Биологические особенности свиней, трансгенных по гену гормона роста человека

Автор: Эрнст Л.К., Зиновьева Н.А., Волкова Н.А., Филимонов А.Ю., Ралков И.А., Гусев И.В., Брем Г.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биология трансгенных животных
Статья в выпуске: 2 т.48, 2013 года.
Бесплатный доступ
Показано повышение экспрессии гормона роста у свиней, трансгенных по генной конструкции WAP-hGH генераций F 3 (холостые и лактирующие свиноматки) и F 5 (молодняк в возрасте 5 и 9 мес). Установлены изменения в некоторых биохимических показателях сыворотки крови трансгенных свиней. Обнаружено изменение корреляционных зависимостей между биохимическими показателями крови трансгенных и контрольных свиней.
Трансгенные свиньи, соматотропин, биохимические показатели, корреляции
Короткий адрес: https://sciup.org/142133391
IDR: 142133391 | УДК: 636.4:636.064:[573.6.086.83+577.21]:577.175.322
Biological characteristics of pigs transgenic for human growth hormone
The increase of growth hormone expression in pigs transgenic for WAP-hGH gene construct of F 3 (non-pregnant and lactating sows) and F 5 generations (young gilts in age of 5 and 9 months) was observed. The alteration of some biochemical characteristics of blood serum in transgenic pigs comparing to control animals was detected. The differences in correlation relationships between some biochemical characteristics of transgenic and control pigs were established.
Текст научной статьи Биологические особенности свиней, трансгенных по гену гормона роста человека
К приоритетным областям развития биотехнологий в животноводстве относится создание трансгенных форм животных, обладающих заданными продуктивными и биологическими свойствами или продуцирующих биологически активные вещества (1, 2). Для обеспечения направленного синтеза рекомбинантных белков в молочной железе в генно-инженерные конструкции включают регуляторные элементы генов белков молока — к -казеина, в -лактоглобулина, а -лактальбумина, кислого сывороточного белка (WAP) (3), при этом в ряде исследований показана неспецифическая экспрессия генных конструкций, обусловленная промотором WAP. Так, C.W. Pittius с соавт. (4) обнаружили мРНК тканевого плазминогенного активатора человека в языке, почках и слюнных железах трансгенных мышей. Экспрессия соматотропина (СТГ) человека, контролируемая промотором WAP мыши длиной 2,6 kb, выявлялась в сыворотке крови мышей в период лактации (5). По данным W.H. G u enzburg с соавт. (6), у 40 % трансгенных мышей, интегрировавших генную конструкцию СТГ человека под контролем промотора WAP длиной 2,4 kb, специфическую мРНК находили в слюнных железах и семенниках. У всех трансгенных мышей трансгенный белок синтезировался в клетках головного мозга. Причина неспецифической экспрессии генов, введенных в геном под контролем этого регуляторного элемента, заключается в неспецифичности экспрессии эндогенного WAP.
Несмотря на многочисленность и многообразие исследований в области трансгенеза, многие факторы, определяющие эффективность экспрессии чужеродных генов в клетках организма, остаются недостаточно изученными. Уже на ранних этапах исследований стало ясно, что взаимодействие интродуцированной конструкции и генома-хозяина не может быть сведено к простому суммированию различных генетических задатков.
Поскольку интеграция чужеродных генов по своей природе сходна мутации (инсерции), следует ожидать проявления в организме трансгенных животных целого спектра непредсказуемых событий модификационной и мутационной природы, так как каждая мутация захватывает не изолированный процесс для отдельного признака, а известную область процессов индивидуального развития (7). Подобное предположение было подтверждено в работах, выполненных на разных трансгенных животных (8-11). Следовательно, оценка влияния трансгена на организм исключительно актуальна как для биотехнологии, так и для селекции.
Мы изучили экспрессию введенных генов и ее взаимосвязи с физиолого-морфологическими признаками у трансгенных свиней 3- й и 5-й генераций.
Методика . Объектом исследований были свиньи, трансгенные по генной конструкции СТГ человека под контролем промотора WAP мыши (WAP-hGH). Для изучения их биологических и хозяйственно полезных признаков в условиях физиологического двора Всероссийского НИИ животноводства формировали группы по принципу одногнездовых аналогов.
