Цитогенетический анализ гибридов F1, полученных от скрещивания линий Triticum aestivum L. с видами Aegilops L

Автор: Намазова Л.Х., Алиева А.Д.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 9 т.9, 2023 года.
Бесплатный доступ
В результате определения степени завязываемости семян в гибридных комбинациях, созданных между линиями Triticum aestivum L. и видами рода Aegilops L., а также цитогенетического анализа гибридов F1 было установлено, что обладание родительских форм одинаковыми субгеномами при гибридизации не может гарантировать относительно высокий процент завязываемости, а при мейозе высокий уровень конъюгации хромосом. В то же время, не обнаружено никаких существенных коррелятивных отношений между уровнем конъюгации хромосом и фертильностью у исследуемых межродовых гибридов F1.
Aegilops, triticum aestivum, межродовая гибридизация, гибридная комбинация, завязывание семян, гибриды f1
Короткий адрес: https://sciup.org/14128699
IDR: 14128699 | УДК: 575.222.73: | DOI: 10.33619/2414-2948/94/04
Cytogenetic analysis of F1 hybrids derived from crossings of Triticum aestivum L. lines with Aegilops L. species
As a result of the studies seed settings of hybrid combinations obtained from the crossings between Triticum aestivum L. lines and Aegilops L. species, also the cytogenetic analysis of the F1 hybrids belonging to these combinations have been established that possession joint subgenomes of parental forms is not guarantee a relatively high percentage of fertility at crosses, and a high level of chromosome pairing in meiosis. At the same time, no significant correlations were found between the level of chromosome conjugation and fertility in the studied intergeneric F1 hybrids.
Текст научной статьи Цитогенетический анализ гибридов F1, полученных от скрещивания линий Triticum aestivum L. с видами Aegilops L
Бюллетень науки и практики / Bulletin of Science and Practice
UDC 575.222.73: 633.11
Интрогрессия полезных генов других видов или пород в геном пшеницы приводит к изменению ее генотипа, что создает благоприятную основу для выбора подходящих форм с точки зрения селекции. Известно, что дикий предок пшеницы, Aegilops L., обладает такими важными агрономическими характеристиками, как количественное и качественное улучшение белка в зерне этого рода [1, 2], устойчивость к грибковым и вирусным заболеваниям [1–7], вредителям и нематодам [8–12], а также засухоустойчивости и солевыносливости [13–16]. Однако лишь небольшое количество его хромосом может конъюгировать с хромосомами пшеницы и передавать им свои полезные гены с помощью обычных методов селекции.
Проведя систематические перекрестные связи между диплоидным типом Ae. umbellulata (UU, 2n=2x=14) и тетраплоидным (AABB, 2n=4x=28) сортом Langdon Triticum durum Desf., выявили 3 типа гибридной несовместимости и стали свидетелями демонстрации, что амфиплоидные растения F 1 (2n=3x=21) были либо сильно недоразвитыми, либо травоядными [5].
Asghar M., Rao A., Farooq S. путем гибридизации определили частоту формирования хиазмы (ЧФХ), образованной конъюгацией между D-геномом Triticum aestivum L. и хромосомами D-генома всех трех видов эгилопсов в полученных гибридах F 1 , обнаружили, что гомология между типом Ae. tauschii и геномами D Triticum aestivum L. выше (на каждую образовалось 11,9 хиазмов), чем у видов Ae. cylindrica и Ae. crassa [4].
Возможность переноса инородных хромосом в мягкую пшеницу даже на самых низких уровнях ЧФХ показывает, что все три вида эгилопсов важны с точки зрения улучшения генофонда Triticum aestivum L. Так как Triticum L. и Aegilops L., которые считаются вторичным генофондом пшеницы, принадлежат к неконгруэнтным скрещиваниям, естественно, что уровень конъюгации между их хромосомами низкий, а различия между уровнями плоидности родительских форм также приводят к образованию слабых и бесплодных и, в лучшем случае, полустерильных гибридов во многих случаях в результате оплодотворения несбалансированных гибридов. Однако все препятствия на пути несовместимости генома и цитоплазмы во время гибридизации между этими видами не только не демотивируют тритикологов, но и побуждают их искать новые способы преодоления несовместимости между родами [17–20].
Основная цель исследования заключалась в проведении межполовой гибридизации для переноса полезных генов рода Aegilops L., которые контролируют важные агрономические признаки, перечисленные выше, путем гибридизации в линии мягкой пшеницы и изучение способности инородных хромосом конъюгировать во время мейоза.
Материалы и методы
В качестве материала исследования были использованы стабильные линии Triticum aestivum L. из коллекции отделения молекулярной цитогенетики 171ACS и 172ACS для рода Aegilops L. Многие виды были собраны из разных регионов Азербайджана сотрудниками отделения в ходе местных и международных экспедиций. Гибридологический метод использовался в основном для получения межродовых гибридов между пшеницей и эгилопсом. Колосья растений-реципиентов опыляли пыльцой опыленных и донорных растений по общепринятому правилу [21].
