Биосинтез фотопреобразующего трансмембранного белка [2Н] бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот [2,3,4,5,6- 2H 5] Phe, [3,5- 2H 2] Tyr и [2,4,5,6,7- 2H 5] Trp
![Биосинтез фотопреобразующего трансмембранного белка [2Н] бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот [2,3,4,5,6- 2H 5] Phe, [3,5- 2H 2] Tyr и [2,4,5,6,7- 2H 5] Trp](/file/picture/14264854/biosintez-fotopreobrazujushhego-transmembrannogo-belka-2n-bakteriorodopsina.png "Биосинтез фотопреобразующего трансмембранного белка [2Н] бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот [2,3,4,5,6- 2H 5] Phe, [3,5- 2H 2] Tyr и [2,4,5,6,7- 2H 5] Trp")
Автор: Мосин Олег Викторович, Швец В.И., Складнев Д.А., Игнатов Игнат
Журнал: Научное приборостроение @nauchnoe-priborostroenie
Рубрика: Методы измерений
Статья в выпуске: 2 т.23, 2013 года.
Бесплатный доступ
Осуществлен биосинтез фотопреобразующего трансмембранного белка бактериородопсина (выход 8-10 мг), меченного дейтерием по остаткам функционально-важных аминокислот — [2,3,4,5,6- 2H 5]Phe, [3,5- 2H 2]Tyr и [2,4,5,6,7- 2H 5]Trp, что важно для наноиндустрии новых современных отечественных фотопреобразующих наноматериалов и биомолекулярной электроники. Белок выделен из мембран экстремальных фотоорганогетеротрофных галобактерий H alobacterium halobium лизисом клеток в дистиллированной воде, обработкой бактериальной биомассы ультразвуком, спиртовой экстракцией низко- и высокомолекулярных примесей, клеточной РНК, каротиноидов и липидов с последующей солюбилизацией конечного продукта в 0.5 %-м ДДС-Na и фракционированием метанолом. Гомогенность синтезируемого [ 2Н]бактериородопсина и селективность включения дейтерия в молекулу исследованы комбинацией методов разделения и анализа белка, включая электрофорез в 12.5 %-м ПААГ с 0.1 %-м ДДС-Na, гель-проникающую хроматографию на сефадексе G-200, ОФ ВЭЖХ, масс-спектрометрию ЭУ метиловых эфиров N-(диметиламино)нафтален-1-сульфонил хлоридных (дансильных) производных аминокислот.
[2н]бактериородопсин, биосинтез, масс-спектрометрия эу
Короткий адрес: https://sciup.org/14264854
IDR: 14264854 | УДК: 579.871.08+577.112.385.4.08
Biosynthesis of transmembrane photo-transforming protein [2н] bacteriorhodopsin, labelled with deuterium on residues of aromatic amino acids [2,3,4,5,6- 2H 5] Phe, [3,5- 2H 2] Tyr и [2,4,5,6,7- 2H 5] Trp
The microbial micropreparative synthesis of transmembrane photo-transforming protein bacteriorhodopsin (output 8-10 mg), labeled with deuterium on residues of [2,3,4,5,6- 2H 5]phenylalanine, [3,5- 2H 2]tyrosine, and [2,4,5,6,7- 2H 5]tryptophan was carried out that is important for nanoindustry of new modern domestic photo-transforming nanomaterials and molecular bioelectronics. The protein was allocated from membranes of photoorganoheterotrophic halobacterium Halobacterium halobium by cellular lysis by distilled water, processing of bacterial biomass by ultrasound, alcohol extraction of low and high-weight molecular impurities, cellular RNA, carotenoids and lipids, with the subsequent solubilization of final product with 0.5 % SDS-Na and fractionation by methanol. The homogenity of the synthesized product, and the selectivity of deuterium encorporation into the molecule was proved by combination of preparative and analytical protein methods including elecrtophoresis in 12.5 % PAAG with 0.1 % SDS-Na, gel filtration chromatography on Sephadex G-200, reverse-phase HCLP, electron impact mass-spectrometry of methyl esters of N-Dns-derivatives of amino acids.
Текст научной статьи Биосинтез фотопреобразующего трансмембранного белка [2Н] бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот [2,3,4,5,6- 2H 5] Phe, [3,5- 2H 2] Tyr и [2,4,5,6,7- 2H 5] Trp
Бактериородопсин (БР), названный по аналогии с белком зрительного аппарата млекопитающих родопсином, был выделен в 1971 г. Д. Остерхель-том и В. Стохениусом (США) из клеточной мембраны экстремальной фотоорганогетеротрофной галобактерии Halobacterium halobium [1]. Этот фотопреобразующий белок молекулярной массой 26 кДа представляет собой хромопротеид, связанный шиффовым основанием (как в зрительных пигментах животных) с остатком Lys-216, который содержит в качестве хромоформной группы эквимолекулярную смесь 13- цис - и полностью 13- транс -ретинольного С 20 -каротиноида, определяющего пурпурно-красный цвет галофильных бактерий. Наряду с БР в клеточной мембране га-лобактерий содержится небольшое количество других сопутствующих каротиноидных пигментов, основной из которых — бактериоруберин, определяющий устойчивость галобактерий к солнечному излучению [2].
Полипептидная цепь БР состоит из 248 аминокислотных остатков, 67 % которых являются гидрофобными, образованными с участием ароматических аминокислот — фенилаланина, тирозина и триптофана [3]. Согласно структурным исследова- ниям, молекула БР состоит из семи находящихся в конформации α-спирали сегментов, пронизывающих всю толщу мембраны клетки в направлении, перпендикулярном ее плоскости [4]. Гидрофобные домены представляют собой трансмембранные сегменты, а гидрофильные домены выступают из мембраны и соединяют отдельные внутримем-бранные α-спиральные тяжи белковой молекулы [5].
Благодаря своей структуре молекула БР выполняет функции светозависимого протонного насоса, перекачивающего протоны через мембрану клетки и создающего электрохимический градиент протонов Н+ на поверхности клеточной мембраны, энергия которого используется клеткой для синтеза АТФ в анаэробном фотосинтетическом фосфорилировании. При поглощении кванта света происходит обратимая изомеризация 13- Z -БР ( λ макс = 548 нм) (квантовый выход 0.03 при 20 ºС) в all-E -БР ( λ макс = 568 нм) [6], инициирующая каскад фотохимических реакций с образованием промежуточных интермедиантов J 625 , K 590 , L 550 , М 412 , N 560 и O 640 с последующим отрывом Н+ из ретинального остатка БР и его присоединением со стороны цитоплазмы (рис. 1). В этом процессе молекула ретиналя изгибается в мембранном туннеле и переносит протон Н+ с внутренней цитоплаз-
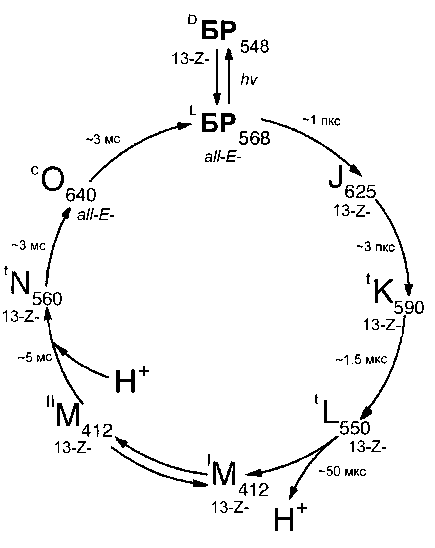
Рис. 1. Фотохимические реакции превращения БР (водная суспензия, рН ∼ 7.2, 20 ºС) [6].
