Динамика формирования пигментного фонда и рост ели в березняке черничном при онтогенезе древостоя

Автор: Зарубина Лилия Валерьевна
Журнал: Молочнохозяйственный вестник @vestnik-molochnoe
Рубрика: Биологические и сельскохозяйственные науки
Статья в выпуске: 2 (22), 2016 года.
Бесплатный доступ
Проведено изучение особенностей роста и динамики накопления фотосинтетических пигментов у подроста ели в березняках черничных разного возраста. Описана методика и приведена краткая характеристика объектов исследования.
Березняк черничный, подрост ели, фотосинтетические пигменты, светособирающий комплекс, хлорофиллбелковый комплекс
Короткий адрес: https://sciup.org/14998843
IDR: 14998843 | УДК: 630*161.4:630*181
Dynamics of pigment fund formation and spruce growth in birch blueberry forest in tree stand ontogeny
The study of growth peculiarities and accumulation dynamics of photosynthetic pigments in spruce undergrowth in birch blueberry forests of different age is made. Methodology is described as well as brief characteristics of experimental objects are given.
Текст научной статьи Динамика формирования пигментного фонда и рост ели в березняке черничном при онтогенезе древостоя
Введение. На Европейском Севере масштабное вовлечение в прошлом столетии хвойных лесов в промышленную эксплуатацию сплошными концентрированными рубками привело к формированию на больших площадях производных лиственных лесов [1]. По данным учета лесного фонда Архангельской области к 2000 г. из общей площади лесов 20,1 млн. га лиственными лесами в области было занято около 20 % всей лесопокрытой площади или 3,9 млн. га, в том числе березовыми – 3,61 млн. га. Среди березняков преобладающим типом являются черничники свежие – 65,9 % [2].
Многочисленные исследования в разных регионах [3, 4] показывают, что под пологом лиственных лесов имеется молодняк ели в количестве (более 10–16 тыс./ га), достаточном для того, чтобы в будущем обеспечить ее господство в составе формирующегося древостоя.
В то же время, несмотря на высокую продуктивность березняков, под их пологом имеется количество подроста ели (более 5 тыс. экз. на 1 га) достаточное для того, чтобы обеспечить ее господство в составе формирующихся древостоев при условии создания для нее благоприятных экологических условий [4].
Многие ученые отмечают успешное предварительное возобновление ели на первых этапах формирования производных лесов и связывают его с достаточно высокой теневыносливостью этой древесной породы [5]. Однако при дальнейшем формировании березового древостоя уменьшение проникающего под полог света и тепла приводит к ухудшению состояния ели, сокращению ее популяции. Поэтому изучение особенностей формирования подроста ели на разных возрастных этапах формирования березового древостоя, в определенной мере, позволит решить практические вопросы по сохранению ели как резерва для восстановления коренных еловых древостоев. Основной причиной слабого роста ели в березняках является то, что корни березы обладают большей энергией роста по сравнению с хвойными породами. На участках с одинаковым качеством лесорастительных условий корневая система березы гуще, и она проникает в почву глубже, чем корневая система хвойных деревьев, вытесняя ее в верхние горизонты. При одновременном поселении с хвойными породами береза быстрее осваивает почву и быстрее восстанавливает листовой аппарат. К 30-летнему своему возрасту, береза развивает корневую систему, значительно превосходящую корневую систему сосны, лиственницы и ели. Вследствие этого верхние слои почвы остаются перегруженными ее корнями. Глубокое и более активное горизонтальное распространение корневых систем березы в сравнении с елью и другими хвойными породами создает превосходящий отток влаги из почвы в березу, препятствует формированию корней хвойных [6].
Корни являются не только органом снабжения растений питательными веществами и водой, но и местом синтеза весьма сложных специфических веществ, таких как пигменты, некоторые фитогормоны, нуклеопротеиды, аминокислоты и др. [7]. Пигменты зеленого листа служат фотосенсибилизаторами лучистой энергии, индикаторами условий местопроизрастания растения, его возрастной динамики.
Изучению пигментного комплекса у древесных растений посвящено значительное число публикаций. Интерес к данной проблеме вызван особой ролью хлорофилла и каротиноидных пигментов зеленого листа в связи с их участием в фотосинтезе в качестве главных фоторецепторов. Считается, что уровень концентрации зеленых и желтых пигментов в фотосинтезирующих органах растения может быть использован в качестве индикатора, определяющего потенциальную фотосинтети- ческую продуктивность вида и его реакцию на динамику факторов внешней среды, степени адаптации к меняющимся экологическим условиям, скорости ассимиляции атмосферного углерода [8]. В связи с этим определенный интерес представляло изучение динамики накопления хлорофилла и каротиноидных пигментов в хвое подроста ели при онтогенезе березняка черничного на разных стадиях его формирования, изучение особенностей их сезонной динамики. На Севере такие исследования до последнего времени не проводились.
