Биосинтез бактериальной целлюлозы культурой мedusomyces gisevii

Автор: Гладышева Е.К., Скиба Е.А.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Биотехнология, бионанотехнология и технология сахаристых продуктов
Статья в выпуске: 3 (65), 2015 года.
Бесплатный доступ
Бактериальная целлюлоза - органический материал, синтезируемый внеклеточно микрорганизмами. Бактериальная целлюлоза может быть использована в различных отраслях промышленности. Особенно широкое применение бактериальная целлюлоза нашла в медицине. Получение бактериальной целлюлозы - сложный и длительный процесс. Главным критерием успешного протекания процесса является получение максимального выхода бактериальной целлюлозы. В России нет действующего производства бактериальной целлюлозы, поэтому исследования в данной области являются ультраактуалными. В статье приведены данные по биосинтезу бактериальной целлюлозы культурой Мedusomyces gisevii. Исследовано влияние уровня активной кислотности на синтез бактериальной целлюлозы. Установлено, что для получения бактериальной целлюлозы с помощью симбиоза Мedusomyces gisevii не требуется искусственное поддержания уровня pH. Изучено влияние концентрации субстрата на выход бактериальной целлюлозы. Установлено, что синтез БЦ сопряжен с ростом уксуснокислых бактерий и условия, отвечающие максимуму численности бактерий, соответствуют максимуму выхода БЦ. Максимальная численность наблюдается при концентрации глюкозы в среде 20 г/л, по мере повышения концентрации глюкозы до 55 г/л, численность уксуснокислых бактерий снижается обратно пропорционально концентрации субстрата, что можно объяснить субстратным ингибированием. Концентрация глюкозы 15 г/л и ниже являетсянедостаточной, вследствие чего численность бактерий снижается прямо пропорционально снижению концентрации субстрата. Показано, что максимальный выход БЦ (8,7-9,0 %) обеспечивается при начальной концентрации глюкозы в среде 20-25 г/л. В условиях, отвечающих максимальному выходу бактериальной целлюлозы, был получен укрупненный образец БЦ массой 605 г. Исследованы физико-химические показатели образца бактериальной целлюлозы. Структура и чистота бактериальной целлюлозы подтверждены методом инфракрасной спектрофотометрии.
Бактериальная целлюлоза, симбиоз, биосинтез, активная кислотность, концентрация субстрата, физикохимические показатели, инфракрасная спектроскопия
Короткий адрес: https://sciup.org/14040456
IDR: 14040456 | УДК: 577.114
Biosynthesis of bacterial cellulose by мedusomyces gisevii
Bacterial cellulose is an organic material that is synthesized by microorganisms extracellularly. Bacterial cellulose can be used in various industries. Especially, bacterial cellulose has found its application basically in medicine. The production of bacterial cellulose is a complicated and long process. The principal criterion for the process to be successful is bacterial cellulose to be obtained in a higher yield. Russia is lacking an operating facility to produce bacterial cellulose; therefore, research in this art is the hottest topic. This paper reports details on the biosynthesis of bacterial cellulose by the Мedusomyces gisevii microbe and investigates the effect of active acidity level on the bacterial cellulose synthesis. It was found that the synthesis of bacterial cellulose by the symbiosis of Мedusomyces gisevii does not require pH to be artificially maintained. The substrate concentration effect on the bacterial cellulose yield was also examined. The bacterial cellulose synthesis was witnessed to be conjugated with the acetic-acid bacterium growth, and conditions corresponding to a maximal bacterial cells number correspond to a maximum microbial cellulose yield. The maximal bacterial cell number was observed when the glucose concentration in the broth was 20 g/l; as the glucose concentration was increased to 55 g/L, the acetic-acid bacterial cell number diminished in inverse proportion to the substrate concentration, which is likely due to the substrate inhibition. A glucose concentration of 15 g/l and lower is not enough, causing a decrease in the cell number, which is directly proportional to a decline in the substrate concentration. The maximum bacterial cellulose yield (8.7-9.0 %) was achieved at an initial glucose concentration of 20-25 g/l in the broth. The conditions providing the maximum bacterial cellulose yield gave an enlarged bacterial cellulose specimen 605 g in weight. The physicochemical properties of the bacterial celluloses were studied. The structure and purity of the bacterial celluloses were confirmed by infrared spectrophotometry.