Латинские цифры обозначают спектральные интермедианты БР. Верхние индексы "c" и "t" относятся к циклам 13- цис - и 13- транс -производных БР, "L" и "D" обозначают световую и темновую формы пигментов. Нижние индексы соответствуют положению максимума поглощения интермедиаторов фотоцикла (в нм)
матической мембраны на внешнюю мембрану клетки. В результате между внутренней и внешней поверхностями мембраны образуется градиент концентрации Н+, приводящий к тому, что освещенные клетки галобактерий начинают синтезировать АТФ, т. е. преобразуют энергию света в химическую энергию связей. Этот процесс обратим и в темноте протекает в обратном направлении, что позволяет галобактериям развиваться в темноте, переключая фотосинтетический метаболизм на гетеротрофный [7].
Несмотря на структурно-функциональную изученность, БР остается в центре внимания био- и нанотехнологии прежде всего благодаря своей высокой светочувствительности и разрешающей способности и используется в молекулярной биоэлектронике, как природный фотохромный материал, в качестве управляемых светом или электрическими импульсами модулей компьютерных и оптических систем [8]. БР-содержащие нанопленки, полученные с использованием БР-содержащих пурпурных мембран галобактерий впервые были получены и исследованы в нашей стране в рамках проекта "Родопсин", в ходе которого продемонстрирована эффективность и перспективы использования наноматериалов "Биохром" в качестве фотохромных материалов для голографической записи. Основной задачей при изготовлении БР-содержащих нанопленок является ориентация пурпурных мембран между гидрофобными и гидрофильными средами, например между водой и воздухом, как это распространено в природе. Как правило, для улучшения характеристик БР-содержащих пленок, используется несколько слоев, которые наносятся на поверхность полимерного носителя и высушиваются в определенных условиях с сохранением своей природной структуры. Наилучшие показатели достигаются при изготовлении пленок на основе желатина. Это позволяет добиться высокой концентрации БР (до 50 %) в нанопленках, избежать агрегации фрагментов пурпурных мембран, а также разрушения бактериородопсина в процессе изготовления. Встроенные в желатиновую матрицу фрагменты пурпурных мембран долговечны (около 104 ч) и устойчивы к воздействию многих факторов как при изготовлении, так и в процессе эксплуатации (колебания температуры, интенсивное воздействие светом с помощью лазера и др.).
Кроме этого, БР очень привлекателен как модельный объект для исследований, связанных с изучением функциональной активности и структурных свойств фотопреобразующих мембранных белков в составе искусственных нативных энерго-и фотопреобразующих мембран [9].
Для структурно-функциональных исследований с БР целесообразно вводить в молекулу белка в качестве изотопной метки дейтерий (2H), позволяющий использовать для анализа изотопного включения метод высокочувствительной масс-спектрометрии ЭУ [10]. Большое научно-прикладное значение имеет БР, меченный дейтерием по остаткам функционально-важных ароматических аминокислот — [2H]фенилаланина, [2H]тирозина и [2H]триптофана, участвующих в гидрофобном взаимодействии полипептидной цепи белка с липидным бислоем клеточной мембраны [11]. 2Н-меченые ароматические аминокислоты могут быть синтезированы методом обратного изотопного обмена (1Н-2Н) в молекулах протонированных аминокислот — [2,3,4,5,6-2Н 5 ]фенилаланин в 85 %-й 2H 2 SO 4 при 50 ºC, [3.5-2H 2 ]тирозин в 15 %-й 2H 2 SO 4 при слабом кипячении реакционной смеси, [2,4,5,6,7-2H 5 ]триптофан в 75 %-й [2H]трифторук-сусной кислоте при 25 ºС [12] . Однако, несмотря на изученность современных методов химического синтеза [2Н]ароматических аминокислот, отечественная индустрия индивидуальных [2Н]трансмем-бранных фотопреобразующих белков не получила должного развития.
Целью настоящей работы являлось разработка технологии микропрепаративного получения БР, меченного дейтерием по остаткам функциональноважных ароматических аминокислот [2,3,4,5,6-2Н 5 ]фенилаланина, [3,5-2H 2 ]тирозина и [2,4,5,6,7-2H 5 ]триптофана, для реконструкции искусственных мембран, а также исследование уровней дей-терированности молекулы БР методом масс-спектрометрии ЭУ метиловых эфиров N-(диметиламино)нафтален-1-сульфонил хлоридных (дансильных) производных аминокислот c последующим разделением методом ОФ ВЭЖХ.
ПРИБОРЫ И МАТЕРИАЛЫ
Для синтеза [2Н]аминокислот использовали протонированные D , L- аминокислоты ("Reanal", Венгрия), 2Н 2 О (99.9 ат. % 2H) и 2HСl (95.5 ат. % 2H), полученные из Российского научно-исследовательского центра "Изотоп" (Санкт-Петербург, РФ). Для синтеза производных аминокислот использовали 5-(диметиламино)нафтален-1-сульфо-нил хлорид (Dns-хлорид) ("Sigma", США) и диазометан, получаемый из N-нитрозомочевины ("Merck", ФРГ). Неорганические соли предварительно перекристаллизовывали в 2Н 2 О, 2H 2 O дистиллировали над KMnO 4 с последующим контролем изотопной чистоты 1Н-ЯМР-спектроскопией на приборе Brucker WM-250 ("Brucker Daltonics", ФРГ) (рабочая частота 70 МГц, внутренний стандарт Me 4 Si).
Электрофорез БР проводили в 12.5 %-м ПААГ с 0.1 %-м ДДС-Na в соответствии с протоколом фирмы LKB (Швеция). Для количественного определения содержания синтезированного в клетке белка проводили сканирование прокрашенного в растворе "кумасси-голубой" R-250 электрофоретического геля на лазерном денситометре Beckman CDS-200 ("Beckman Coulter", США).
Центрифугирование проб проводили на центрифуге Т-26 ("Carl Zeiss", ФРГ) при 2000 g 10 мин.
Масс-спектры метиловых эфиров N-Dns-производных аминокислот получали методом электронного удара на приборе МB-80A ("Hitachi", Япония) при энергии ионизирующих электронов 70 эВ, ускоряющем напряжении 8 кВ и температуре катодного источника 180 - 200 ºС. Сканирование анализируемых образцов проводили при разрешении 7500 усл. ед., используя 10 % резкость изображения.
Спектры 1Н-ЯМР регистрировали в 2Н2О на приборе Brucker WM-250 ("Brucker Daltonics", ФРГ) с рабочей частотой 70 МГц; химические сдвиги протонов ( δ , ppm) измерены в миллионных долях по отношению к Ме 4 Si.