Цель исследования состояла в изучении динамики формирования фонда фотосинтетических пигментов и роста подроста ели при возрастной (сукцессионной) смене березняка черничного.
Объекты и методы исследования. Исследования проводились в 2004—2005 гг. в Архангельской области на стационарных объектах Северного научно-исследовательского института лесного хозяйства «Ломовое» и «Холмогорская», дополнительные исследования – на временных пробных площадях. Объектом исследований являлся подрост ели (Picea abies Karst. (Pinacea) в березняке черничном (Betuletum myrtillosum) разного возраста (табл.1).
Таблица 1. Лесоводственно-таксационная характеристика исследуемых участков
|
Древостой |
Подрост |
|||||||
|
состав |
средние |
полнота |
бонитет |
общий запас, м3/ га |
состав |
кол- во, тыс. экз./га |
высота, м |
|
|
Н, м |
Д, см |
|||||||
|
1-летняя вырубка |
||||||||
|
- |
- |
- |
- |
- |
- |
8Е2Б |
2,6 |
1,48 |
|
8- летний березняк |
||||||||
|
10Б |
4,3 |
3,1 |
1,0 |
II |
23 |
10Е |
1,9 |
0,43 |
|
13- летний березняк |
||||||||
|
9Б1Ос |
4,8 |
2,6 |
1,0 |
III |
20 |
10Е |
2,7 |
0,55 |
|
23- летний березняк |
||||||||
|
9Б1Е+Ос |
10,9 |
6,2 |
0,95 |
III |
61 |
10Е |
3,1 |
0,61 |
|
33- летний березняк |
||||||||
|
8Б2Ос1Е |
12,3 |
10,8 |
0,92 |
III |
108 |
10Е |
3,3 |
0,73 |
|
53- летний березняк |
||||||||
|
7Б3Ос |
16,1 |
14,2 |
0,89 |
IV |
132 |
10Е |
3,6 |
0,72 |
|
65- летний березняк |
||||||||
|
7Б3Ос+С |
15,3 |
18,1 |
0,81 |
IV |
170 |
7Е3Б |
5,2 |
1,86 |
|
80- летний березняк |
||||||||
|
6Б3Е1Ос |
18,7 |
22,3 |
0,70 |
IV |
208 |
8Е2Б |
6,2 |
2,21 |
Закладка пробных площадей велась с учётом требований ОСТ 56-69-83 [9]. Перечёт подроста проводился методом пробных площадей (ПП) с учётом требований ГОСТ 16128-70 [10]. Освещённость внутри древостоев измерялась в период полного облиствения березы и осины в конце июня — в июле на высоте 1,5 м с помощью двух люксметров Ю-116М по 35—40 постоянным точкам в околополуденные часы (13 час.) одновременно в лесу и на открытом месте [11]. Содержание пигментов в хвое подроста ели изучалось в экстрактах из свежих листьев спектрофотометрическим методом на СФ-46М ленинградской фирмы «ЛОМО» [12], содержание хлорофилла в светособирающем и хлорофилл-белковых комплексах фотосистем I и II энергопреобразующей системы хлоропластов – по методике А.Б. Рубина [13].
Результаты и их обсуждение. Результаты исследования светового режима на изучаемых нами объектах показали, что наибольшее количество света еловый подрост получает на свежих вырубках и в 1–8-летних березняках, когда возобновляющиеся лиственные породы для подроста еще не создают серьёзного верхнего отенения. В 13-летних березовых насаждениях береза и осина уже начинают оказывать подросту ели определенную конкуренцию за солнечную радиацию. В около-полуденные часы ясных безоблачных дней июня—июля в условия северотаёжной подзоны под полог такого насаждения проникает около 65 % приходящей к кронам верхнего полога солнечной радиации. При дальнейшем разрастании и укреплении эдафических позиций, береза и осина начинают вытеснять ель во второй ярус, где условия для ее возобновления и роста становятся благоприятными. Продолжительность дневного освещения для подпологовой ели значительно сокращается. Уже к 23-летнему возрасту березы в дневное время под полог проникает не более 10– 12 % приходящей солнечной радиации. Установлено [11], что для формирования перспективного подроста ели необходимый минимум освещенности не должен опускаться ниже 25–40 %. В березняках старшего возраста, несмотря на некоторое возрастание изреженности крон полога, световые условия для подпологовой ели остаются неблагоприятными и не способствуют ее нормальному росту и развитию [14].