Содержание СТГ в сыворотке крови определяли у свиней генераций F3 (холостые и лактирующие свиноматки) и F5 (молодняк в возрасте 5 и 9 мес) иммуноферментным методом на иммуноридере Dinatech («Dynex Technologies», США), используя набор С-kit («Sigma-Aldrich», США) согласно рекомендациям производителя. Экспрессию СТГ в органах и тканях оценивали методом иммуногистохимии с помощью набора ABC-kit («VectorLabs», Великобритания) в соответствии с рекомендациями производителя. В качестве первых антител применяли антитела, специфичные к СТГ человека («Sigma-Aldrich», США). Продукт иммуногистохимической реакции детектировали с использованием 3,3-диаминобензидина тетрахлората (ДАБ) («VectorLabs», Великобритания). Биохимические показатели крови исследовали у свиней генерации F5 в возрасте 7 мес, применяя общепринятые методики (12, 13) .
Полученные показатели обрабатывали биометрическими методами для малых выборок (14).
Результаты . В крови холостых и лактирующих трансгенных свиноматок в F3 содержание СТГ повышалось в 3,30-3,50 раза по сравнению с показателями у контрольных животных. У молодняка F5 в возрасте 5 мес количество СТГ увеличивалось в 2,25 раза, в возрасте 9 мес — в 3,77 раза относительно контроля. Следует отметить, что если у молодняка из контрольной группы различий в концентрации СТГ в разные возрастные периоды выявлено не было, то в группе трансгенных свиней происходил рост концентрации СТГ в 2,0 раза (табл. 1).
1. Содержание соматотропина (СТГ, нг/мл) в крови трансгенных (опыт) и нетрансгенных (контроль) свиноматок (F3) и молодняка (F 5 ) разного возраста ( X ± х )
|
Группа |
| Опыт |
| Контроль |
|
Лактирующие свиноматки |
n = 10 29,00±1,7а , б |
n = 10 8,30±1,9б |
|
Холостые свиноматки |
n = 10 24,00±1,2а , в |
n = 10 7,30±0,8в |
|
Молодняк на откорме: |
||
|
в возрасте 5 мес |
n = 3 1,53±0,2 |
n = 3 0,68±0,3 |
|
в возрасте 9 мес |
n = 3 3,10±0,9 |
n = 3 0,65±0,3 |
Примечание. Использованная генная конструкция — WAP-hGH (ген соматотропного гормона чело века под контролем WAP-промотора мыши). ар < 0,05, б, вр < 0,01 (различия достоверны).
сердце, печени, почках, легких и мышцах у этих животных экспрессию СТГ не наблюдали. Таким образом, трансгенные животные характеризовались более высоким содержанием СТГ в органах и тканях, при этом наиболее активное продуцирование СТГ происходило у трансгенных лакти-рующих свиноматок. Наличие СТГ в слюнной и поджелудочной железах и гипофизе свидетельствует о неспецифической экспрессии, обусловленной промотором WAP. В целом полученные данные можно рассматривать как указание на преимущество трансгенных свиней над контрольными по активности анаболических процессов.
У молодняка свиней в F 5 содержание продуктов метаболизма в сыворотке крови у трансгенных и контрольных особей находилось в пределах физиологической нормы. Среди наиболее заметных следует отметить
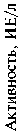
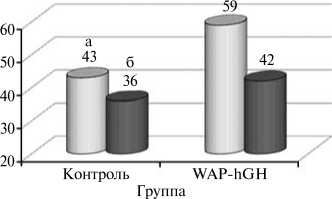
Рис. 1. Активность аланинаминотранферазы (а) и аспартатаминотрансферазы (б) в крови у контрольных и трансгенных свиней генерации F5. Использованная генная конструкция — WAP-hGH (ген соматотропного гормона человека под контролем WAP-промотора мыши).
наблюдаемые изменения в активности аминотрансфераз — аланинаминотрансферазы и аспартатамино -трансферазы (АлАТ и АсАТ). У трансгенных особей концентрация АлАТ была на 37,2 % выше, чем у аналогов из контрольной группы. Активность АлАТ положительно коррелировала с концентрацией глюкозы в сыворотке крови, которая у трансгенных свиней была на 8,8 % выше, чем у аналогов из контрольной группы. Активность АсАТ в сыворотке крови также имела более высокое значение (на 16,7 %) у особей из опытной группы по сравнению с контрольными (рис. 1). Тенденцию к росту активности АсАТ у трансгенных свиней можно рассматривать как указание на интенсификацию белкового и энергетического обмена в печени и мышечной ткани этих животных.