Спектры поглощения регистрировали на программируемом спектрофотометре Beckman DU-6
("Beckman Coulter", США) в диапазоне длин волн 200 - 700 нм.
ОФ ВЭЖХ проводили на жидкостном хроматографе Knauer Smartline ("Knauer", ФРГ), снабженном насосом Gilson ("Gilson Inc.", ФРГ), детектором UF-2563 и интегратором СR-3A ("Shimadzy", Япония), используя колонку 250 × 10 мм с неподвижной обращенной фазой сепарон С18 (Kova, Словакия); элюент: (А) — ацетонитрил— трифторуксусная кислота 100: 0.1–0.5 об. % и (Б) — ацетонитрил 100 об. %; скорость элюирования — 1.5 мл / мин: от 0 до 20 % Б — 5 мин, от 20 до 100 % Б — 30 мин, 100 % Б — 5 мин, от 100 до 0 % Б — 2 мин, 0 % Б — 10 мин. ТСХ N-Dns-производных аминокислот проводили на хроматографических пластинках с закрепленным слоем флуоресцентного носителя Silufol UV-254 ("Kava-lier", Словакия) в системе растворителей (A): н-бутанол—уксусная кислота—вода (12 : 3 : 5 об. %).
УСЛОВИЯ ЭКСПЕРИМЕНТА
Штамм-продуцент БР
В качестве продуцента БР использовали каро-тиноидсодержащий штамм экстремальных фото-органогетеротрофных галобактерий Halobacterium halobium ЕТ 1001, полученный из коллекции культур МГУ. Штамм модифицирован селекцией отдельных колоний на твердой (2 %-й агар) пептоновой среде с 4.3 М NaCl.
Синтез L -[2,3,4,5,6- 2 H 5 ]фенилаланина
40 г фенилаланина растворяли в 300 мл 85 % 2Н 2 SО 4 (в 2Н 2 О) и нагревали с обратным водяным холодильником при 50 - 60 ºС при перемешивании 3 сут. По окончании реакционную смесь охлаждали, нейтрализовали 30 %-м NH 4 OН до рН 5.5. Продукт экстрагировали этанолом. Выход 24 г (58.7 %); Т пл = 271 - 273; [ α ] d 25 = 4.47 (5 М НСl); рK a = 2.20 (СООН), 9.31 (NH 2 ); R f = 0.6 (A). УФ-спектр (0.1 М НCl): λ макс [нм] ( ε М–1 см–1) = 257.5 ( ε = 195). Спектр 1Н-ЯМР (2Н 2 SO 4 в 2Н 2 О) ( δ , ppm): δ 3.25 (2H, H β ), δ 4.4 (1H, H α ), δ 7.2 - 7.4 (0.07Н), УД = 90 ат.% 2Н. Масс-спектр ЭУ ([M]+ — m/z ( I , %): 165 (34)), метиловый эфир N-Dns-[2,3,4,5,6-2H 5 ]фенилаланина: 417 (14), 418 (6).
Синтез L -[3,5- 2 H 2 ]тирозина
100 г тирозина растворяли в 150 мл 3 М 2Н 2 SO 4 . Реакционную смесь нагревали 2 сут. при 40 - 50 ºС с обратным водяным холодильником в токе азота. По окончании нейтрализовали 30 %-м NH4OH до рН 4.5 и охлаждали 1 сут. при 4 ºС. Кристаллический продукт фильтровали, промывали 2Н 2 О и сушили при 10 мм рт. ст. Выход 90 г (86.5 %);
Т пл = 316–317; [ α ] d 25 = 10.03 (5 М НСl); рK a = 2.20 (СООН), 9.21 (NH 2 ); R f = 0.45 (A). УФ-спектр (0.1 М НCl): λ макс [нм] ( ε М–1 см–1) = 223 ( ε = 8200) и 274.5 ( ε = 1340). Спектр 1Н-ЯМР (1 М 2НCl) ( δ , ppm): δ 3.32 (2H), δ 4.35 (1H), δ 6.9 (1H), δ 7.2 (2H), УД = = 96 ат.% 2Н. Масс-спектр ЭУ ([M]+ — m/z ( I , %)): 181 (21); метиловый эфир N-Dns-[3,5-2H 2 ] тирозина: 429 (15), 430 (5).
Синтез L-[2,4,5,6,7-2H5]триптофана
К 40 мл 100 % 2Н 2 О добавляли при 4 ºС и перемешивании 80 мл трифторуксусного ангидрида (0.5 моль) и выдерживали 2 ч при 4 ºС, затем дробными порциями добавляли 25 г триптофана. Реакционную смесь выдерживали 3 сут. в темноте при 22 ºС, растворитель удаляли при 10 мм рт. ст., нейтрализовали 30 %-м NH 4 OH до рН 5.9, охлаждали 1 сут. при 4 ºС. Кристаллический продукт фильтровали, промывали 2Н 2 О и сушили при 10 мм рт. ст. Выход 19 г (60.3 %); Т пл = 283 - 285; [ α ] d 25 = 2.8 (1 М НСl); рK a = 2.46 (СООН), 9.41 (NH 2 ); R f = 0.5 (A). УФ-спектр (0.1 М НCl): λ макс [нм] ( ε М–1 см–1) = 218 ( ε = 3350), 278 ( ε = 5550), 287.5 ( ε = 4550). Спектр 1Н-ЯМР (CF 3 COOH в 2Н 2 О) ( δ , ppm): δ 3.4 (2H, H β ), δ 4.4 (1H, H α ), δ 7.3 (1H, H ε ), δ 7.2 - 7.4 (0.1Н, In-Н), УД = 98 ат.% 2Н. Масс-спектр ЭУ ([M]+ — m/z ( I , %)): 204 (28); метиловый эфир N-Dns-[2,4,5,6,7-2H 5 ]триптофана: 455 (9), 456 (11).
Синтез N-Dns-аминокислот
К 4 мг сухого гидролизата БР в 1 мл 2 Μ NaHCO 3 (рН 9 - 10) порциями при перемешивании добавляли 25.6 мг Dns-хлорида в 2 мл ацетона. Реакционную смесь выдерживали 1 ч при перемешивании при 40 ºС, подкисляли 2 н. HCl до рН 3.0 и экстрагировали этилацетатом (3 × 5 мл). Объединенный экстракт промывали дистиллированной Н 2 О до рН 7.0 (2 × 1 мл), сушили безводным Na 2 SO 4 , растворитель удаляли при 10 мм рт. ст. Выход 15.3 мг (78 %).
Синтез метиловых эфиров N-Dns-аминокислот
Для получения диазометана к 20 мл 40 % KОН в 40 мл диэтилового эфира добавляли 3 г влажной Ν -нитрозометилмочевины и перемешивали 15 20 мин на водяной бане со льдом. После окончания газовыделения эфирный слой отделяли, промывали дистиллированной Н 2 О до рН 7.0, сушили Na 2 SO 4 и использовали для обработки N-Dns-аминокислот. Выход 17.4 мг (69 %).
