Цитогенетическая характеристика архара Ovis ammon ammon, снежного барана O. nivicola borealis и их гибридов

Автор: Багиров В.А., Кленовицкий П.М., Иолчиев Б.С., Зиновьева Н.А., Калашников В.В., Шило О.В., Солошенко В.А., Насибов Ш.Н., Кононов В.П., Колесников А.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетическое разнообразие, цитогенетика, вирусные болезни
Статья в выпуске: 6 т.47, 2012 года.
Бесплатный доступ
Резервом развития отечественного овцеводства может послужить гибридизация домашних овец с дикими представителями рода Ovis, позволяющая обогатить генофонд пород за счет внесения в него ценных генетических задатков, присущих диким сородичам. Гибридизация близкородственных видов также способна оказаться эффективным методом реконструкции и восстановления исчезающих представителей фауны. Нами была выполнена гибридизация снежного барана Ovis nivicola borealis и архара O. ammon ammon (соответственно отцовская и материнская формы), получено жизнеспособное плодовитое гибридное потомство и проведено сопоставление кариотипов архара, снежного барана, а также их гибрида. Поскольку родительские виды имеют различные диплоидные числа и неодинаковое число двуплечих аутосом, при нумерации хромосом и построении кариограммы мы использовали идеограмму, которая была основана на кариотипе козы, дополненном группой метацентрических хромосом. Результаты цитогенетического анализа в совокупности с данными о плодовитости гибридного потомства указывают на высокую степень близости сравниваемых видов.
Снежный баран ovis nivicola borealis, архар o. ammon ammon, гибридизация, кариотип
Короткий адрес: https://sciup.org/142133359
IDR: 142133359 | УДК: 636.32/.38:636.01:599.735.52:575
Cytogenetic characteristic of Ovis ammon ammon, O. nivicola borealis and their hybrids
By a hybridization of wild and domestic sheep, a gene fond of sheep breeds can be enriched with useful traits, and the rare and disappearing species reconstructed. In our experiments, the hybridization between Ovis nivicola borealis and O. ammon ammon resulted in a reproductive offspring. The parental and hybrid animals were compared as to their karyograms which were determined on the base of ideogram for goats with addition of metacentric chromosomes. The cytogenetic data and the reproductive state of the hybrid confirm that the taxa used in hybridization are closely related.
Текст научной статьи Цитогенетическая характеристика архара Ovis ammon ammon, снежного барана O. nivicola borealis и их гибридов
В России в силу суровых природно-климатических условий овцеводство всегда было важной отраслью сельского хозяйства, обеспечивающей потребности населения и легкой промышленности в продуктах питания и специфических видах животного сырья. В ряде случаев овцы — единственный вид животных, способных использовать имеющиеся при родные ресурсы.
Рис. 1. Динамика численности поголовья овец в миллиардах голов в мире (1, ось слева) и в России (2, ось справа) .
Большое разнообразие условий разведения и содержания привело к созданию приспособленных к ним многочисленных пород и породных групп овец с широким спектром генетически обусловленных морфологических и продуктивных качеств. Считается, что в настоящее время в мире существует около 850 пород овец (1).
По данным FAO (2), по сравнению с 1990 годом в 2010 году поголовье овец в мире снизилось на 10,7 %, составив при этом
1079 млн гол. Минимальная численность овец (1060 млн гол.) приходилась на 2000 год, к 2005 году их поголовье незначительно увеличилось и дос тигло примерно 1100 млн гол., однако в течение следующего пятилетия произошел очередной спад численности. Стабильный рост поголовья овец в анализируемый период наблюдался лишь в Азии и Африке (соответст- венно на 29 и 45 %), тогда как в остальных регионах оно значительно уменьшилось (в целом по Америке — на 25 %, по Европе и Океании — на 56 %).
В России в период с 1990 по 2000 год поголовье сократилось в 4,2 раза (с 55 до 13 млн гол.), однако в последнее десятилетие наблюдался его рост, и в 2010 году численность овец составила около 2 млн гол. (рис. 1). Несмотря на наметившиеся положительные перемены, отечественное овцеводство постоянно требует особого внимания. Резервом развития отрасли может послужить гибридизация с дикими представителями рода Ovis , позволяющая обогатить генофонд домашних овец за счет привнесения в него ценных генетических задатков, присущих диким сородичам (3). Гибридизация близкородственных видов способна также оказаться эффективным методом реконструкции и восстановления исчезающих видов (4-10).
Ареал диких овец охватывает обширную территорию от островов Средиземного моря и областей Передней Азии до Камчатки на Евразийском континенте и на западе Северной Америки. Представители рода Ovis хорошо приспособлены к жизни в различных географических зонах (от степей до горных хребтов), верхняя граница их обитания проходит на высоте 5000 м над уровнем моря (11).