Полученные F 1 были тщательно проанализированы [22–25]. Ранней весной колосья гибридных растений фиксировались на стадии трубчатого образования. В качестве фиксатора использовали раствор Карнуа, состоящий из смеси спирта и уксусной кислоты в соотношении 3:1. Фиксированный материал переводили в 80° спиртовой раствор и хранили в холодильнике. Процесс исследования мейоза в материнских пыльцевых клетках (МПК) осуществлялся нижеследующим образом: пыльца снималась с цветков колоса и помещалась в тигли с раствором ацетокармина для окрашивания. Материал хранился в холодильнике до окрашивания при условии нагрева несколько раз в день (Рисунок). Результаты мейотического процесса были обработаны математически и статистически [25].
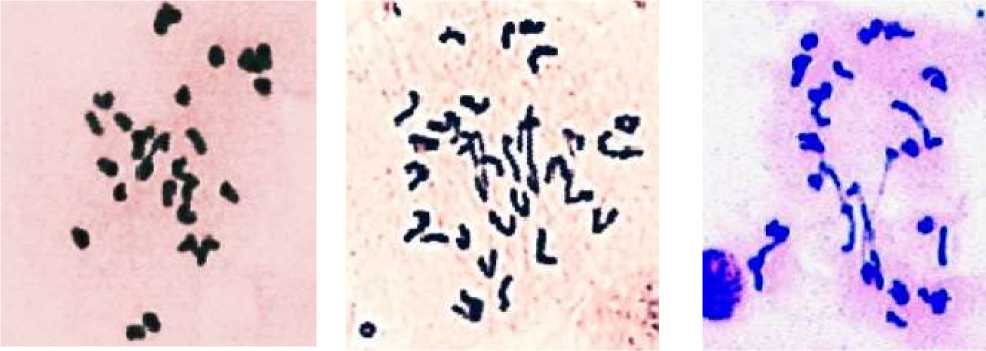
a b c
Рисунок. Метафаза во время мейоза у гибридов F 1 комбинаций: a — 171ACS × Ae. umbellulata , b — 171ACS × Ae. biuncialis и c — 171ACS × Ae. recta
Результаты и обсуждение
Результаты гибридизации линий Triticum aestivum L. 171ACS и 172ACS, принадлежащими к трем секциям Cylindropyrum , Vertebrata и Aegilops рода Aegilops L. подрода Aegilops , представлены в Таблице 1.
Таблица 1
ЗНАЧЕНИЯ ЗАВЯЗЫВАЕМОСТИ И ФЕРТИЛЬНОСТИ РАСТЕНИЙ F1 В ГИБРИДНЫХ КОМБИНАЦИЯХ МЕЖДУ ЛИНИЯМИ Triticum aestivum L. И ВИДАМИ Aegilops L.
|
Комбинации |
Завязываемость, % |
Фертильность, F 1 % |
Высота, см |
|
|
Секция Cylindropyrum |
||||
|
171ACS × Ae. cylindrica (Гобустан) |
2,86 |
стерильны |
64 |
|
|
172ACS × Ae. cylindrica (Oрдубад-Тиви) |
7,14 |
стерильны |
75 |
|
|
Секция Vertebrata |
||||
|
172ACS × Ae. crassa (Азербайджан) k-2422 |
1,35 |
— |
||
|
Ae. crassa (Азербайджан) k-2422 × 172ACS |
50,00 |
стерильны |
64 |
|
|
171ACS × Ae. trivialis (Афганистан) k-1003 |
6,58 |
0,25 |
89 |
|
|
171ACS × Ae. trivialis (Афганистан) k-1012 |
6,48 |
0,14 |
88 |
|
|
172ACS × Ae. vavilovii |
1,32 |
— |
||
|
Секция Aegilops |
||||
|
171ACS × Ae. umbellulata (Гирдиманчай ) |
14,52 |
0,06 |
121 |
|
|
171ACS × Ae. peregrina (Израиль) k-539384 |
1,25 |
стерильны |
60 |
|
|
171ACS × Ae. kotschyi (Азербайджан) k-91 |
32,90 |
0,03 |
75 |
|
|
172ACS × Ae. kotschyi (Азербайджан) k-91 |
8,33 |
0,37 |
84 |
|
|
171ACS × Ae. geniculata (Испания) k-2113 |
10,00 |
стерильны |
65 |
|
|
171ACS × Ae. triuncialis (Гирдиманчай) |
2,70 |
стерильны |
66 |
|
|
(к) Ф Тип лицензии CC: Attribution 4.0 International (CC BY 4.0) |
32 |
|||
Бюллетень науки и практики / Bulletin of Science and Practice Т. 9. №9. 2023
|
Комбинации |
Завязываемость, % |
Фертильность, F 1 % |
Высота, см |
|
172ACS × Ae. triuncialis (Гирдиманчай) |
9,46 |
стерильны |
82 |
|
171ACS × Ae. biuncialis (Гобустан) |
5,17 |
0,63 |
100 |
|
171ACS × Ae. columnaris (Турция) k-3472 |
26,56 |
стерильны |
84 |
|
171ACS × Ae. neglecta (Гирдиманчай) |
31,08 |
0,06 |
75 |
|
171ACS × Ae. neglecta |
1,56 |
стерильны |
55 |
|
172ACS × Ae. neglecta (Гирдиманчай) |
37,14 |
0,07 |
83 |
|
171ACS × Ae. recta |
15,91 |
стерильны |
76 |
Как видно из Таблицы 1, в гибридных комбинациях между линиями Triticum aestivum L. и видами эгилопса прививка зерна в лучшем случае составляла 50,00% ( Ae. crassa × 172ACS) и 1,25% (171ACS × Ae. peregrina ) в худшем случае, варьируя между этими двумя верхними и нижними пределами. Полученные гибридные зерна обычно были мелкими и рыхлыми.