Биосинтез БР
Биосинтетический БР (выход 8 - 10 мг с 1 г биомассы) получен в синтетической среде, в которой протонированные фенилаланин, тирозин и триптофан были замещены их дейтерированными аналогами — L -[2,3,4,5,6-2H 5 ]фенилаланином, L -[3,5-2H 2 ]тирозином и L -[2,4,5,6,7-2H 5 ]трип-тофаном (г/л): D,L -аланин — 0.43; L -аргинин — 0.4; D,L -аспарагиновая кислота — 0.45; L -цистеин — 0.05; L -глутаминовая кислота — 1.3; L -глицин — 0.06; D,L -гистидин — 0.3; D,L -изолейцин — 0.44; L -лейцин — 0.8; L -лизин — 0.85; D,L -метионин — 0.37; D,L -фенилаланин — 0.26; L -пролин — 0.05; D,L -серин — 0.61; D,L -треонин — 0.5; L -тирозин — 0.2; D,L -триптофан — 0.5; D,L -валин — 1.0; АМФ — 0.1; УМФ — 0.1; NaCl — 250; MgSO 4 × × 7H 2 O — 20; KСl — 2; NH 4 Cl — 0.5; KNO 3 — 0.1; KH 2 PO 4 — 0.05; K 2 HPO 4 — 0.05; Na+-цитрат — 0.5; MnSO 4 . 2H 2 O — 3.10–4; CaCl 2 . 6H 2 O — 0.065; ZnSO 4 . 7H 2 O — 4 . 10–5; FeSO 4 . 7H 2 O — 5 . 10–4; CuSO 4 . 5H 2 O — 5 . 10–5; глицерин — 1.0; биотин — 1 . 10–4; фолиевая кислота — 1.5 . 10–4; витамин В12 — 2 . 10–5. Ростовую среду автоклавировали 30 мин при 0.5 атм; рН доводили 0.5 М KОН до 6.5 - 6.7. Выращивание бактерий проводили в колбах Эр-ленмейера, вместимостью 500 мл (объем реакционной смеси 100 мл) 4 - 5 сут. при 35 ºС в условиях интенсивной аэрации на орбитальном шейкере Biorad 380-S (Венгрия) и освещении монохромными лампами ЛДС-40 (3 лампы по 1.5 лк). Последующие операции по выделению БР проводили с использованием светозащитной лампы с оранжевым светофильтром ОРЖ-1.
Выделение фракции пурпурных мембран (ПМ)
Биомассу (1 г) промывали дистиллированной водой и осаждали центрифугированием (1500 g, 20 мин). Осадок суспендировали в 100 мл дистиллированной Н2О и выдерживали 3 ч при 4 ºС. Реакционную смесь центрифугировали (1500 g, 15 мин), осадок ресуспендировали в 20 мл дистиллированной Н2О и дезинтегрировали ультразвуком (22 кГц, 3 раза по 5 мин) в водяной бане со льдом (0 ºС). Клеточный гомогенат после промывки дистиллированной Н2О суспендировали в 10 мл буфера (125 мМ NaCl, 20 мМ MgCl2, 4 мМ Трис-HCl, рН 8.0), добавляли 5 мкг РНК Ι (2-3 ед. акт.) и инкубировали 2 ч при 37 ºС. Затем добавляли 10 мл того же буфера, выдерживали 10-12 ч при 4 ºС. Водную фракцию отделяли центрифугированием (1500 g, 20 мин), осадок ПМ обрабатывали 50 %-м этанолом (5 × 7 мл) при 4 ºС с последующим отделением растворителя. Процедуру повторяли трижды до получения бесцветных промывных вод.
Содержание белка в пробах определяли спектрофотометрически по соотношению D 280 / D 568 ( ε 280 =1.1 . 105 и ε 568 = 6.3 . 104 M–1 см–1 [13]). Регенерацию ПМ проводили, как описано в работе [14]. Выход фракции ПМ 120 мг (х. ч. 80 - 85 %).
Выделение БР
Фракцию ПМ (в Н 2 О) (1 мг / мл) растворяли в 1 мл 0.5 % ДДС-Na, инкубировали 5–7 ч при 37 ºС с последующим центрифугированием (1200 g, 15 мин). Осадок отделяли, к супернатанту добавляли дробными порциями метанол (3 раза по 100 мкл) при 0 ºС, выдерживали 14 - 15 ч при 4 ºС и центрифугировали при охлаждении (1200 g , 15 мин). Фракционирование проводили трижды, уменьшая концентрацию 0.5 % ДДС-Na до 0.2 и 0.1 %. Кристаллический белок (8 - 10 мг) промывали холодной дистиллированной 2Н2О (2 × 1 мл) и центрифугировали (1200 g, 15 мин).
Очистка БР
Пробу белка (5 мг) растворяли в 100 мкл буферного раствора и помещали на колонку размером 150 × 10 мм; неподвижная фаза — сефадекс G-200 ("Pharmaсia", США) (удельный объем упакованных гранул — 30 - 40 ед. на 1 г сух. сефадекса), уравновешенная буферным раствором, содержащим 0.1 %-й ДДС-Na и 2.5 мМ ЭТДА. Элюирование проводили 0.09 М трис-боратным буфером, содержащим 0.5 М NaCl с pH 8.35 ( I = 0.075) со скоростью 10 мл / (см2 ч). Объединенные белковые фракции подвергали лиофильной сушке, запаивали в стеклянные ампулы (10 × 50 мм) и хранили при –4 ºС.
Получение апомембран (АП)
Суспензию ПМ (50 мг) в 50 мл 1 М NH 2 OH (pН 6.0) выдерживали 10 ч при перемешивании на ледяной бане (4 ºС) при освещении реакционной смеси ксеноновой лампой. Осадок отделяли центрифугированием (1000 g, 10 мин), дважды промывали дистиллированной водой и центрифугировали. Фракцию АП суспендировали в 2 мл 5 мМ 2-(N-морфолино)-этансульфониламида в 100 мМ NaCl.
Регенерация АП с 13-транс-ретиналем
К 2 мл суспензии АП (2 . 10–5 М) в кварцевой кювете добавляли при перемешивании 0.1 мл 2 мМ раствора полностью 13-транс-ретиналя в метаноле и выдерживали 6–8 ч в темноте при 40 ºС. Степень регенерации ПМ определяли спек- трофотометрически по соотношению Dнат.280, Dнат.568 и Dрег.280, Dрег.568, где D280 и D568 — поглощение суспензии нативных и регенерированных ПМ при λ = 280 и λ = 568 нм.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Биосинтез БР
Выбор стратегии биосинтеза 2Н-меченного БР c использованием штамма экстремальной фотоор-ганогетеротрофной галобактерии Halobacterium halobium определялся целью исследования, связанной с изучением принципиальной возможности получения 2Н-меченных трансмембранных фотопреобразующих белков в микропрепаративных количествах для реконструкции искусственных мембран в 2Н 2 О и БР-содержащих нанопленок.
