Дифференциация диких и домашних свиней по аллотипам белков сыворотки крови

Автор: Князев С.П., Никитин С.В., Кириченко А.В., Николаев А.Г., Волох A.M., Савина М.А., Данильченко Н.В., Айтназаров Р.Б., Ермолаев В.И., Юдина О.П.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: В порядке обсуждения
Статья в выпуске: 6 т.40, 2005 года.
Бесплатный доступ
На основе встречаемости аллотипов сыворотки крови в популяциях диких кабанов и домашних свиней оценивали филогенетические взаимосвязи между внутривидовыми палеарктическими формами Sus scrofa. Рассматривается возможность монофилетического происхождения разных пород домашней свиньи от их единого общего предка.
Короткий адрес: https://sciup.org/142133124
IDR: 142133124 | УДК: 636.4:575.86:57.088
Differentiation of wild boars and domestic pigs by blood serum proteins allotypes
On the basis of frequency of blood serum protein allotypes the authors estimated the phylogenetic interrelations between intraspecific pale arctic forms of Sus scrofa. Presented investigations suggest about genetic distance of S. s. ussuricus from other wild and domestic pigs. The possibility of monophyletic origin of different breeds of domestic pig from the sole common ancestor is considered.
Текст научной статьи Дифференциация диких и домашних свиней по аллотипам белков сыворотки крови
Настоящая работа является продолжением начатых нами ранее исследований полиморфизма свиней и кабанов по аллотипам — антигенам белков сыворотки крови (1, 6, 7). Мы оценивали филогенетические взаимосвязи и степень сходства/различия генофонда ключевых континентальных подвидов дикого кабана и коренных европейских, а также современных евро-азиатских пород домашних свиней.
Методика. Объектом исследования служили следующие подвиды кабанов: S. s. scrofa (европейские кабаны) — две выборки из популяции Воронежского государственного биосферного заповедника 1978-1980 ( n = = 37) и 2000-2001 ( n = 21) годов; S. s. attila — две популяции — северопричерноморская (юг Украины, n = 18) и северокавказская (Кавказский государственный заповедник, n = 11); S. s. nigripes — четыре особи из Сары-Челекского заповедника (Киргизия); S. s. sibiricus — четыре особи из Тувы; S. s. ussuricus — девять особей из Приморского края. Коренная европейская форма домашних свиней была представлена 212 особями сванетской породы (из горных районов Грузии); современная форма смешанного евро-азиатского происхождения — 1281 особью пяти пород: крупная белая (пять выборок из разных племенных заводов — 48, 74, 75, 120 и 234 гол.), цивильская (три выборки — 56, 221 и 247 гол.), скороспелая мясная СМ-1 (новосибирская селекция) — две выборки из племенного репродуктора «Тулинское» учхоза Новосибирского государственного аграрного университета (30 и 81 гол. с интервалом почти в 10 лет), брейтовская — 53 гол., кемеровская — 42 гол.
В образцах сыворотки крови свиней методом двойной иммунодиффузии в агарозном геле с помощью специфических антисывороток-реагентов определяли семь аллотипов, детерминируемых генами четырех иммуногенетических систем: AMG ( α -макроглобулин), A2G ( α -2-глобулин), LPB (липопротеин низкой плотности), LPR (липопротеин высокой плотности) (6). Было проанализировано более 1500 образцов сыворотки крови.
Оценку статистической значимости различий исследуемых выборок проводили с помощью суммарного критерия χ 2 (8).
Филогенетические взаимосвязи между исследуемыми формами свиней оценивали с помощью карты, построенной в системе двух главных компонент на осно-100
вании межпопуляционной вариации частоты аллотипов. Этот метод (а не построение дендрограмм) выбран нами в связи с тем, что иерархически упорядоченную кластеризацию (древовидную структуру) можно обоснованно применять лишь при уверенности, что она естественным образом описывает анализируемые группы животных (8). Другими словами, мы применили метод анализа, при котором структура подразделения выборок априорно не задается, так как одной из задач нашего исследования было выяснение именно этой иерархичности популяций.