Для определения характера конъюгации инородных хромосом у растений F 1 , полученных в результате прорастания этих зерен, был изучен процесс мейоза в каждом из них и полученные результаты представлены в Таблице 2.
Как видно из Таблицы 2, в гибридной комбинации между линией 171ACS Triticum aestivum L. и Ae. cylindrica , собранными в Гобустане, урожайность составила 2,86%, из обоих созревших зерен выросли пентаплоидные растения F 1 . Растения были стерильны, в среднем в высоту 64 см. Так, что из 392 цветков с колосом не удалось получить ни одного F 2 . При исследовании мейоза было замечено, что уровень конъюгации хромосом был очень низким. Хотя оба родителя, участвовавшие в приобретении гибрида, имели гомологичный субгеном формы D, количество закрытых и открытых бивалентов для каждого МПК составляло 0,58 и 2,32, количество унивалентов составляло 29,22, а ЧФХ составляла 3,48.
В гибридной комбинации между линией мягкой пшеницы 172ACS и Ae. cylindrica , собранными в деревне Ордубад-Тиви, урожай составил 7,14%, 2 из 4 семян проросли, и только один из этих проростков дал начало пентаплоидному растению F 1 . Растения были стерильны, в среднем в высоту 75 см. Таким образом, не удалось получить F 2 из 810 колосков. Однако кроме того, при исследовании мейоза уровень конъюгации хромосом оказался относительно высоким по сравнению с предыдущим одноименным гибридом. Так, что количество закрытых и открытых бивалентов для каждого МПК составило соответственно 5,22 и 1,80, количество унивалентов — 20,22, количество тривалентов — 0,24, а количество ЧФХ — 12,81.
Как видно, несмотря на то, что оба гибрида F 1 получили помеси между линией мягкой пшеницы и Ae. cylindrica , результаты мейотического анализа немного различались и уровень конъюгации хромосом у второго гибрида был примерно в 4 раза выше, чем у первого. С большой вероятностью это можно объяснить использованием в гибридизации двух разных экотипов вида Ae. cylindrica (Гобустан и Ордубад). Клетки были на 100% стерильными, а яйцеклетки были частично фертильными.
В гибридной комбинации между линией 172ACS и Ae. crassa азербайджанского происхождения урожай зерна составил 1,35%, и, несмотря на прорастание одного полученного хрупкого зерна, этот проросток после пересадки в поле был уничтожен. При участии этих родителей в обратной комбинации ( Ae. crassa × 172ACS) урожай зерна был намного выше, т. е. 50%, по сравнению с простой комбинацией, и каждое из 3 полученных гибридных зерен дало начало пентаплоидным растениям F 1 .