Текст научной статьи Биосинтез бактериальной целлюлозы культурой мedusomyces gisevii
Бактериальная целлюлоза (БЦ) характеризуется химической чистотой, в ней отсутствуют примеси лигнина, гемицеллюлоз и других компонентов, сопутствующих растительной целлюлозе. Молекулярная формула и полимерная структура БЦ, соответствуют целлюлозе, выделенной из растительного сырья, но при этом БЦ обладает более высокой кристалличностью, модулем прочности на разрыв, во-допоглащающей способностью, пластичностью, а также характеризуется биосовместимостью и биоразлагаемостью [1].
БЦ может быть использована в различных отраслях. Широкое применение БЦ нашла в медицине – при изготовлении на основе матриц БЦ, композитных раневых покрытий, включающих наночастицы серебра или селена, обладащих антимикробными, противовоспалительными и заживляющими свойствами. Ещё одним активно развивающимся направлением исследований является создание на основе БЦ систем с контролируемым выделением биологически активных соединений. Также возможно использование БЦ в биотехнологической промышленности, в качестве мембран для иммобилизации ферментов и клеток. В электронной промышленности пленки БЦ используются для изготовления органических светоизлучающих диодов, пленочных солнечных батарей, фотохромных материалов и материалов с жидкокристаллическими свойствами. В целлюлозно-бумажной промышленности БЦ используют для изготовления особых сортов бумаги, салфеток и пеленок, в пищевой промышленности – в качестве пищевых добавок и загустителей. Также БЦ можно использовать для получения эфиров, например нитратов с высокой степенью замещения [2].
К синтезу БЦ способны микроорганизмы разных видов. Их объединяет способность производить внеклеточную целлюлозу в виде гель-пленки на поверхности питательной среды [3].
Получение БЦ – сложный и длительный процесс. Главным критерием успешного протекания процесса является получение максимального выхода БЦ. В России нет действующего производства БЦ, поэтому исследования в данной области являются ультраактуалными.
Основными параметрами, влияющими на синтез и выход целевого продукта, являются концентрация субстрата и уровень активной кислотности [4]. Согласно источнику [5] каждый целлюлозосинтезирующий микроорганизм имеет свой оптимальный диапазон pH, отклонение от которого приводит снижению продуктивности биосинтеза БЦ.
Для продуцентов БЦ характерно субстратное ингибирование [6], преодолеть которое позволяет правильный подбор концентрации субстрата в питательной среде. Концентрация субстрата в питательной среде для различных продуцентов должна устанавливаться экспериментально [4, 5]. Оптимальная концентрация субстрата обеспечивает максимальный выход целевого продукта, а также позволяет повысить его качество за счет сокращения образования побочных продуктов (например, для Acetobacter xy-linum IFO 13693 характерно образование глюконовой кислоты, при концентрации глюкозы в питательной среде более 24 г/л [3]).
В данной работе в качестве продуцента использована культура Мedusomyces gisevii , известная как «чайный гриб». Напиток, образующийся в результате культивирования «чайного гриба», широко используется в лечебно-профилактических целях, поскольку положительно влияет на организм человека, благодаря микроорганизмам симбиоза, обладающих антимикробными свойствами и продуцирующих комплекс биологически активных веществ. Технология получения напитков на основе «чайного гриба» в настоящее время продолжает развиваться и занимает значимое место в группе лечебно-профилактических безалкогольных напитков [7].
Продуцент Мedusomyces gisevii представляет собой симбиотическую культуру, состоящую из разных видов уксуснокислых бактерий и дрожжей [4]. Симбиоз обладает огромным адаптивным потенциалом. Благодаря подвижным симбиотическим отношениям культура легко приспосабливается к изменению внешних условий, стрессам, способна утилизировать различные субстраты, устойчива к фагам [4].
