего мышце жесткость, т.е. окоченение (рисунок 1).
4 им
8®$tit Ж^ ЖЖ~ «И9Г
ADP
ADP н------^------>|
Рисунок 1. Схема перемещения нитей актина по поверхности миозина с образованием нерастворимого актомиозинового комплекса (по данным
Тихонова А.Н., 2003)
Целью работы являлось исследование закономерности изменения биохимических и физико-химических превращений при хранении и переработке прудовых рыб.
Для определения глюкозы, АТФ применялись оригинальные методы [2], при этом определение гликогена и актомиозинового комплекса проводили методом ВЭЖХ [3], определение ПВК согласно колориметрическому методу по Умбрайту [2, 3].
Проведенные исследования позволили установить, что интенсивный распад мышечного гликогена (рисунок 2) приводит к резкому снижению величины рН мышечной ткани в кислую сторону, что в свою очередь сказывается на химическом составе и физико-коллоидной структуре белков в результате чего:
-
- увеличивается устойчивость мяса рыбы к действию гнилостных микроорганизмов;
-
- снижается растворимость мышечных белков, уровень их гидратации, водосвязывающей способности;
-
- происходит набухание коллагена соединительной ткани;
-
- повышается активность катепсинов (оптимум рН 5,3), вызывающих гидролиз белков на более поздних стадиях автолиза;
-
- разрушается бикарбонатная система мышечной ткани с выделением углекислого газа;
-
- формируются предшественники вкуса и аромата мяса;
- активизируется процесс окисления ли-
- мышечной ткани прудовых рыб в процессе
хранения
Молекулярная основа процесса такова: поперечные связи между филаментами актина и миозина сохраняются и не могут разорваться из-за отсутствия АТФ. Активный транспорт Cа2+ в саркоплазматический ретикулум оказывается невозможным, так же как и переход миозина в активированное состояние
(конформация миозин – АДФ • Фн). По этой причине продукты гидролиза АТФ (АДФ и Фн) удаляются не вследствие образования комплекса актин - миозин – АДФ • Фн, а диффузией. При этом не совершаются соответствующие конформационные переходы в рабочих элементах клетки, а, следовательно, взаимодействие актинмиозин остается. Актомиозиновый комплекс можно разрушить, приложив внешнюю силу, но восстановить потом нельзя.
В первый период интенсивность гидролиза сравнительно невелика, а в дальнейшем гидролитический распад гликогена резко повышается. Это объясняется присутствием в мышечных волокнах двух ферментативных механизмов гидролитического расщепления гликогена: амилаза и нейтральные олигоглюкозидазы, содержащиеся в матриксе саркоплазме; γ-амилаза и кислые олигоглюкозидазы, содержащиеся в лизосомах, высвобождение из которых и обуславливает рост гидролитического распада. При этом минимальное содержание гликогена наблюдается к 24 часам хранения.
Это связано с тем, что в мышцах живой рыбы отмечается достаточно высокое содержание АТФ, которая удерживает актин и миозин в диссоциированном состоянии, при этом происходит распад находящейся в мышцах АТФ с образованием АДФ, АМФ и фосфорной кислоты под влиянием АТФазной активности миозина.
В уснувшей же рыбе, в отличие от живой, вследствие прекращения доступа кислорода, регулирования обмена веществ и энергии в тканях, обратимые жизненные процессы становятся не обратимыми, при этом распад клеточных веществ превалирует над синтезом. При этом процесс идет преимущественно в направлении автолитического распада энергетических веществ: АТФ, креатин фосфата и гликогена (рисунок 3).
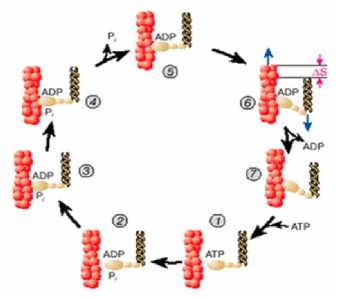
Рисунок 3. Рабочий цикл актомиозинового комплекса (по данным Тихонова А.Н., 2003)
В нашем случае закономерности изменения содержания гликогена имеют общий характер для всех исследуемых видов рыб, что подтверждает классические представления о механизме автолитических превращений [1, 2].
В связи с отсутствием поступления кислорода в организм (рисунок 4) ресинтез гликогена в мясе рыбы после убоя идти не может, и начинается его анаэробный распад, который протекает по пути фосфоролиза и амилолиза с образованием молочной кислоты и глюкозы, а также глюкозо-1-фосфата, глюкозо-6-фосфата и др.