СТГ — один из регуляторов липидного обмена, и его повышенная продукция усиливает перемещение жира с периферии в печень. Учитывая различия в содержании СТГ в крови у контрольных и трансгенных свиней, у последних можно ожидать изменений в липидном обмене. Результаты анализа содержания общих липидов и их классов в сыворотке крови у трансгенных и контрольных особей в F5 показали (табл. 2), что в обеих группах оно находилось в пределах физиологической нормы. При этом в группе трансгенных особей по общим липидам показатели были на 5,6 % выше, чем в контроле, за счет повышения концентрации эфирного холестерола.
2. Содержание общих липидов и классов липидов в сыворотке крови у трансгенного (опыт) и нетрансгенного (контроль) молодняка свиней на откорме в
F 5 ( X ± х )
|
Показатель |
Контроль ( n = 6) |
Опыт ( n = 3) |
Разница «опыт — контроль» |
Опыт к контролю, % |
|
Общие липиды, г% В том числе по классам, %: |
0,40±0,01 |
0,38±0,02 |
0,02 |
5,6 |
|
фосфолипиды |
11,9±0,2 |
10,5±0,2 |
- 1,50 |
- 12,5 |
|
мобильные + депонированные глицеролы |
2,2±0,2 |
1,5±0,1 |
- 0,70 |
- 30,9 |
|
холестерол |
3,6±0,3 |
2,4±0,1 |
- 1,20 |
- 32,0 |
|
свободный холестерол |
37,7±0,3 |
44,4±0,3 |
6,80 |
17,9 |
|
триглицеролы |
38,3±0,2 |
35,6±0,0 |
- 2,70 |
- 6,9 |
|
НЭЖК |
5,9±0,2 |
5,4±0,1 |
- 0,50 |
- 8,5 |
Примечание. НЭЖК — неэтерифицированные жирные кислоты. Использованная генная конструк ция — WAP-hGH (ген соматотропного гормона человека под контролем WAP-промотора мыши).
Необходимо отметить тенденцию к снижению процентного содержания всех классов липидов (за исключением эфирного холестерола) у трансгенных животных по сравнению с контролем, что, возможно, связано у них с повышенной функциональной активностью СТГ, который способствует перемещению липидов с периферии в печень, где происходит активный гидролитический распад с образованием глицерина и жирных кислот и использование их в качестве энергетического материала.
Следует также обратить внимание на увеличение количества поли-ненасыщенных жирных кислот — линолевой, линоленовой и арахидоновой в крови трансгенных свиней. В частности, на долю этих жирных кислот у контрольных особей суммарно приходилось 7,90 %, в то время как у трансгенных показатель был на 1,96 % выше. Суммарное увеличение происходило в результате повышения содержания каждой из вышеназван ных жирных кислот (рис. 2).
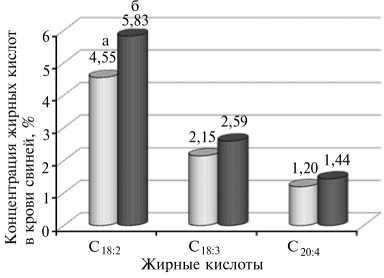
Рис. 2. Содержание полиненасыщенных жирных кислот в сыворотке крови у контрольных и трансгенных свиней генерации F5. С18:2 — линолевая, С18:3 — линоленовая, С20:4 — арахидоновая. Использованная генная конструкция — WAP-hGH (ген соматотропного гормона человека под контролем WAP-промотора мыши).
Противоречивость данных по фенотипу трансгенных животных подтверждает, что проблема регуляции функций организма посредством трансгенеза очень сложна, так как затрагивает многие регулирующие обмен веществ механизмы, воздействие которых при трансгенезе в конечном итоге определяет изменения в фенотипе.
В этом аспекте следует обратиться к трудам выдающегося исследователя И.И. Шмальгаузена, который утверждал: «Наследственная стойкость организма объясняется сложностью системы морфологических связей, объединяющих все части развивающегося организма в одно целое, а не стойкостью наслед ственного вещества, то есть генов».
Как уже отмечалось выше, интеграцию генов в геном можно рассматривать как разновидность мутации (инсерции). По представлению академика И.И. Шмальгаузена, любые изменения в геноме (мутации) и их воздействие на фенотип сглаживаются наличием хорошо развитых у высших животных морфологических корреляций. И.И. Шмальгаузен писал: «В процессе эволюции организм изменяется и эволюционирует как единое целое. Уже отдельные мутации имеют, благодаря связанности процессов индивидуального развития, характер изменений всего организма в целом». По мнению И.И. Шмальгаузена, мутации никогда не проявляются в из менениях отдельных признаков, а всегда проявляются в виде изменения корреляционных отношений, причем существующие корреляции часто теряются, а взамен этого устанавливаются совершенно новые.
