Численно-размерный состав мышечных волокон передней большеберцовой мышцы при регенерации пересеченного седалищного нерва собак в условиях нейтрального и дистракционного остеосинтеза

Автор: Щудло Н.А., Филимонова Г.Н., Панасенко С.В., Виноградова О.А.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 3, 2007 года.
Бесплатный доступ
Изучены изменения численно-размерного состава мышечных волокон передней большеберцовой мышцы беспородных собак при разных условиях регенерации седалищного нерва: после его пересечения и микрохирургического шва (группа I), при сочетании нейротомии с остеотомией бедренной кости в условиях нейтрального остеосинтеза (группа II) и дистракционного остеосинтеза (группа III). Через 4 месяца после операции в группе I распределение мышечных волокон по диаметру сопоставимо с интактной мышцей. В группе II асимметрия гистограмм распределения свидетельствует о сохраняющейся атрофии мышечных волокон в мышце оперированной конечности и их компенсаторной гипертрофии в контралатеральной. Изменения объёмной доли эндоневрия, численной плотности мышечных волокон и микрососудов передней большеберцовой мышцы у животных III серии в сравнении с интактными указывают на усиление васкуляризации и интенсивную пластическую перестройку мышечных волокон в опытах с дистракционным остеосинтезом.
Нерв, регенерация, мышца, нейтральный и дистракционный остеосинтез
Короткий адрес: https://sciup.org/142121022
IDR: 142121022
The numerical-and-sized composition of the muscular fibers of the anterior tibial muscle for regeneration of canine crossed sciatic nerve under the condition of neutral and distraction osteosynthesis
The changes of numerical-and-sized composition of the muscular fibers of the anterior tibial muscle have been studied in mongrel dogs under different conditions of sciatic nerve regeneration: after its crossing and microsurgical suturing (gorup I), in case of neurotomy combined with femoral osteotomy under neutral osteosynthesis (group II) and distraction osteosynthesis (group III). In group I by 4 months after surgery the distribution of muscle fibers by diameter is comparable with anintact muscle. In group II the asymmetry of distribution histograms demonstrates the remaining atrophy of muscular fibers in the muscle of the limb operated and their compensatory hypertrophy in the contralateral limb. The changes in endoneurium volumetric part, those in the numerical density of muscle fibers and microvessels of the anterior tibial muscle in the animals of series III comparing with the intact ones show vascularization increase and the intensive plastic reorganization of muscle fibers in the experiments where distraction osteosynthesis is used.
Текст научной статьи Численно-размерный состав мышечных волокон передней большеберцовой мышцы при регенерации пересеченного седалищного нерва собак в условиях нейтрального и дистракционного остеосинтеза
При сочетании открытого перелома кости с дефектом нерва некоторые авторы считают рациональным вместо аутонейропластики выполнить нейрорафию конец в конец, для чего необходимо укорочение кости [6, 10, 12]. Восстановление длины укороченного сегмента не осуществляют [6, 12] или выполняют через очень длительный срок после первой операции [10] из-за представлений об отрицательном влиянии дистракции на регенерацию нервных волокон. Экс- периментальным исследованиям регенерации сшитого нервного ствола в условиях дистракции посвящены немногочисленные работы [3, 7, 8].
Цель исследования – сопоставительный анализ морфофункциональных характеристик мышц голени собак при регенерации седалищного нерва после его пересечения и шва, а также при сочетании его повреждения с остеотомией бедренной кости и дистракционным остеосинтезом.
МАТЕРИАЛ И МЕТОДЫ
Эксперименты проведены на 10 взрослых бес- группе опытов (n=2) под внутривенным барбиту- породных собаках с длиной голени 10-12 см. В I ровым наркозом выполнено пересечение седа- лищного нерва на уровне средней трети бедра и микрохирургический эпи-периневральный шов нитью 8/0 с применением оптического увеличения микроскопа Opton. Во II (n=2) и III (n=6) группах аналогичная операция на нерве произведена после открытой чрездиафизарной остеотомии бедренной кости пилой Джигли и остеосинтеза спицестержневым аппаратом, причём в одном из опытов III группы вместо нейротомии и остетомии выполняли резекцию нерва и диафиза кости (дефект 2 см 18 % длины бедра). Во II группе фрагменты бедренной кости фиксировали в аппарате в течение 42 и 47 дней до момента консолидации. В III группе начиная с 7-9 дня после операции производили дистракцию с суточным темпом 0,5 и 0,75 мм за 2-3 приёма до восстановления исходной длины бедра. После консолидации дистракционного костного регенерата аппарат снимали. Через 4 месяца после операции животные были эвтана-зированы. Содержание, кормление животных, операции, а также выведение из опыта осуществляли согласно приказу МЗ СССР № 775, 1977 г.