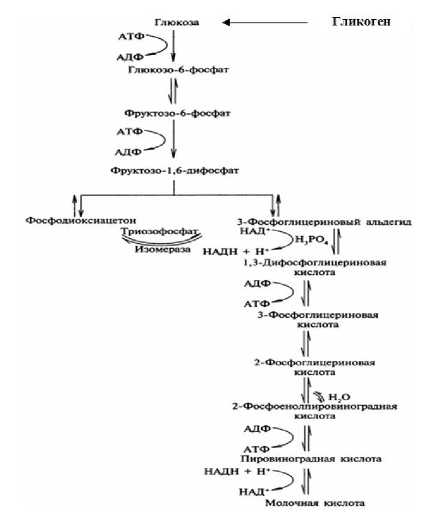
Рисунок 4. Схема распада гликогена по пути гликогенолиза
Совершается незначительный гидролитический распад гликогена за счет различных гликозидаз саркоплазмы.
О протекании биохимических процессов возможно судить по изменению в содержании продуктов распада гликогена, в частности, по образованию глюкозы и пирувата (рисунок 5) [3].
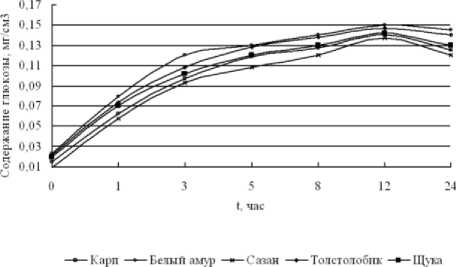
Рисунок 5. Динамика изменения содержания глюкозы в мышечной ткани прудовых рыб в
В дальнейшем глюкоза вовлекается в цикл трикарбоновых кислот, с образованием кислот, способствующих снижению рН среды в мышечной ткани, что является этапом начала действия тканевых ферментов - катепсинов [4].
На рисунке 5 видно, что содержание глюкозы для всех видов исследуемых рыб увеличивается в процессе хранения. Менее заметное увеличение глюкозы происходит в первые часы хранения (от 0 до 5 часов), о чем свидетельствует активный распаде гликогена по пути гликогенолиза.
Пиком накопления глюкозы является 12 часов хранения для всех видов рыб и составляет: 0,14 мг/см3 для карпа, 0,137 мг/см3 для сазана, 0,142 мг/см3 для щуки, 0,147 мг/см3 для толстолобика, 0,15 мг/см3 для белого амура.
Результаты исследований (рисунок 6) показывают, что рост содержания пирувата характерен для всех исследуемых рыб. По сравнению с рыбой, не подвергавшейся хранению, за 24 часа содержание ПВК увеличивается в мясе карпа от 182,3 до 312,4 мг/100 г, для толстолобика – от 194,5 до 320,0 мг/100 г, для белого амура от 201,3 до 332,4 мг/100 г, для сазана от 171,9 до 310,5 мг/100 г и для щуки от 187,1 до 314,8 мг/100 г. Изменения количества пировиноградной кислоты и глюкозы у всех исследуемых рыб носят аналогичный характер. Однако у толстолобика и белого амура выше, чем у щуки, карпа и сазана, что свидетельствует о более активных автолитических превращениях в мясе этих видов рыб [4, 5].
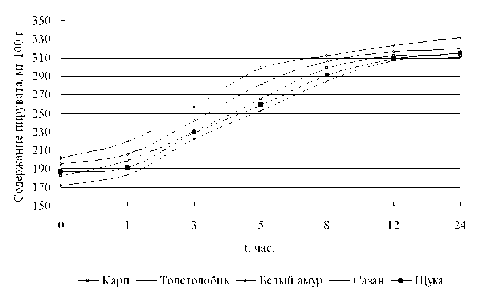
Рисунок 6. Динамика изменения содержания ПВК в процессе хранения прудовых рыб
На начальной стадии автолиза важное значение имеет уровень содержания в мясе рыбы энергоёмкой АТФ, из-за распада которой происходит ферментативный процесс расщепления гликозидных связей (рисунок 7).
процессе хранения
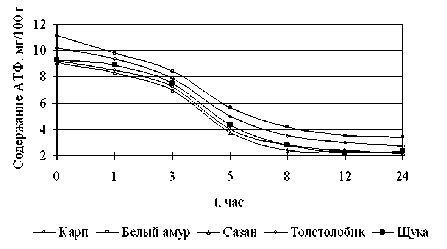
Рисунок 7. Изменение содержания АТФ в мясе прудовых рыб в процессе хранения
Одновременно процесс распада аденаз-интрифосфата обеспечивает сокращение мио-фибриллярных белков. Динамика изменения содержания АТФ для мяса рыбы в послеубой-ный период характерно непрерывное снижение концентрации АТФ, вследствие уменьшения запасов которого не хватает энергии для восстановления состояния релаксации сократившихся волокон (Антипова, Алехина, 2011).