Целью данной работы является биосинтез продуцентом Мedusomyces gisevii гель-пленки БЦ . Для достижения данной цели были решены следующие задачи: исследовано влияние уровня активной кислотности на синтез БЦ, изучено влияние концентрации субстрата на выход бактериальной целлюлозы; в условиях, отвечающих максимальному выходу БЦ, получен укрупнённый образец БЦ, исследованы его физико-химические показатели.
Для экспериментов использованы синтетические питательные среды, приготовленные растворением глюкозы в экстракте черного чая. Доза внесения инокулята составила 10 %. Культивирование проводилось в статических условиях при
30 ±0,4°С. Убыль редуцирующих сахаров контролировалась спектрофотометрически (спектрофотометр «UNICOUV-2804», США) с использованием динитросалицилового реактива, прирост пленки БЦ оценивался гравиметрически (взвешивались суховоздушные образцы весы лабораторные аналитические Explorer EX-224), уровень активной кислотности определялся мощью иономера (иономер И-160 МИ). Микробиологические показатели определяли с методом прямого подсчёта клеток на камере Горяева.
Для исследования влияния уровня активной кислотности на синтез БЦ использовалась питательная среда с начальной концентрацией глюкозы 25 г/л. Изучено пять вариантов, различающихся уровнем рН: 3,0; 4,0; 5,0 и 6,0. Активная кислотность на заданном уровне поддерживалась вручную ежесуточно путем внесения молочной кислоты или аммиака. В качестве контроля использовалась среда с нативным рН (4,1 ед. рН), в контрольном опыте значения активной кислотности фиксировались в ходе эксперимента, но не корректировались.
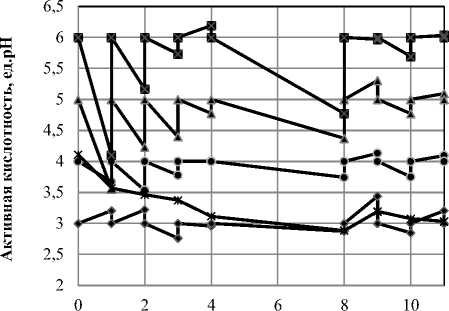
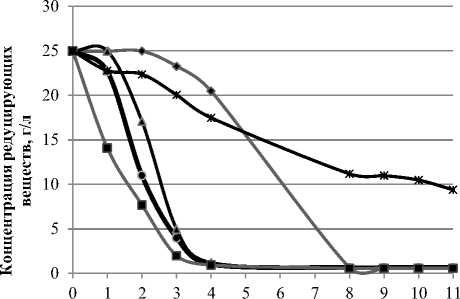
На рисунке 1 представлено изменение уровня активной кислотности сред при культивировании Мedusomyces gisevii. На рисунке 2 представлено изменение концентрации редуцирующих веществ в зависимости от уровня активной кислотности.
Продолжительность культивирования, сутки ф 3 I—е^— 4 —*—5 —о— 6 ж контроль
Рисунок 1. Изменение активной кислотности сред при культивировании Мedusomyces gisevii на средах с разным уровнем рН
Продолжительность культивирования, сутки
—ф— 3 ^■•^* 4 —А— 5 6 контроль
Рисунок 2. Изменение концентрации редуцирующих веществ при культивировании Мedusomyces gisevii на средах с разным уровнем рН
На рисунке 1 видно, что во всех вариантах активная кислотность в процессе культивирования снижалась, особенно значительно уровень pH понизился при pH 5,0 и 6,0 на первые сутки культивирования; исключение представляет образец с начальным pH 3,0 для которого происходит повышение значения рН, то есть микроорганизмы саморегулировали уровень pH сред и приближали его значение к оптимальному. Такое же поведение наблюдалось на 3 и 4 сутки. На четвертые сутки изменение pH не было сильно заметно, далее в течении трех суток уровень pH не контролировался. К 8 суткам произошло снижение или повышение pH, т.е. микроорганизмы снова пытались стабилизировать уровень активной кислотности. На 9 и 10 сутки кислотность не сильно отличалась от заданного уровня. На 11 сутки значение активной кислотности перестало изменяться, что может свидетельствовать о гибели микроорганизмов.
