Биологическое разнообразие и модели эволюции

Автор: Подвальный С.Л., Васильев Е.М.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Информационные технологии, моделирование и управление
Статья в выпуске: 3 (69), 2016 года.
Бесплатный доступ
Обсуждается эволюционное значение биологического разнообразия, системообразующее значение которого в ноосфере Земли закреплено конвенцией ООН. Рассмотрены примеры и роль этого разнообразия на разных уровнях организации жизни. На уровне отдельных организмов указанное разнообразие проявляется в дифференциации и разделении физиологически важных функций, значительно расширяющих экологическую нишу тех особей, у которых такое разделение наиболее совершенно. Однако организменный уровень биологического разнообразия непригоден для построения эволюционных моделей, так как возможность реализации процессов генетической наследственности и изменчивости возникает начиная с минимальной структурной единицы живого мира, которой является популяция. Отмечено, что накопление генофонда, необходимого для эволюции вида, возможно только в достаточно больших популяциях, в которых общая скорость мутационных изменений не уступает скорости вариаций условий внешней среды. Показано, что известные формальные модели видовой эволюции, построенные на основе теоремы Фишера о влиянии генной дисперсии на приспособленность особей, не отвечают реальным условиям существования видов в силу принятого конечного и постоянного числа генотипов в популяции. На экосистемном уровне организации жизни основную роль играет таксономическое разнообразие, которое обеспечивает непрерывность трофической цепи в системе при возникновении неблагоприятных условий для развития отдельных таксонов. Кроме того, существенное стабилизирующее значение для прогрессивной эволюции экосистемы имеют её многоуровневое, иерархическое строение и замкнутый круговорот вещества и энергии. Модели эволюции систем на базе уравнений Лотки-Вольтерры, описывающих взаимодействие элементов разомкнутой экосистемы, в недостаточной мере отражают роль биоразнообразия в эволюционных процессах. Сформулированы требования к таким моделям, учитывающим баланс массы в системе; её трофическую структуру; накопительные (буферные) свойства каждого уровня экосистемы; множественность взаимосвязей между уровнями экосистемы; ограничивающее влияние внутривидовой и межвидовой конкуренции. Перспективным в этом плане является привлечение идей и математического аппарата биокибернетики.
Биологическое разнообразие в природе, модели эволюции
Короткий адрес: https://sciup.org/140229572
IDR: 140229572 | DOI: 10.20914/2310-1202-2016-3-123-127
Biodiversity and models of evolution
Summary. The paper discusses the evolutionary impact of biodiversity, the backbone of noosphere, which status has been fixed by a UN convention. The examples and role of such diversity are considered the various levels of life arrangement. On the level of standalone organisms, the diversity in question manifests itself in the differentiation and separation of the key physiologic functions which significantly broaden the eco-niche for the species with the consummate type of such separation. However, the organismic level of biodiversity does not work for building any developmental models since the starting point of genetic inheritance and variability processes emerges on the minimum structural unit of the living world only, i.e. the population. It is noted that the sufficient gene pool for species development may accumulate in fairly large populations only, where the general rate of mutation does not yield to the rate of ambient variations. The paper shows that the known formal models of species development based on the Fisher theorem about the impact of genodispersion on species adjustment are not in keeping with the actual existence of the species due to the conventionally finite and steady number of genotypes within a population. On the ecosystem level of life arrangement, the key role pertains to the taxonomic diversity supporting the continuous food chain in the system against any adverse developmental conditions of certain taxons. Also, the progressive evolution of an ecosystem is largely stabilized by its multilayer hierarchic structure and the closed circle of matter and energy. The developmental system models based on the Lotka-Volterra equations describing the interaction of the open-loop ecosystem elements only insufficiently represent the position of biodiversity in the evolutionary processes. The paper lays down the requirements to such models which take into account the mass balance within a system; its trophic structure; the accumulative (buffering) properties of each ecosystem layer; the multiple interconnections among the ecosystem species; and the restricting impact of the intraspecific and interspecific competition. From this perspective, the use of bio-cybernetic concepts and mathematic tools looks quite promising.
Текст научной статьи Биологическое разнообразие и модели эволюции
Значение биологического разнообразия, зафиксированное конвенцией ООН [1], остаётся в настоящее время широко обсуждаемым свойством природы не только в силу его стабилизирующих экосистемных функций и жизнеобеспечивающей роли для человечества, но всё в большей степени как одним из механизмов эволюционных процессов на всех уровнях организации живого мира [2–4]. Актуальность и практическая значимость понимания этого механизма определяется появившейся в настоящее время возможностью целенаправленного влияния на эволюционные процессы природы со стороны «нового геологического фактора» – человека, получившего такое «определение» в работах В. И. Вернадского, раскрывшего антропогенный характер современной биосферы [5]. Научный подход к реализации указанного антропогенного воздействия на природу приводит к необходимости построения соответствующих формальных моделей, отражающих роль многообразия в протекании эволюционных процессов.