Бюллетень науки и практики / Bulletin of Science and Practice Т. 9. №9. 2023
Таблица 2
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ПОВЕДЕНИЯ ХРОМОСОМ В МЕЙОЗЕ ГИБРИДОВ F 1
Cylindropyrum
171ACS x Ae. cylindrica (Гобустан )
119 2,90±0,65 0,58±0,42 2,32±0,26 29,20±1,29 3,48±0,1835
172ACS × Ae. cylindrica (Oрдубад-Тиви)
112 7,03±0,38 5,22±0,25 1,80±0,41 20,22±0,79 0,24±0,10 12,81±0,5235
Vertebrata
Ae. crassa (Азербайджан) k-2422 × 172ACS
118 3,52±0,52 0,09±0,12 3,42±1,50 27,18±1,33 0,26±0,19 4,13±0,4635
171ACS × Ae. trivialis (Афганистан) k-1003
133 5,08±0,37 1,94±0,41 3,14±2,36 31,50±0,55 0,11±0,08 6,31±0,4242
171ACS × Ae. trivialis (Афганистан ) k-1012
101 4,87±0,45 0,86±0,22 4,00±3,31 31,60±0,88 0,22±0,12 6,17±0,4942
Aegilops
171ACS × Ae. umbellulata (Гирдиманчай)
135 2,93±0,25 0,31±0,13 2,62±0,31 21,15±0,55 0,33±0,12 3,90±0,3028
171ACS × Ae. peregrina (Израиль) k-539384
135 3,91±0,40 0,56±0,26 3,36±2,22 26,99±0,94 0,06±0,08 4,61±0,4335
171ACS × Ae. kotschyi ( Азербайджан ) k-91
109 4,48±0,18 0,46±0,12 4,02±0,16 26,04±0,39 7,74±0,2035
172ACS × Ae. kotschyi ( Азербайджан ) k-91
144 3,57±0,29 0,51±0,36 3,06±0,40 26,99±0,84 0,15±0,11 4,76±0,7435
гексав
0,05±0,11
171ACS × Ae. geniculata (Испания) k-2113
119 4,28±0,22 0,86±0,23 3,43±0,37 25,90±0,82 0,18±0,14 5,52±0,4035
171ACS × Ae. triuncialis (Гирдиманчай )
108 2,10±0,27 0,18±0,12 1,92±2,82 30,62±0,70 0,06±0,09 2,40±0,4635
172ACS × Ae. triuncialis (Гирдиманчай )
80 3,30±0,26 — 3,30±0,26 28,10±0,71 0,10±0,14 3,51±0,1335
171ACS × Ae. biuncialis (Гобустан)
186 3,01±0,46 0,20±0,09 2,81±0,48 26,97±1,00 0,67±0,29 4,55±0,6035
171ACS × Ae. columnaris (Турция) k-3472
102 4,90±0,22 1,91±0,35 3,00±0,39 24,67±0,92 0,18±0,12 7,19±0,4335
171ACS × Ae. neglecta (Гирдиманчай )
111 4,11±0,36 0,32±0,11 3,78±1,87 26,40±0,86 0,13±0,09 4,81±0,4735
171ACS × Ae. neglecta
124 2,51±0,25 0,13±0,12 2,38±2,59 29,69±0,72 0,10±0,12 2,84±0,4435
172ACS × Ae. neglecta (Гирдиманчай)
123 5,41±0,50 1,56±0,31 3,98±2,29 23,93±1,36 0,08±0,08 7,26±0,4435
171ACS × Ae. recta
130 6,34±0,24 0,34±0,19 6,00±0,34 29,32±0,48 6,68±0,2742
Средняя высота растений составила 64 см, все 328 колосков были стерильными. При исследовании мейоза было замечено, что уровень конъюгации хромосом очень низкий. Таким образом, количество закрытых и открытых бивалентов для каждого МПК составило 0,09 и 3,42 соответственно, количество унивалентов — 27,18, количество тривалентов — 0,26, а ЧФХ — 4,13.
Наибольшее количество уникальных аллелей было зарегистрировано Нагави и др. в образцах Ae. crassa , что указывает на то, что этот вид является отличным потенциальным источником новых генов для улучшения Triticum aestivum L. [13].
В гибридной комбинации между линией 171ACS и Ae. trivialis афганского происхождения урожай составил 6,58%, при этом каждое из 5 зерен прорастало и давало нормальные основы гексаплоидных растений F 1 . Средняя высота растений составила 89 см, фертильность 0,25%. Таким образом, было получено 4 зерна F 2 по 1622 цветков колоса. При изучении мейотического процесса у растений F 1 было определено, что количество закрытых и открытых бивалентов для каждого МПК составляло 1,94 и 3,14, количество унивалентов — 31,50, количество тривалентов — 0,11, количество ЧФХ равнялось 6,31.
В гибридной комбинации между линией 171ACS и другим образцом Ae. trivialis (k-1012) афганского происхождения урожай зерна составил 6,48%, при этом 5 из 7 семян проросли, из которых только 4 дали основу растения F 1 . Средний рост растений составил 88 см, фертильность — 0,14%, всего из 2 зерен F 2 было получено 1402 колоска. При изучении мейотического процесса у гексаплоидных растений F 1 количество закрытых и открытых бивалентов для каждого МПК составило 0,86 и 4,00, количество унивалентов — 31,60, количество тривалентов — 0,22, количество ЧФХ — 6,17.
В гибридной комбинации между линией 172ACS и Ae. vavilovii урожайность составила 1,32%, но хотя прорастал единственный полученный гибрид, он был уничтожен до конца вегетационного периода. Интересен тот факт, что виды Cylindropyrum и Vertebrata , участвующие в гибридизации, имели общий, т. е. гомологичный субгеном D с мягкой пшеницей, и на самом деле этот факт должен был положительно влиять на конъюгацию хромосом и фертильность гибридов F 1 между ними.
Однако в нашей практике оба экземпляра Ae. cylindrica с Triticum aestivum L., а также гибриды F 1 между Ae. crassa оказались стерильными, а растения F 1 между другим образцом Ae. crassa с Triticum aestivum L. и Ae. vavilovii были уничтожены до конца вегетационного периода. С этой точки зрения полустерильные гибриды F 1 между Triticum aestivum L. и двумя образцами Ae. trivialis афганского происхождения различались незначительно, и их фертильность составила 0,25 и 0,14% соответственно.