По данным рисунка 2 видно, что потребление субстрата в опытах, где уровень активной кислотности искусственно поддерживался на заданном уровне, протекало быстрее, чем в контроле. Однако образование гель-плёнки БЦ произошло только в контрольном варианте.
Можно предположить, что при отклонении рН от нативного субстрат тратится не на синтез БЦ, а на поддержание жизнеспособности микроорганизмов. При этом сдвигается равновесие между симбионтами и создаются условия, неблагоприятные для развития целлюлозосинтезирующих микроорганизмов. При рН 5 и 6 отмечено газообразование, гель-плёнка отсутствует; при рН 4 образовались рваные нити БЦ, покрытые слизью; при рН 3 синтез БЦ не происходил.
Из полученных данных можно сделать вывод, что для получения БЦ с помощью симбиоза Мedusomyces gisevii не требуется искусственное поддержания уровня pH.
Для изучения влияния концентрации субстрата на выход бактериальной целлюлозы использовались питательные среды, с разными начальными концентрациями глюкозы: 5, 10, 15, 20, 25, 35, 45, 55 г/л. Культивирование проводилось в статических условиях при 27 °С.
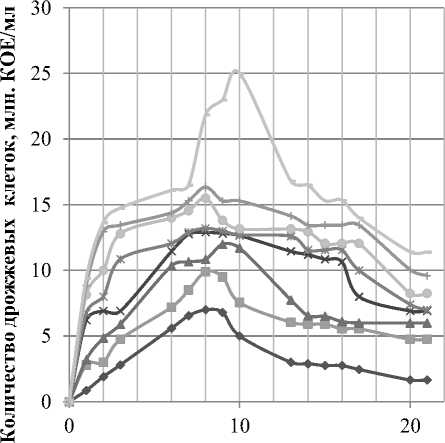
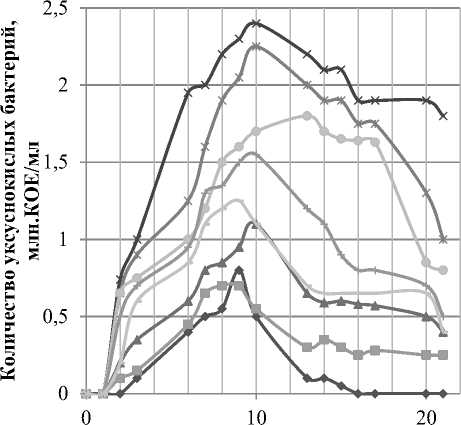
На рисунке 3 и 4 показаны изменения количества клеток дрожжей и уксуснокислых бактерий в зависимости от различной концентрации глюкозы в питательной среде.
Продолжительность культивирования, сутки
Продолжительность культивирования, сутки
5 10 1520
25 35 4555
5 10 1520
25 35 4555
Рисунок 3. Изменение количества дрожжевых клеток при культивировании Medusomyces gisevii на средах с разной концентрацией глюкозы
Рисунок 4. Изменение количества уксуснокислых бактерий при культивировании Medusomyces gisevii на средах с разной концентрацией глюкозы
Из рисунка 3 можно сделать вывод, что при увеличении концентрации глюкозы в питательной среде прямо пропорционально увеличивается концентрация дрожжевых клеток. Также следует отметить, что численность дрожжевых клеток изменялась прямо пропорционально концентрации субстрата в среде: при концентрации глюкозы в питательной среде 5, 10, 15, 20, 25, 35, 45 г/л увеличение количества дрожжевых клеток происходило до 8 суток культивирования, далее отмечалось их снижение; при концентрации 55 г/л увеличение численности дрожжевых клеток происходило до 10 суток, а потом также происходил спад.
Численность уксуснокислых бактерий зависит от концентрации глюкозы в среде не столь однозначно. Из рисунка 4 следует, что по максимальной численности уксуснокислых бактерий в зависимости от концентрации глюкозы в среде результаты экспериментов можно представить в порядке убывания: 20>25>35>45>55>15>10>5. Максимальная численность наблюдается при концентрации глюкозы в среде 20 г/л, затем, по мере повышения концентрации глюкозы до 55 г/л, численность уксуснокислых бактерий снижается обратно пропорционально концентрации субстрата, что можно объяснить субстратным ингибированием. Концентрация глюкозы 15 г/л и ниже является недостаточной, вследствие чего численность бактерий снижается прямо пропорционально снижению концентрации субстрата.