Проведенные нами исследования взаимозависимостей между некоторыми биохимическими показателями в опытных и контрольных группах свиней показали, что в организме в результате интеграции генной конструкции существенно изменилась не только продукция СТГ, но и сложившиеся корреляции (табл. 3). Как следует из представленных в этой таблице данных, из 13 пар признаков в шести случаях коэффициент корреляции для трансгенных и контрольных особей имел отрицательное значение.
Например, если коэффициент корреляции между содержанием креатинина и креатинкиназы у трансгенных свиней равнялся - 0,64, то у контрольных — +0,58. Корреляция между содержанием глюкозы и инсулина составляла соответственно +0,49 и - 0,13. Даже если знак коэффициента корреляции был одинаковым у животных обеих групп, в большинстве случаев величина последнего оказалась существенно меньшей у трансгенных животных по сравнению с контрольными аналогами. Так, по содержанию общего белка и мочевины в крови коэффициент корреляции у трансгенных свиней составлял - 0,46, у контрольных — - 0,83, по концентрации глюкозы и триглицеридов — соответственно - 0,66 и - 0,99, амилазы и глюкозы — соответственно +0,03 и +0,52.
3. Корреляционные зависимости между биохимическими показателями крови у трангсенных (опыт) и нетрансгенных (контроль) свиней (генерации F 3 и F 5 )
|
Пара сравниваемых показателей |
Коэффициент корреляции r |
|
|
опыт |
контроль |
|
|
Креатинин—креатинкиназа |
- 0,64 |
+0,58 |
|
Общий белок—мочевина |
- 0,46 |
- 0,83 |
|
Глюкоза—инсулин |
+0,43 |
- 0,13 |
|
Глюкоза—триглицериды |
- 0,66 |
- 0,99 |
|
АсАТ—общий белок |
+0,50 |
+0,50 |
|
АлАТ—общий белок |
+0,51 |
+0,21 |
|
Тироксин—мочевина |
- 0,66 |
- 0,78 |
|
Тироксин—инсулин |
- 0,39 |
+0,02 |
|
Амилаза—глюкоза |
+0,03 |
+0,52 |
|
Трийодтиронин—тироксин |
+0,10 |
- 0,76 |
|
Лактатдегидрокиназа—холестерин |
- 0,87 |
+0,33 |
|
Кортизол—мочевина |
+0,01 |
+0,88 |
|
Лактатдегидрокиназа—молочная кислота |
+0,39 |
- 0,07 |
|
Примечание. АсАТ — аспартатаминотрансфераза, АлАТ - |
- аланинаминотрансфераза. |
В целом можно отметить, что интеграция генной конструкции WAP-hGH приводит не только к модификации ряда фенотипических признаков, но одновременно существенно изменяет направление и величину многих корреляций в организме животных. Это служит определенным преимуществом для селекции, так как сложившиеся связи способствуют стабилизации организма и препятствуют его изменению.
Из полученных нами результатов становится очевидным, что высокий уровень СТГ в крови трансгенных свиней не может быть полностью реализован в нормальном организме, так как все другие механизмы участия СТГ в метаболизме оказываются неизменными. Так, возможно противоречие между высоким содержанием СТГ и количеством рецепторов этого гормона.
Представленные данные подтверждают разработанную И.И. Шмаль-гаузеном теорию организма как целостной системы в индивидуальном и историческом развитии. «Так как все органы животного образуют единую систему, части которой зависят друг от друга и действуют и противодействуют одна по отношению к другой, никакое изменение не может обнаружиться в одной части без того, чтобы не вызвать соответствующие изменения во всех остальных частях».
Итак, трансгенные животные характеризуются перестройкой сложившихся коррелятивных связей, которые, несомненно, оказывают влияние на процессы развития особей. Все это свидетельствует о том, что интеграция и последующая экспрессия генов существенным образом меняет характер обмена веществ, затрагивает величину важнейших биохимических показателей. Разумеется, сейчас еще трудно определить значение каждого из этих факторов для развития хозяйственно ценных признаков, это будет предметом дальнейших исследований.