В систематике диких баранов до настоящего времени нет единого взгляда на структуру рода Ovis . Наиболее обосновано выделение семи видов диких овец — уриал ( Ovis vignei ), муфлон европейский ( O . musimon ), муфлон азиатский ( O . orientalis ), архар, или горный баран ( O . ammon ), снежный баран ( O . nivicola ), толсторог ( O . canadensis ), тонкорог ( O . dalli ). Результаты цитогенетических (12-15) и молекулярно-генетических (16-19) исследований однозначно свидетельствуют о том, что предком домашней овцы был азиатский муфлон ( O . orientalis ). Участие в доместикации уриала и архара противоречит этим данным. В отношении муфлона европейского ( O . musimon ) высказано предположение, что он является потомком одичавших овец (1). Хотя в овцеводстве имеется опыт получения гибридов домашней овцы с представителями разных видов диких баранов (5-7, 20-23), данные о гибридизации видов диких баранов между собой в литературе отсутствуют.
Цель настоящей работы заключалась в получении гибридов архара ( O . ammon ammon ) со снежным бараном ( Ovis nivicola borealis ) и их кариотипическом изучении.
Методика . В экспериментах по гибридизации в качестве отцовской формы был взят снежный баран ( Ovis nivicola borealis ), материнской — матки архара ( O . ammon ammon ). При цитогенетическом анализе исходных видов и гибрида материалом для исследования служила 72-часовая культура периферических лимфоцитов, стимулированных конканавали-ном А (ООО «ПанЭко», Россия) в дозе 10 мкг/мл. Культивирование, получение и обработку препаратов хромосом проводили по общепринятой методике с внесенными нами модификациями (24).
Результаты исследования документировали с помощью цифровой видеокамеры КС-583С («Digital», Тайвань) и пакета программ, совместимых с Windows. Обработку изображений и кариотипирование осуществляли по описанной ранее методике (25).
Резуёътаты . Полагают, что исходный кариотип полорогих ( Bovidae ) содержал 60 хромосом, включая 29 пар одноплечих аутосом. Расхождение ветвей, давших начало современным представителям этого семейства, произошло 15-20 млн лет назад. Из современных полорогих кариологически наиболее близки к предковой форме представители рода Capra , обладающие хромосомной мономорфностью (26, 27).
Род Ovis характеризуется четко выраженным хромосомным полиморфизмом. Входящие в него виды образуют четыре хромосомных класса. Принято считать, что хромосомные наборы в пределах рода Ovis , как и у всех полорогих, эволюционировали посредством центрических слияний. Первый, самый древний хромосомный класс с 2 n = 58 включает один вид — O . vignei . Кариотип уриала содержит только одну пару метацентрических хромосом. Ко второму классу относится архар, или горный баран ( O . ammon , 2 n = 56), в кариотипе которого имеются две пары метацентрических хромосом. В третий класс входит муфлон европейский ( O . mu-simon ), муфлон азиатский ( O . orientalis ), толсторог ( O . canadensis ), тонкорог ( O . dalli ) и домашняя овца ( O . aries ). Кариотипы указанных видов идентичны, они содержат 54 хромосомы, в том числе три пары метацентриков. Эволюционно более молодой четвертый класс с 2 n = 52, включающий единственный вид — снежного барана ( O . nivicola ). В кариотипе снежного барана представлены четыре пары метацентрических хромосом.
К ранним попыткам выявить соответствие между хромосомами козы и разных видов рода Ovis относятся работы T.D. Bunch с соавт. (28, 29). Первоначально T.D. Bunch на основании результатов дифференциального G-окрашивания считал, что метацентрические хромосомы у всех видов овец возникли в результате четырех робертсоновских транслокаций, соответствующих центрическим слияниям козьих хромосом 1/5-й, 3/10-й, 4/9-й и 11/17-й (29). Позднее, исследуя кариотип у барана Северцова ( O . ammon severtzovi ), T.D. Bunch с соавт. (30) показали, что это подвид архара. Авторы отметили, что метацентрики у архара эквиваленты 1-й, 3-й, 5-й и 11-й хромосомам предкового кариотипа Ovis .