Из верхней, средней и нижней трети передней большеберцовой мышцы иссекали фрагменты размером 0,7 мм3, погружали в жидкий азот и изготавливали поперечные криостатные срезы (10,0 мкм, t -20 ºС). Для выявления мышечных волокон и мик- рососудов ставили гистохимическую реакцию на миозиновую АТФ-азу по Padycula, Herman в модификации З. Лойда [4]. Условным контролем служили мышцы контралатеральной конечности, а также мышцы интактных собак. Количественную оценку васкуляризации и стромально-паренхиматозных отношений проводили методом стереологического анализа с использованием проекционного микроскопа «Визопан» фирмы Reichert Jung (Австрия) с тестовой решеткой коротких отрезков [1]. Статистическую обработку первичных данных проводили в «Секторе аналитико-статистических исследований и математического моделирования» РНЦ «ВТО» им. акад. Г.А. Илизарова: достоверность различий вычисляли по критерию Манна-Уитни (двусторонний вариант) с уровнем значимости 0,05 [5]. Используя аппаратно-программный комплекс "ДиаМорф" (Россия), в программе "Медиас" в оцифрованных изображениях поперечных срезов исследуемой мышцы (увеличение 200 ) измеряли диаметры 300-700 мышечных волокон, строили гистограммы их распределения с шагом 10,0 мкм. Оценку статистической значимости различий проводили с использованием критерия Вилкоксона, вычисления теста выполнены с использованием программы Atte Stat.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Опорная функция оперированной конечности и походка животных существенно различались в зависимости от характера травмы и оперативного вмешательства. В первой группе ослабление опорной функции и характерные для повреждений седалищного нерва денервационные проявления (трофические расстройства, опора не на подушечки, а на дорзальную поверхность пальцев в положении подошвенной гиперфлексии плюснефаланговых и межфаланговых суставов, периодическое "запрокидывание" стопы с гиперэкстензией в скакательном суставе) к 4 месяцам после операции компенсировались, что в значительной мере было связано с успешно протекающим процессом реиннервации.
Во II группе более тяжёлая травма – сочетание перерезки седалищного нерва и остеотомии приводила к аналогичным, но более продолжительным нарушениям опорно-двигательной функции конечности. Однако к концу эксперимента у одной собаки наблюдалось функциональное выздоровление, а у другой сохранялось только ослабление опоры на оперированную лапу.
В III группе опытов через 4 месяца после операции все 6 собак приступали на оперированную конечность, при этом у 5 собак с остеотомией и удлинением бедренного сегмента она находилась в положении отведения. Нормальный рисунок походки восстановился у собаки, которой была выполнена резекция нерва и кости и у которой по окончании периода дистракции достигнуто равенство длины конечностей. Клинически выраженная атрофия мышц голени к концу эксперимента наблюдалась у 2 собак из 6 (периметр голени на уровне выпуклостей брю-шек икроножной мышцы оперированной конечности был меньше соответствующего показателя контралатеральной примерно на 20 %).
Анализ гистохимических препаратов передней большеберцовой мышцы показал, что у интактных собак диаметры мышечных волокон варьируют от 10 до 90 мкм (рис. 1), их распределение близко к нормальному, количество классов гистограммы – 8, наибольшее число волокон принадлежит классу 41-50 мкм (31,18 %) (усредненная гистограмма). У некоторых интактных животных этот пик сглажен – то есть, представительство волокон в 3, 4, 5 классах гистограммы практически одинаковое и составляет 23-24 % (индивидуальная гистограмма).