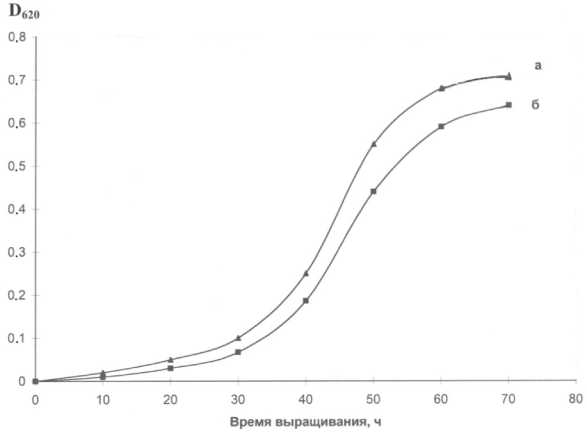
При выборе [2,3,4,5,6-2Н 5 ]фенилаланина, [3,5-2H 2 ]тирозина и [2,4,5,6,7-2H 5 ]триптофана в качестве источников дейтерия учитывалась их исключительная важность в гидрофобном взаимодействии молекулы БР с липидным бислоем клеточной мембраны, устойчивость к реакциям (1H-2H) обмена в водной среде в условиях выращивания штамма-продуцента, а также возможность применения метода высокочувствительной масс-спектрометрии ЭУ для последующего анализа. 2Н-меченный БР получен микробным синтезом штамма-продуцента фотоорганогетеротрофных галобактерий H. halobium при выращивании в синтетической среде (4.3 М NaCl) с [2,3,4,5,6-2Н 5 ]фе-нилаланином, [3,5-2H 2 ]тирозином и [2,4,5,6,7-2H 5 ]триптофаном. В оптимальных условиях выращивания H. halobium (период инкубации 4 - 5 сут., 35 ºС при освещении монохромным светом при λ = 560 нм) в клетках синтезировался каротиноид-содержащий фиолетовый пигмент, по спектральному соотношению белкового и хромофорного фрагментов молекулы D 280 / D 568 1.5 : 1.0 идентичный природному БР. Как показали результаты исследования, рост штамма на синтетической среде (рис. 2, б) ингибировался незначительно по сравнению с контролем (рис. 2, а) на протонированной среде, что существенно упрощает оптимизацию условий биосинтеза 2Н-меченного БР, заключающуюся в эквивалентной замене протонированных ароматических аминокислот их дейтерированными аналогами — [2,3,4,5,6-2Н 5 ]фенилаланином (0.26 г/л), [3,5-2H 2 ]тирозином (0.2 г/л) и [2,4,5,6,7-2H 5 ]триптофаном (0.5 г/л).
Основными этапами эксперимента являлись: выращивание штамма продуцента H. halobium на синтетической среде с [2,3,4,5,6-2Н 5 ]фенилалани-ном (0.26 г/л), [3,5-2H 2 ]тирозином (0.2 г/л) и [2,4,5,6,7-2H 5 ]триптофаном (0.5 г/л), отделение от клеточного содержимого, выделение фракции
D
300 400 500 600 700 нм
Рис. 2. Динамика роста штамма H. halobium в различных экспериментальных условиях.
Протонированная синтетическая среда (а); синтетическая среда с [2,3,4,5,6-2Н5]Phe (0.26 г/л), [3,5-2H 2 ]Tyr (0.2 г/л) и [2,4,5,6,7-2H 5 ]Trp (0.5 г/л) (б). Период инкубации 4– 5 сут., 35 ºС, освещение монохромным светом при λ = 560 нм
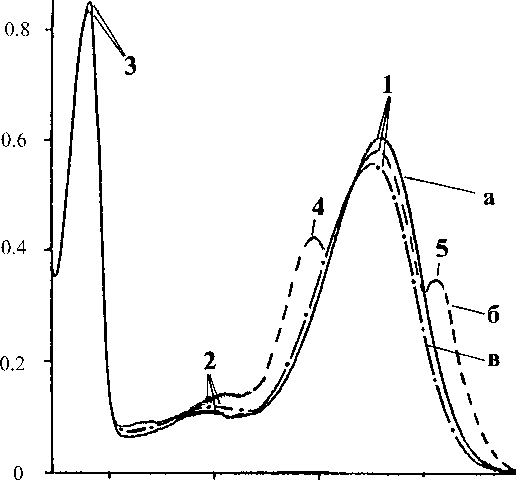
Рис. 3. Спектры поглощения ПМ (в 50 %-м этаноле) на различных стадиях обработки.
Природный БР (а); ПМ после промежуточной обработки (б); очищенные от посторонних каротиноидов ПМ (в). Полоса 1 — спектральная форма БР568, 2 — примесь спектральной формы мета-бактериородопсина М412, 3 — полоса поглощения ароматических аминокислот, 4 и 5 — посторонние каротиноиды. В качестве контроля использовали природный БР пурпурных мембран (ПМ), отделение низко- и высокомолекулярных примесей, клеточной РНК, пигментов (преимущественно каротиноидов) и липидов, фракционирование солюбилизированного в 0.5 % SDS-Na белка метанолом, ГПХ на сефадексе G-200, электрофорез в 12.5 %-м ПААГ с 0.1 %-м ДДС-Na. Поскольку белок локализуется в ПМ, освобождение от низкомолекулярных примесей и внутриклеточного содержимого достигали осмотическим шоком клеток дистиллированной водой на холоде после удаления 4.3 М NaCl и последующим разрушением клеточной оболочки ультразвуком при 22 кГц. Для разрушения клеточной РНК проводили обработку клеточного гомогената РНК-азой Ι (2-3 ед. акт.). Фракция ПМ наряду с искомым белком в комплексе с липидами и полисахаридами содержала примесь связанных каротиноидов и посторонних белков. Поэтому применялись специальные методы фракционирования белка без повреждения его нативной струк- туры и диссоциации, что требовало применения специальных методов очистки от каротиноидов и липидов и последующей колоночной гель-проникающей хроматографии на сефадексе G-200. Очистка от каротиноидов, заключающаяся в многократной обработке ПМ 50 %-м этанолом при 4 ºС, являлась рутинным, но обязательным этапом, несмотря на значительные потери хромопротеина. Использовалось не менее пяти обработок 50 %-м этанолом, чтобы получить спектр поглощения суспензии ПМ, очищенный от каротиноидов 4 и 5 (степень хроматографической чистоты 80-85 %), показанный на рис. 3, б и в, на различных стадиях обработки относительно природного БР (рис. 3, а). Образование ретинальпротеинового комплекса в молекуле БР приводит к батохромному сдвигу в спектре поглощения ПМ (рис. 3, в): основная полоса (1) при максимуме поглощения λ = 568 нм, вызванная световой изомеризацией хромофора по связи С13=С14, определяется наличием 13-транс-ретинального остатка ретиналя БР568, дополнительная малоинтенсивная полоса 2 при λ = 412 нм характеризует незначительную примесь образующейся на свету спектральной формы мета-бактериородопсина M412 c депротонированной альди-минной связью между остатком 13-транс-ретиналя и белком, а полоса 3 при λ = 280 нм определяется поглощением ароматических аминокислот в поли-пептидной цепи белка (для чистого БР соотношение D280 / D568 равно 1.5 : 1.0).