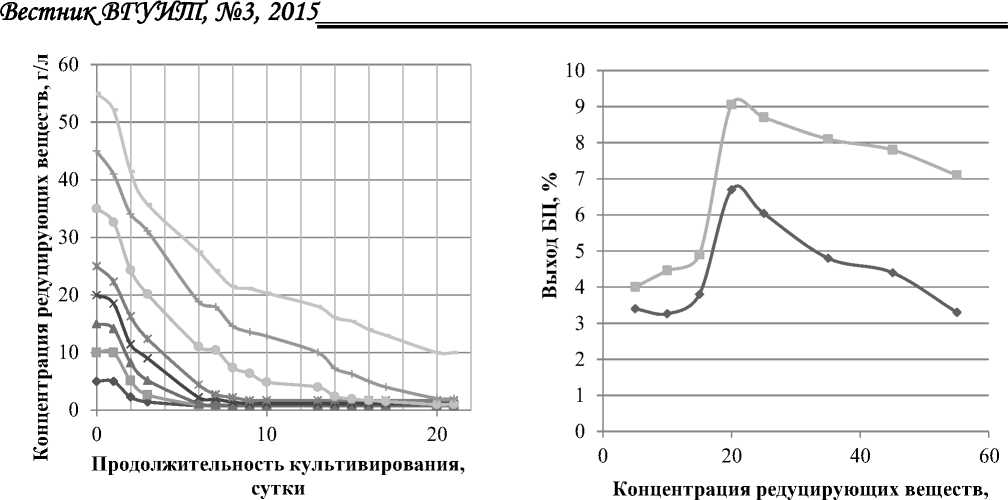
На рисунке 5 представлены изменения концентрации редуцирующих веществ в процессе культивирования Medusomyces gisevii на средах с различной концентрацией глюкозы. На рисунке 6 представлена зависимость выхода БЦ от различной концентрации глюкозы в питательной среде на 7 и 21 сутки культивирования.
—0= 5 г/л ■ 10 г/л —*— 15 г/л 20 г/л г/л
25 г/л • 35 г/л I 45 г/л ^^^^^^^* 55 г/л
—0= 7 сутки ■ 21 сутки
Рисунок 5. Изменения концентрации редуцирующих веществ в процессе культивирования Medusomyces gisevii на средах с различной концентрацией глюкозы
Рисунок 6. Зависимость выхода БЦ от различной концентрации субстрата в питательной среде
Из рисунка 5 видно, что потребление субстрата при разной концентрации глюкозы в среде протекает неодинаково. Полная утилизация субстрата при концентрации глюкозы 5, 10 и 15 г/л зафиксирована на 6 сутки культивирования, при концентрации 20 и 25 г/л – на 10 сутки, при концентрации 35 г/л – на 17 сутки, при концентрации 45 г/г – на 20 сутки. При концентрации 55 г/л не происходит полной утилизации субстрата, в питательной среде остается около 10 г/л глюкозы.
На рисунке 6 представлена зависимость выхода БЦ от различной концентрации глюкозы в питательной среде на 7 и 21 сутки культивирования. Наиболее высокий выход БЦ обеспечивают концентрации глюкозы в питательной среде 20 и 25 г/л. Выход составил на 7 сутки 6,8 и 6,1 %, на 21 сутки – 9,0 и 8,7 %. Следует отметить, что более длительное культивирование вести нецелесообразно, т.к. основной рост БЦ происходит в первые 7 – 10 суток культивирования, далее за последующие 11-21 сутки выход увеличивается всего на 3,2-2,6 %. Также ухудшается состояние пленок: теряется эластичность, пленки становятся неравномерными, водянистыми, начинают расслаиваться. Вследствие этого увеличивается продолжительность их очистки.