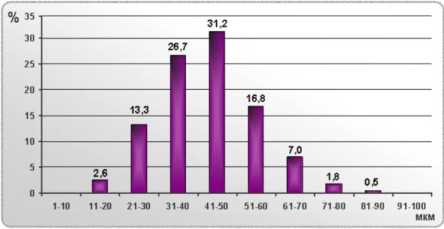
Рис. 1. Гистограмма распределения мышечных волокон по диаметрам в интактной группе
В I группе пик гистограммы распределения мышечных волокон по диаметрам у одной собаки смещён на 1 класс влево по сравнению с интактной, у второй совпадает с интактной (рис. 2). В отличие от интактных собак индивидуальные и усреднённая гистограммы в этих опытах шире на 1 класс за счёт представительства небольшого количества мышечных волокон в классе 1-10 мкм. Как ширина, так и форма индивидуальных и усреднённых гистограмм мышц оперированной и контралатеральной конечностей практически совпадают.
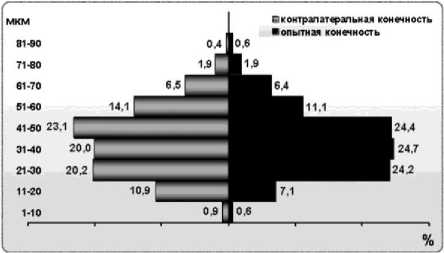
Рис. 2. Гистограммы распределения мышечных волокон по диаметрам через 4 месяца после нейротомии и шва седалищного нерва
Во II экспериментальной группе в опытной конечности диаметры мышечных волокон передней большеберцовой мышцы распределены по Гауссу. Так же, как в контроле I, присутствует небольшое количество волокон в размерном диапазоне 1-10 мкм. Наибольшее число волокон в мышце оперированной конечности принадлежит классу 31-40 мкм (34,89 %), контралатеральной – 41-50 мкм (35,26 %) (рис. 3). То есть, в мышце оперированной конечности по сравнению с интактной пик распределения мышечных волокон смещён на 1 класс влево, а в контралатеральной занимает положение, характерное для интактной мышцы. Однако в гистограммах мышц и оперированной, и контралатеральной конечностей количество классов уменьшено до 7, поскольку волокон диаметром более 70 мкм нет.
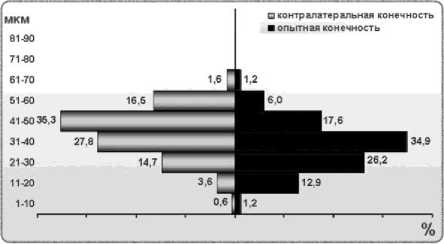
Рис. 3. Гистограммы распределения мышечных волокон по диаметрам при регенерации седалищного нерва в условиях нейтрального остеосинтеза
В III экспериментальной группе 6 исследованных случаев разделены на 3 подгруппы, в которых существенно различаются гистограммы распределения мышечных волокон по диаметрам. В первой подгруппе диаметры мышечных волокон варьируют от 1 до 81 мкм, гистограммы оперированной и контралатеральной конечностей состоят из 8 классов. По сравнению с интактной мышцей, в мышце оперированной конечности пик гистограммы смещён на 2 класса влево, а в контралатеральной наоборот – на 1 класс вправо (рис. 4, а). На оперированной стороне наибольшее число волокон в классе 2130 мкм (42,17 %), на противоположной – в классе 51-60 мкм (33,25 %).
Во второй подгруппе III группы характер распределения мышечных волокон в мышцах оперированной и контралатеральной конечностей также различен, в размерном диапазоне 110 мкм волокон нет (рис. 4, б). Волокна с диаметром более 70 мкм отсутствуют, поэтому гистограмма состоит из 6 классов – меньше, чем в контрольных группах. В мышце оперированной конечности по сравнению с интактной пик смещён на 2 класса влево, максимальное число волокон в диапазоне 21-30 мкм (38,52 %). В контралатеральной конечности диаметры волокон распределены по Гауссу, максимум принадлежит классу 31-40 мкм (33,43 %).