Выделение и очистка БР
Следующим необходимым этапом являлось фракционирование и хроматографическая очистка белка. Поскольку БР, будучи трансмембранным белком (Мr 26.7 кДа), пронизывает билипидный слой в виде семи α-спиралей, применение сульфата аммония и других традиционных высаливающих агентов не дает положительного результата. Решение проблемы заключалось в переводе белка в растворимую форму солюбилизацией в 0.5 %-м ДДС-Na. Использование ионного детергента ДДС-Na диктовалось необходимостью максимальной солюбилизации белка в нативной биологически-активной форме в комплексе с 13-транс-ретина-лем с комбинированием стадии делипидизации и осаждения, поскольку солюбилизированный в слабоконцентрированном растворе ДДС-Na (0.5 %) БР, сохраняет спиральную α-конфигурацию [15]. Поэтому отпала необходимость использования органических растворителей — ацетона, метанола и хлороформа для очистки от липидов, а делипиди-зация и осаждение белка совмещались в одну стадию, что существенно упрощало дальнейшее фракционирование. Значительным преимуществом метода является то, что выделяемый белок в комплексе с молекулами липидов и детергента распределяется в надосадочной жидкости, а другие высокомолекулярные примеси — в непрореагировавшем осадке, легко отделяемом центрифугированием. Фракционирование солюбилизован-ного в 0.5 %-м растворе ДДС-Na белка с его последующим выделением в кристаллическом виде достигали в три стадии дробным низкотемпературным (4 ºС) осаждением метанола, уменьшая концентрацию детергента соответственно в 2.5 и 5 раз. Окончательная стадия очистки БР заключалась в отделении белка от низкомолекулярных примесей гель-проникающей хроматографией, для чего БР-содержащие фракции дважды пропускали через колонку с декстрановым сефадексом G-200, уравновешенным 0.09 М трис-боратным буфером (рН 8.35) с 0.1 %-м ДДС-Na и 2.5 мМ ЭТДА. Согласно разработанному методу, получено 8-10 мг 2Н-меченного БР из 1 г бактериальной биомассы, гомогенность которого удовлетворяла требованиям, предъявляемым для реконструкции мембран, и подтверждалась электрофорезом в 12.5 %-м ПААГ с 0.1 %-м ДДС-Na, регенерацией апомембран с 13-транс-ретиналем и ОФ ВЭЖХ метиловых эфиров N-Dns-аминокислот. Небольшой выход БР не был препятствием для последующего масс-спектрометрического анализа, однако необходимо подчеркнуть, что для обеспечения высокого выхода белка нужно наработать большее количество сырьевой биомассы.
Условия проведения гидролиза 2Н-меченного БР определялись необходимостью предотвращения реакций изотопного (1Н-2Н) обмена водорода на дейтерий в молекуле фенилаланина и сохранения остатков триптофана в белке. Рассматривались два альтернативных варианта — кислотный и щелочной гидролиз. Кислотный гидролиз белка в стандартных условиях (6 н. HСl или 8 н. H 2 SO 4 , 110 ºС, 24 ч) приводит к полному разрушению триптофана и частичному разрушению серина, треонина и некоторых других аминокислот в белке [16], которые для настоящего исследования не играют существенной роли. Модификация этого метода, заключающаяся в добавлении в реакционную среду фенола, тиогликолевой кислоты, β -меркапто-этанола, позволяет сохранить до 80 85 % триптофана [17]. Использование п-толуол сульфокислоты с 0.2 % 3-(2-аминоэтил)-индолом или 3 М меркаптоэтансульфокислоты также эффективно для сохранения триптофана (до 93 %) [18]. Однако для решения поставленной задачи вышеперечисленные методы непригодны, поскольку обладают существенным недостатком: в условиях кислотного гидролиза с высокой скоростью происходит изотопный (1Н-2Н) обмен ароматических протонов (дейтеронов) в молекулах триптофана,
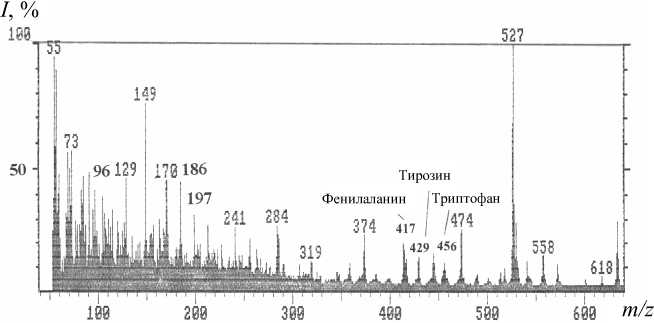
Рис. 4. Полный масс-спектр ЭУ метиловых эфиров N-Dns-[2H]производных ароматических аминокислот, выделенных из гидролизата БР из H. halobium (70 эВ, ускоряющее напряжение 8 кВ, 180–200 ºС).
Условия получения: синтетическая среда с [2,3,4,5,6-2H 5 ]Phe, [3,5-2H 2 ]Tyr и [2,4,5,6,7-2H 5 ]Trp. Условия гидролиза БР: 4 н. Ba(OH) 2 (в 2Н 2 О), 110 ºС, 24 ч. Пики молекулярных ионов аминокислот соответствуют их производным. I — интенсивность пиков (%)
тирозина и гистидина, а также протонов при атоме С3 аспарагиновой и С4 глутаминовой кислот [19]. Поэтому даже проведение гидролиза в дейтерированных реагентах (6 н. 2HCl, 4 н. 2H 2 SO 4 в 2H 2 O) не позволяет получать реальные данные о включении дейтерия в белок.
В условиях щелочного гидролиза (4 н. Ba(OH) 2 или 4 н. NaOH, 110 ºC, 24 ч) реакций изотопного (1Н-2Н) обмена практически не наблюдается (исключением является протон (дейтерон) у атома С2 гистидина, а триптофан не разрушается, что определило выбор этого метода гидролиза в настоящей работе. Упрощение процедуры выделения смеси свободных аминокислот за счет нейтрализации H 2 SO 4 (в 2H 2 O) явилось причиной выбора в качестве гидролизующего агента 4 н. Ba(OH) 2 . Возможная D , L -рацемизация аминокислот при щелочном гидролизе не влияла на результат последующего масс-спектрометрического исследования уровня дейтерированности молекул аминокислот.
Исследование уровня дейтерированности БР масс-спектрометрией ЭУ
Для изучения включения [2Н]ароматических кислот в молекулу БР использовали метод масс-спектрометрии ЭУ после модификации смеси аминокислот гидролизата БР в метиловые эфиры N-Dns-производных аминокислот. Чтобы получить воспроизводимый результат по уровню дей-терированности 2Н-меченного белка, сначала регистрировали полный скан масс-спектра ЭУ смеси метиловых эфиров N-Dns-производных [2Н]амино-кислот, полученных из гидролизата БР. Уровень дейтерированности рассчитывали по пикам молекулярных ионов [М]+ производных. Затем проводили разделение метиловых эфиров N-Dns-[2Н]производных ароматических аминокислот обращенно-фазовой ВЭЖХ и получали масс-спектры ЭУ для каждой выделенной индивидуальной [2Н]аминокислоты.