По выходу БЦ начальные концентрации глюкозы (г/л) в питательных средах можно по мере убывания расположить в ряд: 20>25>35>45>55>15>10>5. Таким образом, синтез БЦ сопряжен с ростом уксуснокислых бактерий и условия, отвечающие максимуму численности бактерий, соответствуют максимуму выхода БЦ. Следовательно, БЦ является первичным метаболитом целлюлозосинтезирующих микроорганизмов.
С учетом изученных факторов был проведен синтез БЦ в реакторе объёмом 16 л, коэффициент заполнения 50 %. Использовалась синтетическая питательная среда, с начальной концентрацией глюкозы 20 г/л, уровень активной кислотности саморегулировалcя симбиозом. Культивирование проводилось в статических условиях при 26±2 °С в течение 12 суток.
На рисунке 7 представлено фото укрупненного образца бактериальной целлюлозы. Диметр гель-плёнки 33 см, толщина 0,9±0,1 см, масса влажного образца составила 605 г, масса БЦ в пересчете на абсолютно сухое вещество – 6,9 г. Таким образом, выход целевого продукта составил 4,25 %. Можно констатировать, что при увеличении объёма культуральной жидкости от 100 мл до 8 л, отмечается снижение выхода БЦ в 2 раза. Возможно, это связано с колебаниями температуры в процессе культивирования.

Полученный образец БЦ был проанализирован по физико-химическим показателям. Структура полученного образца была исследована с помощью инфракрасного спектрофотометра «Инфралюм ФТ-801» в таблетках KBr.
Определение физико-химических характеристик БЦ (массовой доли кислотонерастворимого лигнина, массовой доли золы, массовой доли альфа-целлюлозы, степени полимеризации целлюлозы) проводили согласно стандартным методикам [7]. Влажность была найдена на определителе влажности МВ 23.
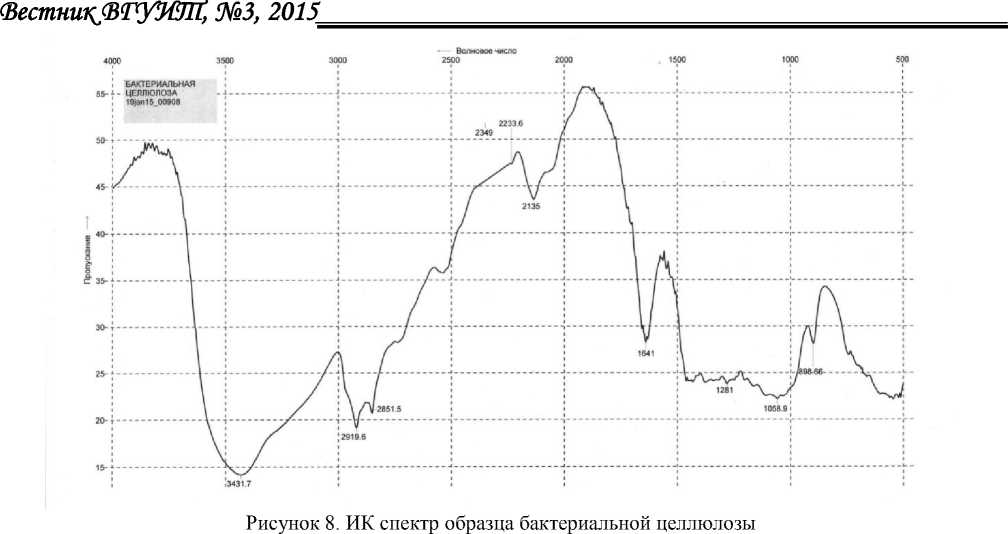
На рисунке 8 представлен ИК-спектр образца бактериальной целлюлозы. Полосы поглощения функциональных групп в образце бактериальной целлюлозы соотносятся с основными функциональными группами БЦ, представленные в литературе [8]. Пик при 3432 см-1 обусловлен валентными колебаниями OH-групп. Пик при 2919 и 2852 см-1 характеризует валентные колебания групп CH и CH2. Пик при 1641 см-1 обусловлен колебаниями связей прочно связанной воды. Полоса при 1281 указывает на валентные колебания OH-групп в спиртах. Полоса поглощения при 1059 см-1 обусловлена в основном валентными колебаниями C-O-C и C-O в спиртах. Полоса при 899 см-1 подтверждает наличие β-1,4 связей. ИК-спектры показывают, что БЦ не содержит примесей, в частности лигнина, присутствующего в растительной целлюлозе, так как отсутствуют полосы поглощения, характерные для ароматических примесей (лигнина), что доказывает чистоту БЦ.