-
1.1 Многообразие на уровне организмов
В биологическом плане источником разнообразия живых организмов является соответствующее разнообразие их генетических структур, определяемое, в свою очередь, дискретным набором нуклеотидов, число возможных комбинаций которых в нуклеотидных последовательностях значительно превосходит количество известных в настоящее время белковых соединений.
На уровне отдельных организмов указанное разнообразие достаточно очевидно и проявляется в дифференциации и разделении физиологически важных функций, значительно расширяющих экологическую нишу тех особей, у которых такое разделение наиболее совершенно, рисунок 1.
Защитные жёсткие крылья и мягкие крылья для полёта
Внутривидовое разделение на озимые и яровые формы
Intraspecific division into winter and spring forms
Protective hard wings and soft wings for flight

Water-salt balance sturgeon in the river and the sea
Рисунок 1. Иллюстрация примеров разделения приспособительных функций у различных биологических видов
Figure 1. Illustration of examples of adaptive division of functions in different species
В качестве примеров можно привести существование у жуков Coleoptera мягких и жёстких пар крыльев, дающих жукам возможность полёта и механическую защиту при перемещениях в почве или древесине; наличие у актинобактерий рода Rhodococcus четырёх типов клеток, обеспечивающих, в результате межклеточной кооперации, жизнедеятельность бактериальных колоний в агрессивных средах с высокой степенью загрязнения минеральными солями и нефтепродуктами; существование в жаберном эпителии проходных рыб ионофильтрующих клеток как морского, так и пресноводного типов; образование раннелетних и позднелетних растений, вплоть до разделения на яровые и озимые формы одного вида. Исключительно важным фактом, подтверждающим предпочтительность биологической специализации функций перед их универсальностью, является существование у высших организмов функционально неспециализированных, так называемых стволовых клеток, способных, при возникновении необходимости в регенерации того или иного органа или ткани, дифференцироваться в специализированные клетки.
-
1.2 Многообразие на уровне популяции
Несмотря на отмеченную очевидность приведённых примеров, рассмотренный организменный уровень биологического разнообразия непригоден для построения эволюционной модели в силу того, что минимальной структурной единицей живого мира, в которой возможно протекание эволюционных процессов, является популяция. Более того, накопление генофонда, необходимого для сохранения вида, возможно только в достаточно больших популяциях, в которых общая скорость мутационных изменений не уступает скорости вариаций условий внешней среды. Указанное обстоятельство частично учитывает известная теорема Фишера [ 6 ] , согласно которой средняя скорость приспособленности популяции возрастает пропорционально генной дисперсии (вариации) внутри популяции:
x £ ( w j x j ) x ;
-
V j = 1 7
n
Ё x j = 1; i = 1, n ,
, j = 1
где w i – степень приспособленности особей с генотипом i ; x i – частота встречаемости генотипа i в популяции;
^L w i x i = w , (2)
i = 1
где w – средняя приспособленность всех генотипов; n – число различных генотипов.
Дальнейшее развитие модели Фишера связано с устранением ограничения на исходное разнообразие, накладываемое конечным числом n генотипов, что позволит приблизить эту модель к реальным условиям. К таким условиям следует отнести бесконечно большое разнообразие изменений внешней среды, вследствие которых ограниченный набор генотипов в популяции постепенно будет использован и популяция будет обречена на вымирание. Отметим попутно, что это требование к модели вытекает из известного эволюционного принципа необходимого разнообразия У. Эшби: “Variety can destroy variety”.
-
1.3 Разнообразие на уровне экосистемы
Переходя к биоразнообразию на экосистемном уровне организации жизни, следует, прежде всего, отметить таксономическое многообразие, обеспечивающее в экосистеме непрерывность трофической цепи в условиях, ставших неблагоприятными для развития отдельных таксонов, освободившуюся нишу которых занимают другие виды, ранее не игравшие доминирующего значения, но оказавшиеся более приспособленными к новым условиям.