Экспериментальный случай с резекцией кости и нерва (№ 2403) имеет свои особенности. В мышце оперированной конечности пик распределения мышечных волокон приходится на классе 41-50 мкм (так же, как в интактной мышце), характер распределения нормален, но почти отсутствуют волокна диаметром меньше 30 мкм, число классов равно пяти (рис. 4, в). В контралатеральной конечности наблюдается смещение пика распределения мышечных волокон влево на 1 класс по сравнению с интактной мышцей: максимальное число волокон в диапазоне 31-40 мкм (31,74 %). При этом насчитывается 9 классов гистограммы, так как некоторые волокна превышают 90 мкм в диаметре.
Наряду с отличиями численно-размерного состава мышечных волокон III группа значительно разнится с интактной стромальнопаренхиматозным соотношением и численными параметрами. В передней большеберцовой мышце животных III группы средний диаметр мышечных волокон уменьшен на 20 % – настолько же повышена объёмная плотность эн-домизия. При этом численная плотность кровеносных сосудов увеличена в среднем на 70 %, а численная плотность мышечных волокон – на 50 %.
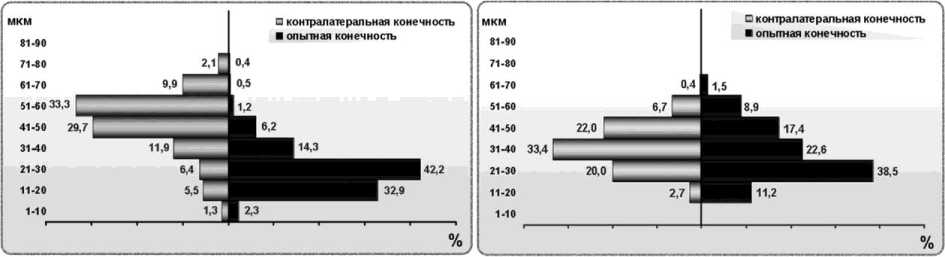
б
а
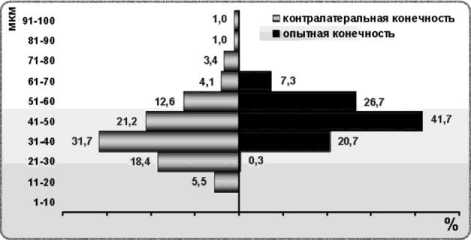
в
Рис. 4. Гистограммы распределения мышечных волокон по диаметрам при регенерации седалищного нерва в условиях дистракционного остеосинтеза: а подгруппа 1; б подгруппа 2; в подгруппа 3
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Модель пересечения седалищного нерва, а не резекции выбрана нами потому, что у животных с короткими конечностями резекция даже небольшого участка нерва приводит к несоответствию пучковых паттернов его отрезков. Степень несоответствия определяется индивидуальными особенностями внутристволового строения нервов, что затрудняет сравнительную оценку регенерации.
Полный анатомический перерыв (нейротмезис) седалищного нерва относится к числу тяжёлых повреждений, после которых восстановление чувствительности и движений редко бывает полным. Степень восстановления иннервируемых повреждённым нервом мышц определяется как параметрами регенерации нервных волокон, так и денервационными процессами, происходящими в мышцах.
Восстановление сократительной функции мышц проблематично, если численность регенерирующих аксональных отпрысков, реиннервирующих денервированные моторные бляшки, недостаточна или если при интенсивной невротизации нервно-мышечных соединений выражены явления кросс-иннервации (иннервации мышечных волокон определённого типа несоотвующими нервными волокнами). Двигательная реиннервация может быть скомпрометирована сенсорным дефицитом – прежде всего недостаточностью про-приорецепции. И наконец – выраженный внутримышечный фиброз, развивающийся в течение периода денервации, снижает эффективность вызываемых нервными импульсами мышечных сокращений на этапе реиннервации.
Оценка численно-размерного состава мышечных волокон и мышечно-стромального отношения в опытах с регенерацией нерва отражает качество реиннервации. Денервация приводит к атрофии мышечных волокон. Они уменьшаются в размерах и разделяются широкими соединительнотканными прослойками, которые, по мнению некоторых авторов, увеличиваются за счёт интенсивной пролиферации фибробластов [9]. Считается, что явления гибели мышечных волокон наблюдаются в поздние сроки денервации более 6 месяцев [9]. В обзоре Ch. Rehfeldt et al. [11] цитируются работы о влиянии физической активности на численность мышечных волокон: бездействие мышцы приводит к снижению численности, но активирующие миосателлитоциты стимулы способствуют эффективному возмещению ее потери.