Полный масс-спектр электронного удара ЭУ смеси метиловых эфиров N-Dns-производных аминокислот, показанный на рис. 4 (сканирование при m/z 50 - 640, базовый пик m/z 527, 100 %), характеризуется непрерывностью: пики в интервале m/z от 50 до 400 на шкале массовых чисел представлены фрагментами метастабильных ионов, низкомолекулярных примесей, а также продуктами химической модификации аминокислот. Анализируемые [2Н]ароматические аминокислоты, занимающие шкалу массовых чисел m/z от 415 до 456, представлены смесями молекул с различным количеством включенных атомов дейтерия, поэтому молекулярные ионы [М]+ полиморфно расщеплялись на отдельные кластеры со статистическим набором значений m/z в зависимости от количества водородных атомов в молекуле. Учитывая эффект изотопного полиморфизма, подсчет уровня дейтерированности молекул [2Н]аминокис-лот проводили по наиболее распространенному пику молекулярного иона [М]+ в каждом кластере с математически усредненной величиной [М]+ (рис. 4) — для фенилаланина пик молекулярного
Величины пиков молекулярных ионов [М]+ в масс-спектре ЭУ метиловых эфиров N-Dns-[2,3,4,5,6-2H5]Phe, N-Dns-[3,5-2H2]Tyr и N-Dns-[2,4,5,6,7-2H5]Trp и уровни дейтерированности
|
Соединение |
Величина пика [М]+ |
Интенсивность, % |
Количест во атомов * дейтерия |
Уровень дейтери-рованности, % от общего количества атомов водорода** |
|
N-Dns- |
413 |
7 |
1 |
13 |
|
[2,3,4,5,6- 2H 5 ]Phe-OMe |
414 |
18 |
2 |
25 |
|
415 |
15 |
3 |
38 |
|
|
416 |
11 |
4 |
50 |
|
|
417 |
14 |
5 |
63 |
|
|
418 |
6 |
6 |
75 |
|
|
N-Dns-[3,5- |
428 |
12 |
— |
— |
|
2H 2 ]Tyr-OMe |
429 |
15 |
1 |
14 |
|
430 |
5 |
2 |
29 |
|
|
N-Dns- |
453 |
5 |
2 |
26 |
|
[2,4,5,6,7- 2H 5 ]Trp-OMe |
454 |
6 |
3 |
38 |
|
455 |
9 |
4 |
50 |
|
|
456 |
11 |
5 |
64 |
|
|
457 |
5 |
6 |
77 |
Примечание.
* — прочерк означает отсутствие включения дейтерия; ** — при подсчете уровня дейтерированности протоны (дейтероны) при карбоксильных СООН- и амино NH 2 -группах аминокислот не учитывались из-за легкости изотопного (1Н-2Н) обмена.
иона определялся [М]+ при m/z 417, 14 % (вместо [М]+ при m/z 412, 20 % для немеченного производного (пики немеченных аминокислот не показаны)), тирозина — [М]+ при m/z 429, 15 % (вместо [М]+ при m/z 428, 13 %), триптофана — [М]+ при m/z= = 456, 11 % (вместо [М]+ при m/z 451, 17 %). Уровень дейтерированности, соответствующий увеличению молекулярной массы, составил для [2H]тирозина — 2 (96 ат.% 2Н), [2H]фенилала-нина — 5 (90 ат. % 2Н) и [2H]триптофана —5 (98 ат.% 2Н) атомов дейтерия, что совпадает с данными по уровню дейтерированости исходных ароматических аминокислот — [3,5-2H 2 ]Tyr, [2,3,4,5,6-2H 5 ]Phe и [2,4,5,6,7-2H 5 ]Trp, добавляемых в ростовую среду штамма продуцента. Полученные данные по уровню дейтерированности
[2H]фенилаланина, [2H]тирозина и [2H]триптофана позволяют сделать вывод о высокой селективности включения [2H]ароматических аминокислот в молекулу БР: дейтерий детектировался во всех остатках [2H]ароматических аминокислот (таблица). Присутствие в масс-спектре ЭУ пиков [M]+ протонированных и полудейтерированных аналогов фенилаланина с [M]+ при m/z 413 - 418, тирозина с [M]+ при m/z 428 - 430 и триптофана с [M]+ при m/z 453 - 457 с различными вкладами в уровни дей-терированности молекул, свидетельствует о сохранении небольшой доли минорных путей биосинтеза de novo , приводящим к разбавлению дейтериевой метки и определяется условиями биосинтеза 2Н-меченного БР (таблица).
а
б
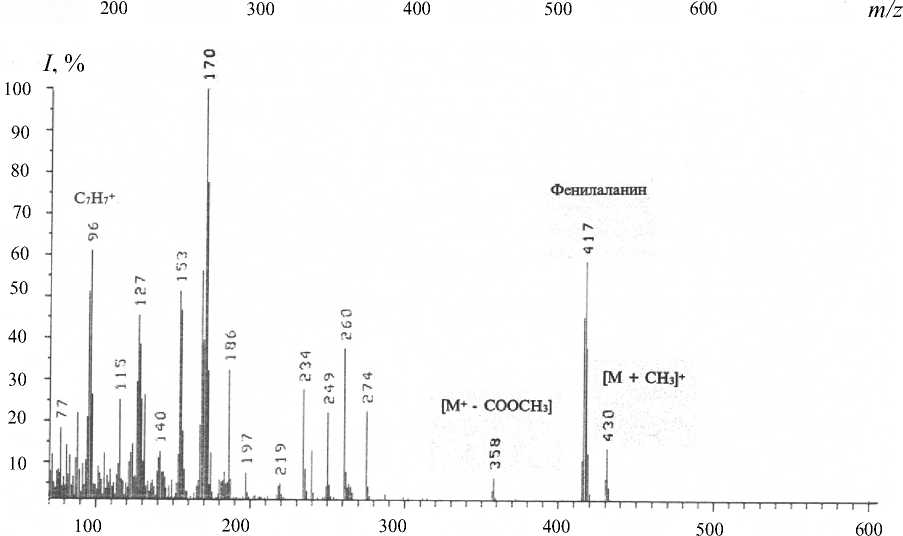
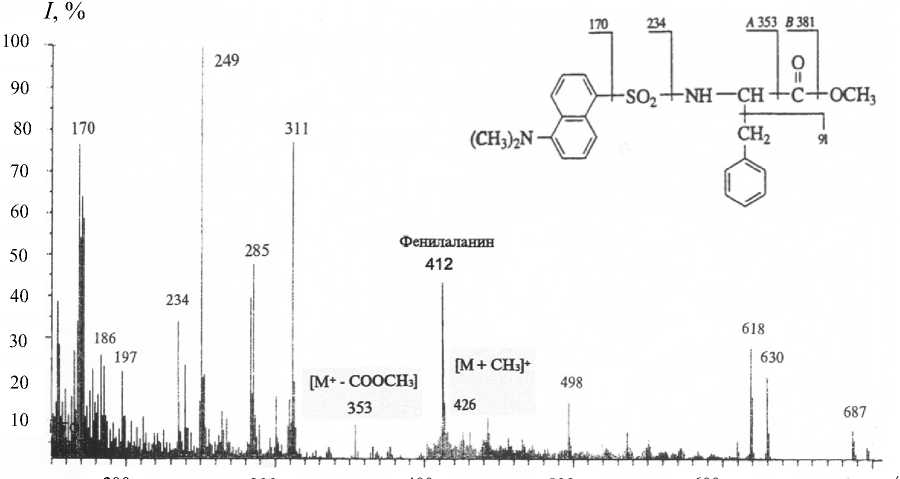
Рис. 5. Масс-спектры ЭУ метилового эфира N-Dns-[2,3,4,5,6-2H 5 ]Phe в различных экспериментальных условиях.