Генетические дистанции для построения карты в системе двух главных ком- mT, 27
понент определяли по формуле D = I ^ ( p, - q, ) , где т — число феноти-
-
V i =1
пов, p i и q i — частота встречаемости фенотипов в сравниваемых группах (8).
Результаты. В таблице 1 отражена частота встречаемости аллотипов в исследованных популяциях кабанов и свиней, которую определяли по частоте позитивных фенотипов (особей, в сыворотке крови которых обнаружен соответствующий аллотип).
-
1. Частота встречаемости аллотипов сыворотки крови в популяциях диких и домашних форм Sus scrofa L.
Популяция, выборка
Аллотип
AMG1
AMG5
A2G1
LPB3
LPB12
LPB30
LPR1
S. s. scrofa (1980)
0
0,162
0,486
0,595
0,811
0,703
0,459
S. s. scrofa (2001)
0
0,190
0,143
0,476
0,810
0,238
0,190
S. s. attila (северопричерноморская популя-
ция)
0
0,333
0,278
0,278
0,500
0,167
0
S. s. attila (северокавказская популяция)
0
0
0,090
0,273
1,000
0,091
0,273
S. s. nigripes
0
1,000
0
1,000
1,000
1,000
0,750
S. s. sibiricus
0
1,000
1,000
0
1,000
1,000
1,000
S. s. ussuricus
0,222
0,778
0,889
1,000
0
0,444
1,000
S. s. domesticus (аборигенная европейская
0,028
0,415
0,118
0,452
0,948
0,906
0,108
порода)
S. s. domesticus (современные породы)
0,117
0,107
0,140
0,089
0,891
0,168
0,049
-
2. Генетические дистанции между популяциями диких и домашних свиней Sus scrofa L.
Популяция, выборка
S. s.
scrofa
S. s.
scrofa
S. s. attila
S. s. attila
S. s. nig-ripe
S. s. sibi-ricus
S. s. ussu-ricus
S. s. do-mesticus
S. s. domes-ticus
S. s. scrofa (1980)
–
0,649
0,876
0,855
1,145
1,318
1,331
0,635
0,924
S. s. scrofa (2001)
S. s. attila (северопричер-
–
–
0,464
0,380
1,372
1,700
1,611
0,724
0,449
номорская популяция)
S. s. attila (северокавказская
–
–
–
0,690
1,597
1,728
1,571
0,907
0,524
популяция)
–
–
–
–
1,609
1,804
1,864
0,949
0,359
S. s. nigripes
–
–
–
–
–
1,436
1,503
1,039
1,690
S. s. sibiricus
–
–
–
–
–
–
1,556
1,460
1,780
S. s. ussuricus –
S. s. domesticus (аборигенная
–
–
–
–
–
–
1,724
1,904
европейская порода)
–
–
–
–
–
–
–
–
0,887
В таблице 2 представлены генетические дистанции между этими популяциями. По данным оценки все популяции отличаются друг от друга
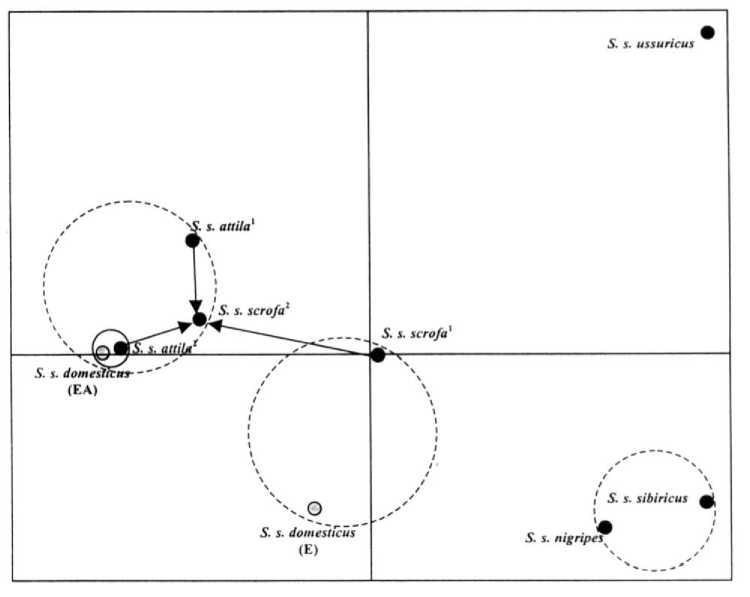
Рис. 1. Карта распределения популяций диких кабанов и различных форм домашних свиней в системе двух главных компонент: S. s. attila 1 — северопричерноморская форма; S. s. attila 2 — северокавказская форма; S. s. scrofa1 и S. s. scrofa 2 — воронежская популяция (соответственно 1980 и 2001 годы); S. s. domesticus (E) — коренная европейская домашняя форма; S. s. domesticus (ЕА) — современная домашняя форма смешанного евро-азиатского происхождения.