При этом распад АТФ до АДФ, АМФ приводит к повышению кислотности среды в мясе рыб, что предопределяет ФТС мяса рыбы. При этом в образовавшейся кислой среде происходит частичное накопление неорганического фосфора. Кислая среда и наличие неорганического фосфора считается причиной диссоциации актомиозинового комплекса на актин и миозин. Распад этого комплекса снимает явления окоченения и жесткости мяса. Следовательно, фазу окоченения от других фаз обособить нельзя и ее необходимо считать одним из этапов процесса созревания мяса рыбы.
Характер кривых на рисунке 7 говорит о том, что распад АТФ в мясе рыбы происходит в первые часы хранения. Минимальное его содержание приходится на 24 часа: для щуки -2,314, карпа - 2,2, белого амура - 3,416, толстолобика - 2,713, сазана - 2,103 мг/100 г [3, 4].
Исследования закономерностей изменения содержания актомиозинового комплекса (рисунок 8) в мышцах всех исследуемых видов рыб в процессе хранения имеют аналогичный характер.
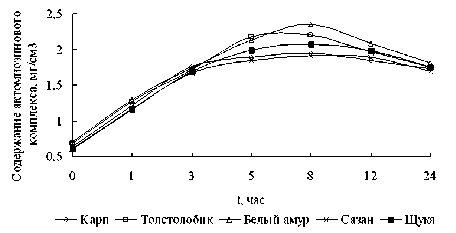
Рисунок 8. Динамика изменения содержания актомиозинового комплекса в мясе прудовых рыб в процессе хранения
Затвердение мышечной ткани наблюдается в первые часы хранения и обусловлено образованием из белков актина и миозина нерастворимого актомиозинового комплекса. Наивысший уровень содержания актомиозина отмечался в первые 3-5 часов, что свидетельствует о разной скорости протекающих биохимических процессов, и связано с уровнем активности ферментных систем. Максимум накопления актомиозинового комплекса приходится на 8 ч хранения для всех исследуемых видов рыб [2, 3].
Кислая среда усиливает мышечное окоченение вследствие не поступления в мышечную ткань рыбы кислорода, торможения окислительных процессов, накопления избытка молочной и фосфорной кислот. Реакция среды при этом снижается с 7,26 до 6,02. Также при накоплении молочной кислоты происходит коагуляцией белка. При этом АМК теряет свою растворимость, белки стабилизируются, а кальций выпадает из коллоидов белка и переходит в мясной сок рыбы. Полное окоченение рыбы наступает к 8 часам хранения. Далее по мере увеличения концентрации молочной кислоты и коагуляции белков происходит распад АМК. Свернувшиеся белки теряют свои коллоидные свойства, становятся неспособными удерживать воду и в известной степени лишаются своей дисперсной среды (воды).
В результате накопления молочной, фосфорной и других кислот в мясе рыбы увеличивается концентрация водородных ионов, результатом чего является снижение рН. Резко проявляющаяся кислая среда и наличие неорганического фосфора считается причиной распада актомиозинового комплекса на актин и миозин [4], который начинается после 8 часов хранения, т.е. наступает период расслабления мышечных волокон и период разрешения окоченения, а затем последняя стадия созревания мяса – глубокий автолиз [3, 4].
Таким образом, на основе классических представлений о биохимических изменениях мяса наземных животных и суммируя полученные данные по посмертным изменениям в мышечной ткани рыб, можно сделать вывод, что они имеют аналогичный характер закономерности по сравнению с мышечной тканью наземных животных, но главным их отличием является более высокая скорость протекания автолитических превращений. Это в свою очередь приводит к более быстрому изменению ФТС мяса рыб, которые являются определяющими показателями при разработке ассортиментных групп продуктов с учетом стадий автолиза в мясе.
Вестник ВГУИТ, №2, 2015
Список литературы Биохимический механизм автолитических процессов мышечной ткани рыб
- Буй С.Д., Мукатова М.Д. Способ выделения ферментного препарата из внутренностей прудовых видов рыб//Современные достижения биотехнологии: сб. материалов междунар. науч.-практ. конф; СевКавГТУ. Ставрополь, 2011. С. 440-443.
- Антипова Л.В., Алехина А.В. Автолитические превращения в мясе прудовых рыб//3-я конференция молодых ученых и специалистов институтов отделения «Хранения и переработки сельскохозяйственной продукции» Россельхозакадемии «Обеспечение качества и безопасности продукции агропромышленного комплекса в современных социально-экономических условиях». М., 2009. С. 47-51.
- Дворянинова О.П., Алехина А.В., Сторублевцев С.А. Получение и исследование свойств ферментного комплекса мяса пресноводного карпа//Известия высших учебных заведений. Пищевая технология. 2010. №4. С. 13-15.
- Mills D. J. Underreported and undervalued: Small-scale fisheries in the developing world scale fisheries management: frameworks and approaches for the developing world. 2011. Р. 1-15.
- Mora C. Management effectiveness of the world’s marine fisheries//PLoS Biology. 2009. № 7(6). Р. 12-19.