В результате применения дифференциального окрашивания с высоким разрешением удалось уточнить, какие эволюционные преобразования предкового кариотипа происходили в процессе дивергенции рода Ovis . Первая пара двуплечих хромосом у всех современных овец образовалась в результате слияния акроцентрических хромосом, соответствующих 1-й и 3-й парам хромосом предковой формы. Впервые она появилась у уриала ( О . vignei , 2 n = 58) и присутствует у всех истинных овец с хромосомными числами 56, 54 и 52. Вторая пара метацентрических хромосом возникла при объединении 2-й и 8-й хромосом, которое произошло у предка архара и сохраняется у горных баранов с диплоидными числами 56, 54 и 52. Третья пара двуплечих хромосом сформировалась вследствие центрического слияния 5-й и 11-й хромосом, что привело к появлению видов с кариотипом, содержащим 54 хромосомы. Этот метацентрик присутствует и у снежного барана, кариотип которого содержит 52 хромосомы. Уменьшение диплоидного числа при образовании кариотипа O . nivicola произошло в результате слияния 9-й и 19-й хромосом (31).
Как показал анализ специальной литературы, цитогенетические исследования диких овец, выполненные на уровне подвидов, далеко не полны. В частности, из семи подвидов азиатского снежного барана (32, 33) кариотип описан только у O . nivicola nivicola (14, 34).
При сопоставлении кариотипов архара и снежного барана, а также их гибрида мы столкнулись с методической проблемой: поскольку оба исходных вида имеют различные диплоидные числа и неодинаковое число двуплечих аутосом, построение и сравнительное описание кариотипов с применением стандартных принципов кариотипирования было затруднено. Поэтому при нумерации хромосом и построении кариограммы мы использовали идеограмму, основанную на кариотипе козы (базовая часть), которую дополняли группой метацентрических хромосом. В зависимости
A
Б

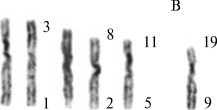
Кариотипы самцов архара ( Ovis
Рис. 2.
ammon ammon) (A), снежного барана (O. nivicola borealis) (Б) и их гибрида (В). Дифференциальное G-окрашивание, увеличение *100.
хромосом равнялось 52,
как и у
O .
от видовой принадлежности исследуемого животного число хромосомных элементов и их размещение изменялись.
В кариотипе архара (рис. 2, A) хромосомы 3-6-й пар соответствуют 4-7-й парам базового кариотипа, с 7-й по 27-ю пару — 9-29-й парам хромосом козы, метацентрические хромосомы — 1/3-й и 2/8-й хромосомам базового кариотипа. Диплоидное число хромосом не отличалось от видовой нормы (2n = 56). В кариотипе снежного барана O. nivicola borealis (см. рис. 2, Б) диплоидное число ветствовали 4-й, 6-й, 7-й, 10-й, с
nivicola nivicola (35), акроцентрики соот-12-й по 18-й и далее с 20-й по 29-ю хро- мосому базового кариотипа, метацентрики — 1/3-й, 2/8-й, 5/11-й и 9/19-й хромосомам базового кариотипа. У гибрида снежного барана с архаром (см. рис. 2, В) хромосомное число равнялось теоретически ожидаемому (2n = 54). Метацентрические хромосомы были представлены двумя парами гомологов, соответствующими хромосомам 1/3-й и 2/8-й исходного кариотипа, унаследованным как от отца, так и от матери, и двумя непарными метацентрика-ми (5/11-я и 9/19-я), полученными от снежного барана. Парные акроцентрики, унаследованные от обоих родителей, соответствовали базовым хромосомам 4-й, 6-й, 7-й, 10-й, 12-18-й и 29-29-й, непарные акроцентрики, полученные от архара, — базовым хромосомам 5-й, 9-й, 11-й и 19-й. У животных исходных видов и гибридов в базовой части идеограммы отсутствовали 1-я, 2-я, 3-я и 8-я хромосомы, соответствующие 1-й и 2-й метацентрическим.
У всех исследованных животных фенотипический пол совпадал с определенным по составу половых хромосом. У архара, снежного барана и их гибридов X-хромосома представляла собой крупный акроцетрик, тогда как Y-хромосома была мелким метацентриком.
Как показали исследования, гибридные самцы первого поколения F 1 были плодовитыми, что редко происходит при отдаленной гибридизации. Аналогичные результаты получены при гибридизации домашних овец с камчатским снежным бараном O . nivicola nivicola (35). В совокупности с данными цитогенетического анализа это указывает на высокую степень близости сравниваемых видов.
Таким образом, получено жизнеспособное плодовитое гибридное потомство и проведено сопоставление кариотипов архара, снежного барана и их гибрида. Результаты цитогенетического анализа и данные о плодовитости гибридного потомства указывают на высокую степень близости сравниваемых исходных видов. Плодовитость созданных гибридных самцов — важное качество, позволяющее тиражировать ценных особей в широких масштабах, что составляет основу для формирования высокопродуктивных популяций овец.