с вероятностью ошибки от Р < 0,05 до Р < 0,001, что свидетельствует о достоверности построенной карты филогенетических взаимосвязей между изучаемыми формами свиней (рис. 1).
При анализе карты обращают на себя внимание следующие особенности:
— Удаленность уссурийского подвида кабана S. s. ussuricus от всех других исследованных подвидов диких кабанов и пород домашних свиней и близость сибирского подвида кабана S. s. sibiricus к среднеазиатскому подвиду S. s. nigripes. Эти данные не согласуются с филогенетическими построениями некоторых авторов, в которых восточные кабаны ( S. s. ussuricus, S. s. sibiricus, S. s. moupinensis, S. s. leucomystax, S. s. riukiuanus, S. s. taivanus ) объединены в один таксон (вид, подвид), а западные ( S. s. scrofa, S. s. meridionalis, S. s. algira, S. s. attila, S. s. libicus, S. s. nigripes ) — в другой (10, 11). Однако в большинстве современных сводок по систематике свиней подвиды S. s. ussuricus и S. s. sibiricus все же рассматриваются как безусловно четко дифференцируемые (12). «Отдельность» положения уссурийского подвида кабанов ( S. s. ussuricus ) выявлена также Бромлеем с соавт., которые отмечали весьма специфические экологические и этологические особенности, не характерные для других палеарктических представителей Sus scrofa (13) . Кроме того, скрещивание кабана этого подвида с особями современной евро-азиатской формы домашних свиней (порода ланд-рас) сопровождалось резко выраженным гибридным дисгенезом, дающим основания полагать об их различиях, близких к межвидовому уровню (14).
— Генетическая близость кабанов (S. s. scrofa) Воронежского заповедника (2001 год) к двум популяциям подвида S. s. attila и современной евроазиатской домашней свинье S. s. domesticus и их генетическая удаленность от генофонда кабанов Воронежского заповедника, обследованных в 1978-1980 годах. Вероятно, это может быть обусловлено тем, что в восточно-европейскую часть ареала S. s. scrofa (в данном случае в «островной» Усманский лес Воронежского заповедника) на протяжении последних двух десятилетий происходила миграция с юга представителей подвида S. s. attila. Поэтому современный генофонд кабанов восточно-европейской (воронежской) популяции дает основание рассматривать ее как промежуточную форму между этими двумя подвидами, а сам регион Центрального Черноземья — как возможную зону гибридизации, в которой «южная» форма кабана S. s. attila вытесняет при своей экспансии «северную».