Что касается уровня конъюгации хромосом, за исключением гибрида F 1 , полученного только из комбинации 172ACS × Ae. cylindrica , количество хиазм на МПК у других гибридов первого поколения варьировало от 4 до 6, что намного ниже теоретически ожидаемого. Однако парадоксально, что количество хиазмов на МПК в гибриде вышеупомянутой комбинации 172ACS × Ae. cylindrica близко к теоретическому ожиданию, то есть около 13, но в обоих гибридах, полученных из комбинации 171ACS × Ae. trivialis , каждая МПК полностью стерильна, и несмотря на то, что число хиазмов было 6, они были несколько фертильными. Это показывает, что фертильность гибридных растений зависит не только от уровня конъюгации инородных хромосом.
Наличие гомологичных геномов в родительских формах не всегда обеспечивает фертильность гибридов F1. Так, что и они сообщили о низкой фертильности (0,1–6,5%) у гибридов Triticum aestivum L. и Ae. trivialis и Ae. vavilovii и F1, гексаплоидных видов эгилопса, малой и плохой всхожести эндосперма зерна, а приобретение гаплоидного растения (2n = 21) от комбинации Ae. vavilovii × T. aestivum объясняли склонностью к апомиксису или партенокарпию [17].
Участие линий мягкой пшеницы в гибридизации с видами, принадлежащими к секции Aegilops , и изучение мейотического процесса у полученных гибридов F 1 представляло особый интерес в связи с тем, что родительские формы не имели общего субгенома.
В гибридной комбинации между линией 171ACS и Ae. umbellulata , собранными в Гирдманчае, урожай зерна составил 14,52%, при этом только 1 из 9 семян проросло, дав начало нормальному тетраплоидному растению F 1 . Средний рост растения составлял 121 см, фертильность 0,06%. Таким образом, из 1554 колосков получено одно зерно. Во время исследования мейоза наблюдалась очень низкая хромосомная конъюгация, в среднем для каждого МПК учитывалось 0,31 закрытый, 0,62 открытый бивалент, 21,15 унивалент, 0,33 тривалент и 3,90 хиазм. В гибриде F 1 комбинации 171ACS × Ae. umbellulata образование закрытых и открытых бивалентов, а также тривалентных конфигураций, несомненно, стало возможным благодаря конъюгации между гомеологическими хромосомами пшеницы и эгилопса.
Дж. Дворжаком и другими было обнаружено, что гомологичная конъюгация происходит между хромосомой 1U Ae. umbellulata и хромосомами 1A, 1B и 1D Triticum aestivum L., и что хромосома 1U, которая контролирует 7 субъединиц глиадина, ближе к 1B и 1D, чем 1A.
В гибридной комбинации между линией 171ACS и Ae. peregrina израильского происхождения урожай составил 1,25%, и пентаплоид F 1 смог дать начало растению путем прорастания из единственного полученного семени. Высота этого растения была небольшой — 60 см. К сожалению, он оказался совершенно стерильным (бесплодным), и нам не удалось достать даже одно зерно из 584 колосков. Во время исследования мейоза количество закрытых и открытых бивалентов для каждого МПК составляло 0,56 и 3,36 соответственно, количество унивалентов — 26,99, количество тривалентов — 0,06, а ЧФХ — 4,61.
В гибридной комбинации между линией 171ACS и Ae. kotschyi азербайджанского происхождения урожай зерна составил 32,90%, 23 из 25 семян проросли, и только 19 из них смогли завершить вегетационный период и дать начало пентаплоидным растениям F 1 . Средний рост этих растений составлял 75 см, а их фертильность была очень низкой и составляла 0,03%. Таким образом, из 3232 колосков был получен только один F 2 . При изучении мейоза у растений-гибридов F 1 было обнаружено, что количество закрытых и открытых бивалентов для каждого МПК составляло 0,46 и 4,02 соответственно, количество унивалентов составляло 26,04, а количество ЧФХ составляло 7,74, что указывает на то, что уровень конъюгации низкий.
В гибридной комбинации другой линии Triticum aestivum L. 172ACS и Ae. kotschyi азербайджанского происхождения урожайность составила 8,33%, 1 из полученных 4 семян проросло, и смогло пустить ростки для пентаплоидного растения F 1 . Средний рост этого растения составлял 84 см, а его фертильность была очень низкой — 0,37%. Так, что из 2142 цветков колоса были получены только 8 зерен F 2 . При исследовании мейоза у пентаплоидного гибрида F 1 количество закрытых бивалентов для каждого МПК составило 0,51, количество открытых бивалентов — 3,06, количество унивалентов — 26,99, количество квадривалентов и гексавалентов — 0,15 и 0,05 соответственно, и ЧФХ — 4,76.