а — немеченный метиловый эфир N-Dns-Phe; б — метиловый эфир N-Dns-[2,3,4,5,6-2H5]Phe, выделенный из гидролизата БР методом ОФ ВЭЖХ. Условия разделения: жидкостной хроматограф Knauer ("Knauer", ФРГ), неподвижная фаза — сепарон С18 (Kova, Cловакия), колонка 250 × 10 мм; элюент: (А) — CH 3 CN—CF 3 COOH (100 : 0.1–0.5 об.%) и (Б) — CH 3 CN (100 об.%)
m/z
Согласно данным масс-спектрометрического анализа, пики молекулярных ионов [М]+ метиловых эфиров N-Dns-[2H]-производных ароматических аминокислот обладали низкой интенсивностью и полиморфно расщеплялись, поэтому области их молекулярного обогащения были сильно уширены.
Кроме этого, масс-спектры компонентов смеси аддитивны, поэтому смеси можно анализировать, только если имеются спектры различных компонентов, записанные в тех же условиях. Проводимые вычисления предусматривают решение системы из n уравнений с n неизвестными для смеси из n компонентов. Для компонентов, концентрация которых превышает 10 мол. %, правильность и воспроизводимость результатов анализа составляет ± 0.5 мол. % (при доверительной вероятности 90 %). Поэтому для получения воспроизводимого результата необходимо хроматографически выделять индивидуальные производные [2Н]амино-кислот из белкового гидролизата. Для решения поставленной задачи использовали ОФ ВЭЖХ на октадецилсилановом селикагеле силасорб С18, эффективность которого подтверждалась разделением смеси метиловых эфиров N-Dns-произ-водных [2Н]аминокислот из других микробных объектов, таких как метилотрофные бактерии и микроводоросли [20]. Метод адаптировали к условиям хроматографического разделения смеси метиловых эфиров N-Dns-[2H]-производных аминокислот гидролизата БР, что заключалось в оптимизации соотношения элюентов, форме градиента и скорости элюирования с колонки. Наилучшее разделение достигалось при градиентном элюировании метиловых эфиров N-Dns-[2H]-производных аминокислот смесью растворителей CH3CN— CF3COOH (100 : 0.1–0.5 об. %). При этом удалось разделить триптофан и трудно разрешимую пару фенилаланин / тирозин. Степени хроматографической чистоты выделенных из смеси белкового гидролизата БР метиловых эфиров N-Dns-[2,3,4,5,6-2H5]фенилаланина, N-Dns-[3,5-2H2]тиро-зина и N-Dns-[2,4,5,6,7-2H5]триптофана составили 89, 91 и 90 % соответственно. Полученный результат подтвердил рис. 5, б, на котором приведен масс-спектр ЭУ метилового эфира N-Dns-[2,3,4,5,6-2H5]фенилаланина, выделенного ОФ ВЭЖХ (сканирование при m/z 70-600, базовый пик m/z 170, 100 %) (масс-спектр приведен относительно немеченного метилового эфира N-Dns-фенилаланина (рис. 5, а), сканирование при m/z= = 150-700, базовый пик m/z 250, 100 %). Доказательством включения дейтерия в молекулу фенилаланина является пик тяжелого молекулярного иона метилового эфира N-Dns-[2H]фенилаланина ([М]+ m/z 417, 59 % вместо [М]+ m/z 412, 44 % для немеченного производного фенилаланина) и дополнительный пик обогащенного дейтерием бензильного С6Н5СН2-фрагмента молекулы [2H]фе-нилаланина m/z 96, 61 % (вместо m/z 91, 55 % в контроле (не показан)) (рис. 5, б). Пики второстепенных фрагментов различной интенсивности со значениями m/z 249, 234 и 170 являются продуктами вторичного распада дансильного остатка до N-диметиламинонафталина, низкоинтенсивный пик [M - COOCH3]+ m/z 358, 7 % (m/z 353, 10 %, контроль) является продуктом отщепления кар-боксиметильной СООСН3-группы из метилового эфира N-Dns-[2H]фенилаланина, а пик [M + CH3]+ m/z 430, 15 % (m/z 426, 8 %, контроль) — продуктом дополнительного метилирования по α-амино-группе [2H]фенилаланина (рис. 5, б). Согласно данным масс-спектрометрии ЭУ, разница между молекулярной массой легкого и тяжелого пиков [M]+ метилового эфира N-Dns-фенилаланина составляет пять единиц, что совпадает с первичными данными по уровню дейтерированности исходного [2,3,4,5,6-2H5]фенилаланина, добавляемого в среду выращивания. Масс-спектрометрические данные по уровням дейтерированности [3,5-2H2]тирозина и [2,4,5,6,7-2H5]триптофана также находятся в корреляции и подтверждены спектроскопией 1Н ЯМР.
ЗАКЛЮЧЕНИЕ
Полученные экспериментальные данные свидетельствуют о высокой эффективности мечения БР дейтерием по остаткам ароматических аминокислот Phe, Tyr и Trp с выходом [2H]БР 8–10 мг на 1 г биомассы. Главным преимуществом метода является то, что выделенный [2Н]БР сохраняет природную биологически активную конфигурацию в комплексе с 13- транс -ретиналем и способность к фотохимическим превращениям in vitro , что было доказано электрофорезом в 12.5 %-м ПААГ с 0.1 %-м ДДС-Na и регенерацией апомембран с 13- транс -ретиналем. Метод также применим к получению других аналогичных БР трансмембранных белков галофильных бактерий — сенсородопсина и гало-родопсина. Уникальные свойства природных бактериородопсинов обеспечивают широкий диапазон био- и нанотехнологических приложений, в которых могут найти применение их 2Н-меченные аналоги. В дальнейшем планируется получать полностью дейтерированные препараты БР для реконструкции функционально активных систем трансмембранных белков в тяжелой воде с очищенными [2Н]жирными кислотами и другими биологически активными соединениями. Эти исследования позволят дать ответ на вопрос, как функционирует БР в составе нативных мембран в условиях полного замещения протонов на дейтерий.
Работа осуществлялась при поддержке Научноисследовательского центра медицинской биофизики (Болгария). Грант № 012.