— Относительная близость коренной европейской формы домашней свиньи к современным кабанам Воронежского заповедника. По нашему мнению, этот феномен хорошо объясняет предложенная еще в начале прошлого века гипотеза, согласно которой мелкие, малоплодные, внешне более примитивные короткоухие свиньи коренных пород Европы появились в результате скрещиваний относительно более доместицированных крупных, многоплодных длинноухих свиней с европейским диким кабаном (15). В качестве аборигенной европейской домашней формы мы исследовали короткоухих свиней сванетской породы из горных регионов Грузии, в то время как основой для выведения современных заводских пород Европы послужили длинноухие свиньи маршевой породы (16, 17). Отсюда, вероятно, и близость аборигенной европейской формы домашней свиньи к европейскому дикому кабану S. s. scrofa — расстояние на карте даже короче, чем до современной заводской формы (см. рис. 1).
Особый интерес вызывает близость северокавказского дикого кабана и современной домашней формы свиней, которые явно обособлены от остальных в виде отдельного кластера. При этом последняя значительно удалена от выборки европейского кабана воронежской популяции (1978-1980 годы), которую мы считаем относительно чистой, еще не подвергшейся метизации с S. s. attila, и от коренной европейской домашней формы, а еще больше — от азиатских подвидов кабанов. Такой результат противоречит мнению о том, что дикие формы, чьи ареалы соприкасаются с областью восточно-азиатского центра доместикации, должны быть близки к современным домашним свиньям, выведенным при участии коренных азиатских пород (2, 15, 18-20). Построенная нами карта позволяет предположить, что либо при создании современных пород «улучшате-лями» примитивных свиней Западной Европы были только средиземноморские породы, что не соответствует историческим данным (17, 20), либо предком домашних свиней Юго-Восточной Азии был среднеазиатский подвид кабана S. s. attila или близкий к нему подвид.
На рисунке 2 в виде гистограмм представлены генетические дистанции между аллотипированными нами выборками из популяций диких кабанов и двумя формами домашних свиней — коренной европейской короткоухой (сва-нетская порода) и современной, смешанного евро-азиатского происхождения. Тот факт, что максимальное генетическое сходство проявилось у свиней сванет-ской породы с наиболее удаленной в северо-западном направлении популяцией S. s. scrofa, а не с географически близкой северокавказской популяцией S. s. attila , позволяет предположить, что предки свиней сванетской породы попали на Кавказ из северо-западного или западного региона Европы, а не были одомашнены «на месте» (см. рис. 2). Очевидно, благодаря тому, что свиней сванетской породы содержали в горно-лесных районах, выше области распространения местного дикого кабана (репродуктивная изолированность), у них сохранился генофонд, близкий к первоначальному.
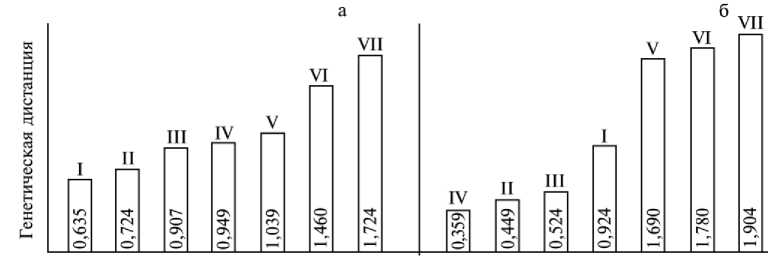
Рис. 2. Генетические дистанции между выборками из популяций диких кабанов палеарктики и двумя формами домашней свиньи: а — европейская аборигенная порода (короткоухие свиньи); б — современные породы смешанного евро-азиатского происхождения. I, II, III, IV, V, VI и VII — соответственно S. s. scrofa (воронежская популяция, 1980 год), S. s. scrofa (воронежская популяция, 2001 год), S. s. attila (северопричерноморская форма), S. s. attila (северокавказская форма), S. s. nigripes , S. s. sibiricus и S. s. ussuri-cus.
Таким образом, проведенное нами исследование свидетельствует, во-первых, о генетической удаленности уссурийских кабанов S . s. ussuricus от прочих диких и домашних форм свиней и, во-вторых, о возможности выдвижения концепции одомашнивания Sus scrofa L., отличной от общепринятой, то есть о происхождении разнообразных форм домашней свиньи от единого прямого предка в одном центре доместикации.