Воздушно-сухая БЦ содержит 4,7 масс. % влаги. Степень полимеризации БЦ составила 2000, что соответствует степени полимеризации растительной хлопковой целлюлозы [9], пригодной для химической модификации.
Известно, что в составе БЦ отсутствуют примеси лигнина и других сопутствующих компонентов [1]. Для подтверждения чистоты БЦ в образце была определена массовая доля лигнина, нерастворимого в серной кислоте, которое составило 0,80 %. Его содержание оказалось выше, чем в хлопковой целлюлозе – 0,50 % [9]. Предположительно, это не истинный лигнин, а лигниноподобные вещества, нерастворимые в серной кислоте, например, фенольные и красящие вещества чая.
Массовая доля зольности в образце БЦ составила 0,14 %. Полученный результат сопоставим с зольностью хлопковой целлюлозы – 0,10 % [9]. При попытке определить массовую долю альфа-целлюлозы по стандартной методике для хлопковой целлюлозы было обнаружено, что измельченный образец, помещенный в 17,5 % раствор гидроксида натрия, не набухает и не растворяется. Содержание альфа-цел-люлозы в образце составило 101,6 % – это свидетельствует о недостаточной промывке образца от гидроксида натрия, а также о высоком содержании высокополимерной целлюлозы.
По результатам работы можно сделать следующие выводы:
-
- исследовано влияние уровня активной кислотности на синтез БЦ. Установлено, что для получения БЦ с помощью симбиоза
Список литературы Биосинтез бактериальной целлюлозы культурой мedusomyces gisevii
- Belgacem M.N., Gandini A. Monomers, Polymers and Composites from Renewable Resources. Amsterdam: Elsevier. 2008, 553 p.
- Гладышева Е.К. Обоснование выбора питательной среды для синтеза бактериальной целлюлозы//Вестник Алтайской науки. 2014. № 1. С. 307-310.
- Koon-Yang Lee, Gizem Buldum, Anthanasios Mantalaris at al. More than Meets the Eye in Bacterial Cellulose: Boisynthesis, Bioprocessing, and Applications in Advanced Fiber Composites//Macromolecular Bioscience. 2014. № 6. P. 10-32.
- Юркевич Д.И., Кутышенко В.П. Медузомицет (Чайный гриб): научная история, состав, особенности физиологии и метаболизма//Биофизика. 2002. № 6. С. 1116-1129.
- Goh W.N., Rosma A., Kaur B. et al. Fermentation of black tea broth (Kombucha): I. Effects of sucrose concentration and fermentation time on the yield of microbial cellulose//International Food Research Journal. 2012. № 19(1). P. 109-117.
- Masaoka S., Ohe T., Sakota N. Production of cellulose from glucose by Acetobacter xylinum//Journal of Fermentation and Bioengineering. 1993. № 75 (1). P. 18-22.
- Жумабекова Б.К., Жумабекова К.А. Технология получения чайного кваса с добавлением экстракта душицы//Фундаментальные исследования. 2015. № 2 (11). С. 2370-2373.
- Оболенская А.В., Ельницкая З.П., Леонович А.А. Лабораторные работы по химии древесины и целлюлозы: учебное пособие для вузов. М.: Экология, 1991. 320 с.
- Xueqiong Yin, Changjiang Yu, Xiaoli Zhang at al. Comparison of succinylation methods for bacterial cellulose and adsorption capacities of bacterial cellulose derivatives for Cu2+ ion//Polymer Bulletin. 2011. № 67. P. 401-412.
- Гисматулина Ю.А., Будаева В.В. Сравнение целлюлоз, выделенных из мискантуса, с хлопковой целлюлозой методом ИК-Фурье спектроскопии//Ползуновский вестник. 2014. № 3. С. 177-181.