Кроме таксономического разнообразия важную роль в экосистеме играет её иерархическая строение, образующее структуру с замкнутым циклом (круговоротом) вещества и энергии (рисунок 2). Наиболее известной моделью взаимодействия элементов экосистемы является балансовая модель Лотки-Вольтерры, однако аспекты эволюционной роли многообразия, иерархичности структуры экосистемы и её замкнутость в этой модели не затрагиваются. В этом плане перспективны работы в области биокибернетики [ 7,8 ] . Биокибернетическая модель экосистемы должна учесть следующие свойства экосистемы и особенности её функционирования:
─ баланс массы – сохранение вещества и энергии, приведённой к эквивалентной массе вещества, в их замкнутом круговороте;
─ трофическую структуру передачи вещества в экосистеме, для которой характерна быстро уменьшающаяся доля перерабатываемой массы вещества при переходе от нижних к верхним иерархическим уровням системы – от продуцентов к консументам;
─ накопительные (буферные) свойства каждого уровня экосистемы, способного создавать запас вещества, не полностью включаемого в его общий круговорот;
─ многомерность (множественность) взаимосвязей каждого биологического вида любого уровня экосистемы с каждым видом нижестоящего и вышестоящего смежного уровня;
─ ограничивающее влияние внутривидовой и межвидовой конкуренции каждого уровня экосистемы на скорость потребления и прироста биомассы на рассматриваемом и смежных уровнях трофической цепи.
Energy loss in the form of heat
Потери энергии в виде тепла
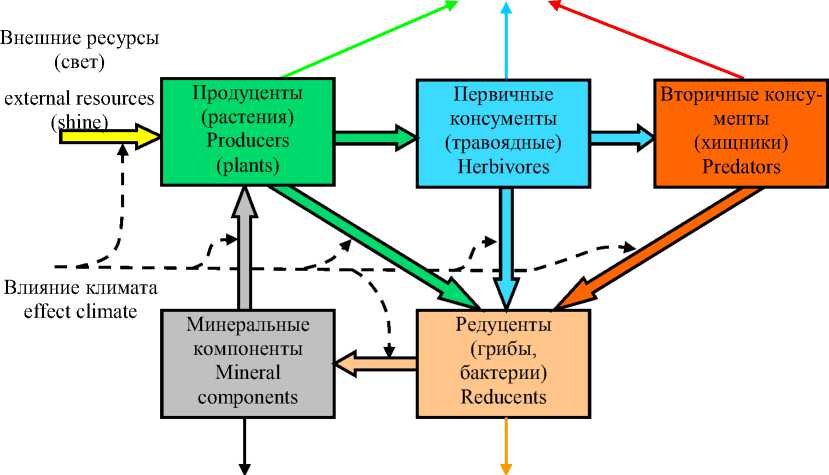
Потери вещества (осадочные породы)
The loss of substance (sedimentary rocks)
Рисунок 2. Замкнутая структура иерархической системы Figure 2. The closed structure of the hierarchical ecosystem
Важным аспектом построения модели функционирования экосистемы является выбор критерия её качества с точки зрения эволюционных возможностей. Традиционное моделирование экосистемы с целью анализа устойчивости не отвечает целям исследования её эволюционных свойств, так как установившееся, стабилизированное, равновесное состояние системы является эволюционно застойным. Эволюционные изменения в экосистеме представляют собою достаточно быстрый в палеонтологическом масштабе времени переходный процесс, который в простейшем случае предполагает переход из одного равновесного состояния в другое, а в более общем плане – заключается в непрерывно существующем неравновесном режиме функционирования экосистемы.
Список литературы Биологическое разнообразие и модели эволюции
- Convention on biological diversity. New York: United Nations Treaty Series, 2001. V. 1760. i. nos. 30690. P. 79-307.
- Nikisianis N., Stamou G.P. Harmony as ideology: questioning the diversity-stability hypothesis//Acta Biotheoretica. 2016. V. 64. Issue 1. P. 33-64.
- Rodríguez R.A. et al. Thermostatistical distribution of a trophic energy proxy with analytical consequences for evolutionary ecology, species coexistence and the maximum entropy formalism//Ecological Modelling. 2014. V. 296. P. 24-35.
- Venjakob C. et al. Plant diversity increases spatio-temporal niche complementarity in plant-pollinator interactions//Ecology and Evolution. 2016. V. 6. Issue 8. P. 2249-2261.
- Вернадский В.И. Биосфера и ноосфера. М.: Айрис-пресс, 2012. 576 с.
- Fisher R.A. The genetical theory of natural selection. Oxford: Oxford University Press, 2011. 318 p.
- Podvalny S.L., Vasiljev E.M. A multi-alternative approach to control in open systems: origins, current state, and future prospects//Automation and Remote Control. 2015. V. 76. № 8. P. 1471-1499.
- Podvalny S.L., Vasiljev E.M., Barabanov V.F. Models of multi-alternative control and decisionmaking in complex system//Automation and Remote Control. 2014. V. 75. № 10. P. 1886-1891.
- Cope E.D. The primary factors of organic evolution. Miami: HardPress Publishing, 2013. 586 p.
- Андреева С.И., Андреев Н.И. Эволюционные преобразования двустворчатых моллюсков Аральского моря в условиях экологического кризиса. Омск: Изд-во Омского гос. педагогич. ун-та, 2003. 382 с.