В гибридной комбинации между линией 171ACS и Ae. geniculata израильского происхождения (MoMoUU, 2n=4x=28) урожай составил 10%, 6 из 7 семян проросли, и эти проростки дали основу 5 пентаплоидным растениям F1 (ABDMoU, 2n = 5x = 35). Высота растений 65 см, а сами стерилизованы — из 508 колосков не получили ни одного зерна. При исследовании мейоза у гибридов F1 количество закрытых и открытых бивалентов для каждого МПК составило 0,56 и 3,36 соответственно, количество унивалентов — 26,99, количество тривалентов — 0,06, количество ЧФХ — 4,61. В проведенной практике Aшгара M. и др., у гибридов F1, полученных от скрещивания сортов Triticum aestivum L. Chinese Spring и Ae. geniculata наблюдались 1 закрытый и 4 открытых бивалента, а также 2 тривалента, было определено, что ЧФХ составляет 4 для каждой клетки, что полностью аналогично нашим результатам мейотического анализа.
В гибридной комбинации между линией 171ACS и Ae. triuncialis , собранными в Гирдманчае, урожай зерна составил 2,70%, при этом 1 из 2 прорастающих семян давал начало нормальному F 1 растению. Высота растения 66 см, а само растение стерилизовано. Так, из 652 колосовых цветов не удалось получить ни одного зерна. При изучении мейоза у пентаплоидного растения F 1 было обнаружено, что количество закрытых и открытых бивалентов для каждого МПК составляло 0,18 и 1,92 соответственно, количество унивалентов — 30,62, количество тривалентов — 0,06, а количество ЧФХ равнялось 2,40.
В гибридной комбинации между другой линией Triticum aestivum L. 172ACS и Ae. triuncialis , собранной из Гирдманчая, урожайность (всхожесть) составила 9,46%, 7 из 7 семян проросли, и из этих проростков развились только 5 пентаплоидов F 1 . Растения были стерильными, высота составляла 82 см. Так, что из 190 колосков не было получено ни одного зерна. При изучении мейоза у растений F 1 не были зарегистрировано закрытых бивалентов, а открытые биваленты, униваленты, триваленты и ЧФХ оказались равными 3,30, 28,10, 0,10 и 3,51 для каждого МПК соответственно.
В гибридной комбинации между линией 171ACS и Ae. biuncialis , собранными в Гобустане, урожайность составила 5,17%, и 3 из 3 полученных семян прорастая, давали основу пентаплоидным растениям F 1 . Высота растений составляла 100 см, фертильность 0,63%. Таким образом, из 2062 цветков колоса было получено 13 зерен F 2 . При изучении мейоза у растений F 1 общее количество бивалентов для каждого МПК составило в среднем 3,01 (закрытый — 0,20 и открытый — 2,81), количество унивалентов — 26,97, количество тривалентов — 0,67, ЧФХ — 4,55.
В гибридной комбинации между линией 171ACS и Ae. columnaris турецкого происхождения урожайность зерна составила 26,56%, все 17 семян проросли, и из этих проростков было получено 15 пентаплоидных растений F 1 . Растения были ростом 84 см, сами же были стерильными — не было получено ни одного зерна из 3004 цветков колоса. При изучении мейоза у растений F 1 было определено, что количество закрытых и открытых бивалентов для каждого МПК составило 1,91 и 3,00 соответственно, количество унивалентов — 24,67, количество тривалентов — 0,18, а количество ЧФХ — 7,19.
В гибридной комбинации между линией 171ACS и Ae. neglecta , собранной с Гирдманчае, урожайность составила 31,08%, при этом 20 из 23 семян проросли, из которых только 18 дали начало пентаплоидным растениям F 1 . Средняя высота растений 75 см, фертильность 0,06%. Так, что из 1632 цветков колоса было получено только одно зерно. При изучении мейоза у гибридов F 1 количество закрытых и открытых бивалентов для каждого МПК составило 0,32 и 3,78 соответственно, количество унивалентов — 26,40, количество тривалентов — 0,13, количество ЧФХ — 4,81.
В гибридной комбинации между линией 171ACS и другим образцом Ae. neglecta урожайность составила 1,56%, прорастая от одного полученного гибрида и давая начало пентаплоидному растению F 1 .
Высота растения 83 см, само стерильное. Таким образом, из 242 цветков колоска не было получено ни одного зерна. При изучении мейоза у растений F 1 было определено, что количество закрытых и открытых бивалентов для каждого МПК составило 0,13 и 2,38 соответственно, количество унивалентов — 29,69, количество тривалентов — 0,10, а количество ЧФХ — 2,84.
В гибридной комбинации между другой линией мягкой пшеницы 172ACS и Ae. neglecta , собранными в Гирдманчае, урожайность составила 37,14%, 18 из 26 семян проросли, и только 14 из проросших дали начало пентаплоидным растениям F 1 . Средняя высота растений 55 см, фертильность 0,07%. Таким образом, из 1370 цветков колоска удалось получить только одно зерно. При изучении мейоза у гибридов F 1 количество закрытых и открытых бивалентов для каждого МПК составило 1,56 и 3,98 соответственно, количество унивалентов — 23,93, количество тривалентов — 0,08, количество ЧФХ — 7,26.