По-видимому, только выполнение первичного шва нерва позволяет предотвратить грубые нарушения архитектоники мышечных волокон. Как показывают результаты первой серии опытов, через 4 месяца после первичного шва седалищного нерва у собак наступает клиническое функциональное выздоровление. Численно-размерный состав мышечных волокон реиннервированной передней большеберцовой мышцы мало отличается от интактной, что наглядно подтверждает оптимистические представления об обратимости денервационной атрофии.
Сочетание перерезки седалищного нерва с остеотомией бедренной кости во второй серии опытов замедляет функциональное выздоровление. Асимметрия гистограмм распределения мышечных волокон по диаметру в ипси- и контралатеральной мышцах, а значения их мод в сравнении с интактной мышцей наглядно свидетельствуют о сохраняющейся атрофии мышечных волокон в мышце оперированной конечности и их компенса- торной гипертрофии в контралатеральной. Как показывают результаты ранее выполненных исследований, в этой серии опытов замедлены регенерация аксонов и дифференцировка нервных волокон [7]. В данной работе объективизированы проявления более длительного бездействия мышцы оперированной конечности и функциональной перегрузки контралатеральной мышцы во второй группе опытов по сравнению с первой.
Фактор неравномерной нагрузки мышц оперированной и контралатеральной конечностей проявляется в третьей группе в подгруппах 1 и 2. Как показывают результаты наблюдений, животные слабо опираются на удлинённую конечность, но несколько лучше используют при стоянии и ходьбе предварительно укороченную конечность и опираются на неё после восстановления длины даже при дефиците функции седалищного нерва. Режим функциональной нагрузки неизбежно меняет численно-размерный состав мышечных волокон. В III серии опытов у разных индивидов наблюдается либо выраженное сужение гистограммы распределения мышечных волокон по диаметру на 2-3 класса либо уменьшение величины его моды. При ишемии передней большеберцовой мышцы [2], сопровождающейся повреждением митохондриального аппарата мышечных волокон, отмечается одновременное уменьшение вариативности их диаметров и снижение среднего диаметра. В наших опытах в подгруппах III-1 и III-2 наряду со снижением функциональной активности мышцы вероятны ритмически повторяющиеся периоды кратковременной ишемии, вызванные спазмированием бедренной артерии при "подкрутках" дистракционного аппарата; в подгруппе III-3 в начальном периоде опыта кровоток изменён "гофрированием" бедренной артерии при укорочении костного сегмента. Однако через 4 месяца после операции изменения размерных показателей мышечных волокон, объёмной плотности эндомизия, численной плотности мышечных волокон и микрососудов передней большеберцовой мышцы у животных III группы в сравнении с интактными не свидетельствуют об ишемии. Соотношения этих показателей заставляют предположить, что увеличение объёмной плотности эндомизия обусловлено главным образом уменьшением диаметров мышечных волокон, а не разрастанием соединительной ткани. При этом увеличение численной плотности мышечных волокон слишком велико, чтобы объяснить его только их топографическим перераспределением. Ещё большее увеличение численной плотности микрососудов указывает на усиление васкуляризации мышечных волокон.
ВЫВОДЫ
-
1. Через 4 месяца после первичного шва повреждённого седалищного нерва собак происходит эффективная компенсация денервационной атрофии передней большеберцовой мышцы с восстановлением численно-размерного состава мышечных волокон.
-
2. Сочетание повреждения нерва с остеотомией замедляет процесс восстановления морфофункциональных характеристик денервированной передней большеберцовой мышцы.
-
3. При регенерации седалищного нерва в условиях дозированного растяжения реиннервация передней большеберцовой мышцы сопровождается усилением васкуляризации, что предопределяет прогрессивную пластическую перестройку мышечной ткани и позволяет рекомендовать использование дистракционного остеосинтеза в лечении открытых переломов, сочетающихся с анатомическими перерывами и дефектами нервов.