В гибридной комбинации между линией 171ACS и Ae. recta урожайность зерна составила 15,91%, единственное полученное проросшее слабое зерно, прорастая дало рост нормальному гексаплоидному растению F 1 . Высота растения 76 см, само было стерильным, т. е. получить зерно с 546 цветков колосков не удалось. При изучении мейоза у растений F 1 для каждого МПК было зарегистрировано в среднем 0,34 закрытых и 6,00 открытых бивалентов, а также 29,32 унивалента. У этих гибридов F 1 не наблюдалось мультивалентных ассоциаций, было обнаружено, что в среднем на каждый МПК рассчитывается 6,68 хиазмов. В телофазе II наряду с микроядерными тетрадами были обнаружены и пентады.
Анализ полученных результатов показывает, что хотя виды Ae. kotschyi , Ae. columnaris , Ae. neglecta и Ae. recta , включенные в секцию Aegilops , не являются носителями общего субгенома с линиями мягкой пшеницы, уровень конъюгации у гибридов F 1 ЧФХ между ними несколько выше, для каждого МПК составляет 7 штук. Однако, хотя гибриды F 1 , принадлежащие комбинациям 171ACS × Ae. kotschyi и 171ACS × Ae. neglecta , были в некоторой степени фертильными, гибриды F 1 , принадлежащие комбинациям 171ACS × Ae. columnaris и 171ACS × Ae. recta , были полностью стерильными.
Все же, как это ни парадоксально, гибриды F 1 последних двух комбинаций показали полную стерильность, несмотря на то, что число хиазмов на каждую МПК во время мейоза составляло 7, наличие 4–5 ЧФХ для каждого МПК в гибридах F 1 , принадлежащих к комбинациям 171ACS × Ae. umbellulata , 171ACS × Ae. geniculata , 172ACS × Ae. triuncialis и 171AC × Ae. biuncialis , смогло спасти эти гибриды от полной стерильности. Это еще раз доказывает, что нет никакой корреляционной связи между уровнем конъюгации инородных хромосом в гибридных организмах F 1 и фертильностью этих F 1 растений, и что высокая степень как способности конъюгации, так и фертильности, возможна не только из-за уровня конъюгации инородных хромосом, но также зависит от характера ядерно-цитоплазматических отношений, экотипа образцов, взятых в качестве родительской формы, влияния факторов окружающей среды
Следует отметить и то, что у всех полученных нами гибридов пшеница-эгилопс мейоз сопровождался рядом нарушений, характерных для инконгруэнтных гибридов: открытыми бивалентами и унивалентами в метафазе I, расщеплением унивалентов на хроматиды, задержками хромосом в основной фазе I и II, неравномерное распределение, хромосомные и хроматидные мостики, формирование трехполярных и мультиполярных клеток, образование микроядерных тетрад и полиад в телофазе I и II, а также формирование многочисленных микроядер и фрагментов.
Список литературы Цитогенетический анализ гибридов F1, полученных от скрещивания линий Triticum aestivum L. с видами Aegilops L
- Chhuneja P., Kaur S., Goel R. K., Aghaee-Sarbarzeh M., Prashar M., Dhaliwal H. S. Transfer of leaf rust and stripe rust resistance from Aegilops umbellulata Zhuk. to bread wheat (Triticum aestivum L.) // Genetic Resources and Crop Evolution. 2008. V. 55. P. 849-859. https://doi.org/10.1007/s10722-007-9289-3
- Coriton O., Barloy D., Huteau V., Lemoine J., Tanguy A. M., Jahier J. Assignment of Aegilops variabilis Eig chromosomes and translocations carrying resistance to nematodes in wheat // Genome. 2009. V. 52. №4. P. 338-346. https://doi.org/10.1139/G09-011
- Jaffar Aghaei M., Naghavi M. R., Taleei A. R., Omidi M. A., Mozafari J. A study of chromosome homology between three Iranian Aegilops species with D genome and bread wheat (T. aestivum) // Iranian Journal of Rangelands and Forests Plant Breeding and Genetic Research. 2007. V. 15. №2. P. 95-112. https://doi.org/10.22092/ijrfpbgr.2007.114957
- Asghar M., Rao A., Farooq S. Evidence of homoeologous relationship between chromosomes of wheat and Aegilops geniculata // Pak. J. biol. Sci. 2001. V. 4. P. 411-413.
- Dvořák J., Zhang H. B., Kota R. S., Lassner M. Organization and evolution of the 5S ribosomal RNA gene family in wheat and related species // Genome. 1989. V. 32. №6. P. 1003- 1016. https://doi.org/10.1139/g89-545
- Inbart-Pompan H., Eilam T., Eshel A. Searching for salt tolerance among wild relatives of wheat: What should we look for? //Australian Journal of Crop Science. – 2013. – Т. 7. – №. 13. – С. 2116-2127. https://doi.org/10.3316/informit.801313484508105
- Quan X., Liang X., Li H., Xie C., He,W., Qin Y. Identification and characterization of wheat germplasm for salt tolerance // Plants. 2021. V. 10. №2. P. 268. https://doi.org/10.3390/plants10020268
- Aycan M., Baslam M., Asiloglu R., Mitsui T., Yildiz M. Development of new high-salt tolerant bread wheat (Triticum aestivum L.) genotypes and insight into the tolerance mechanisms // Plant Physiology and Biochemistry. 2021. V. 166. P. 314-327. https://doi.org/10.1016/j.plaphy.2021.05.041
- Marais F., Marais A., McCallum B., Pretorius Z. Transfer of leaf rust and stripe rust resistance genes Lr62 and Yr42 from Aegilops neglecta Req. ex Bertol. to common wheat // Crop Science. 2009. V. 49. №3. P. 871-879. https://doi.org/10.2135/cropsci2008.06.0317
- Guo X., Huang Y., Wang J., Fu S., Wang C., Wang M., Han F. Development and cytological characterization of wheat–Thinopyrum intermedium translocation lines with novel stripe rust resistance gene // Frontiers in Plant Science. 2023. V. 14. P. 1135321. https://doi.org/10.3389/fpls.2023.1135321
- Mitrofanova O. P., Khakimova A. G. New genetic resources in wheat breeding for increased grain protein content // Russian Journal of Genetics: Applied Research. 2017. V. 7. №4. P. 477-487. https://doi.org/10.1134/S2079059717040062
- Montes M. J., Andrés M. F., Sin E., López-Braña I., Martín-Sánchez J. A., Romero M. D., Delibes A. Cereal cyst nematode resistance conferred by the Cre 7 gene from Aegilops triuncialis and its relationship with Cre genes from Australian wheat cultivars // Genome. 2008. V. 51. №5. P. 315-319. https://doi.org/10.1139/G08-015
- Naghavi M. R., Aghaei M. J., Taleei A. R., Omidi M., Mozafari J., Hassani M. E. Genetic diversity of the D-genome in T. aestivum and Aegilops species using SSR markers // Genetic resources and crop evolution. 2009. V. 56. P. 499-506. https://doi.org/10.1007/s10722-008-9381-3
- Okada M., Yoshida K., Takumi S. Hybrid incompatibilities in interspecific crosses between tetraploid wheat and its wild diploid relative Aegilops umbellulata // Plant molecular biology. 2017. V. 95. P. 625-645. https://doi.org/10.1007/s11103-017-0677-6
- Özgen M., Yildiz M., Ulukan H., Koyuncu N. Association of gliadin protein pattern and rust resistance derived from Aegilops umbellulata Zhuk. in winter Triticum durum Desf // Breeding science. 2004. V. 54. №3. P. 287-290. https://doi.org/10.1270/jsbbs.54.287
- Bocianowski J., Prażak R. Genotype by year interaction for selected quantitative traits in hybrid lines of Triticum aestivum L. with Aegilops kotschyi Boiss. and Ae. variabilis Eig. using the additive main effects and multiplicative interaction model // Euphytica. 2022. V. 218. №2. P. 11. https://doi.org/10.1007/s10681-022-02967-4
- Siddiqui K. A., Jones J. K. Genetic necrosis in Triticum× Aegilops pentaploid hybrids // Euphytica. 1969. V. 18. №1. P. 71-78. https://doi.org/10.1007/BF00021984
- Rakszegi M., Molnár I., Lovegrove A., Darkó É., Farkas A., Láng L., Shewry P. Addition of Aegilops U and M chromosomes affects protein and dietary fiber content of wholemeal wheat flour // Frontiers in Plant Science. 2017. V. 8. P. 1529. https://doi.org/10.3389/fpls.2017.01529
- Sohail Q., Inoue T., Tanaka H., Eltayeb A. E., Matsuoka Y., Tsujimoto H. Applicability of Aegilops tauschii drought tolerance traits to breeding of hexaploid wheat // Breeding science. 2011. V. 61. №4. P. 347-357. https://doi.org/10.1270/jsbbs.61.347
- Yudina R. S., Leonova I. N., Salina E. A., Khlestkina E. K. Change in salt tolerance of bread wheat as a result of the introgression of the genetic material of Aegilops speltoides and Triticum timopheevii // Russian Journal of Genetics: Applied Research. 2016. V. 6. P. 244-248. https://doi.org/10.1134/S2079059716030151
- Горин А. П., Дунин М. С., Коновалов Ю. Б. Практикум по селекции и семеноводству полевых культур. М.: Колос, 1968. 439 с.
- Доспехов Б. А. Методика полевого опыта: (с основами статистической обработки результатов исследований). М. : Альянс, 2011. 350 с.
- Лакин Г. Ф. Биометрия. М.: Высшая школа, 1990. 351 с.
- Паушева З. П. Практикум по цитологии растений. М.: Агропромиздат, 1988. 270 с.
- Тихомирова М. М. Генетический анализ. Л.: Изд-во ЛГУ, 1990. 280 с.