Пигменты, как составная часть фотосинтетического аппарата, учеными часто используются в качестве диагностического показателя для определения реакции растений на воздействия внешней среды, для оценки их продуктивности и количественного фотосинтетического связывания атмосферного углерода [8, 15].
Функциональное состояние ассимиляционного аппарата зависит не только от освещенности и температурного режима местообитания, но и от многих других внешних и внутренних факторов и, прежде всего, от наличия в листе фотосинтетических пигментов. Прямыми опытами А.А. Красновским (1974) доказано сильное форматирующее действие света на внутренние механизмы, обеспечивающие нормальную работу фотосинтезирующей клетки и активную работу ее пигментной системы [16].
В области низкой освещенности количество зеленых пигментов в листе увеличивается линейно с повышением освещенности, а в области высокой освещенности и при похолодании – уменьшается. Считается, что причиной снижения концентрации пигментов у деревьев при повышенной освещенности и похолодании является деградация внутренней структуры пластид и молекул хлорофилла [17]. Однако, вследствие увеличения ассимилирующей массы, общее содержание пигментов у деревьев в условиях хорошей освещенности всегда выше, чем на затененных участках.
Исследования фонда фотосинтетических пигментов у подроста ели при онтогенезе березняка черничного показали (табл. 2), что наименьшая концентрация зеленых пигментов в его хвое содержится на однолетней вырубке (0,40 мг хлорофилла а и 0,15 мг хлорофилла b на 1 г свежей хвои) и соответствует максимальной освещенности. Это в 1,5–2 раза меньше, чем в спелом березняке. По мере поселения и разрастания на вырубке березы в результате ухудшения световых условий концентрация пигментов в хвое подроста ели начинает постепенно увеличиваться. Уже в 23-летнем возрасте березняка концентрация зеленых пигментов в хвое ели значительно повышается и становится на 29 % выше, чем у подроста на однолетней вырубке. Особенно активно в этот период накапливается хлорофилл b.
Таблица 2. Динамика накопления пигментов у подроста ели при возрастном формировании березняка черничного
|
Возраст березового древостоя, лет |
Пигменты, мкг |
|||||||
|
хлорофилл а |
хлорофилл b |
а+ b |
а: b |
каротиноиды |
ССК |
ХБК |
ССК ХБК |
|
|
1 |
395 |
120 |
515 |
3,29 |
110 |
264 |
251 |
1,05 |
|
8 |
400 |
156 |
556 |
2,56 |
140 |
343 |
213 |
1,61 |
|
13 |
420 |
168 |
588 |
2,50 |
150 |
370 |
218 |
1,70 |
|
23 |
450 |
200 |
650 |
2,25 |
150 |
440 |
210 |
2,10 |
|
33 |
550 |
225 |
775 |
2,44 |
120 |
495 |
280 |
1,77 |
|
53 |
670 |
260 |
930 |
2,58 |
120 |
572 |
358 |
1,60 |
|
65 |
718 |
295 |
1013 |
2,43 |
135 |
649 |
364 |
1,79 |
|
80 |
700 |
290 |
990 |
2,41 |
121 |
638 |
352 |
1,89 |
Наблюдаемое в этот период снижение в хвое отношения Хл.а/Хл.b (с 3,2–3,0 до 2,4–2,5) указывает на ослабление позиций ели по отношению к сопутствующим породам и ухудшение ее жизненного состояния, обуславливаемое недостатком световой энергии, а также возможной переориентацией метаболических процессов с синтеза клетчатки на биосинтез дополнительного количества пигментов, необходимых для улавливания рассеянной радиации. При дальнейшем увеличении возраста березы содержание хлорофилла у подроста ели продолжает увеличиваться. Максимальное количество пигментов в хвое ели накапливает в спелых березняках. В 53-летнем березняке у подроста ели в однолетней хвое содержится 650–690 мкг хлорофилла а и 250–295 мкг хлорофилла b. Увеличение возраста березы до 80 лет уже мало влияет на концентрацию зеленых пигментов у подроста ели. Это можно объяснить тем, что недостаток света и тепла в спелых древостоях ингибирует у ели дыхание корней, снижая их энергетический потенциал.
Также нами были проведены расчеты компонентов энергопреобразующего комплекса пластид (светособирающего и хлорофиллбелковых комплексов фотосистем I и II) у подроста ели на разных стадиях формирования березняка черничного и соотношений между ними (ССК:ХБК). Интерес к проведению таких расчетов обусловлен тем, что была установлена обратная статистически значимая закономерность между продуктивностью растения и величиной отношения ССК:ХБК в его ассимилирующих органах. Отмечается, что чем выше значение этого отношения, тем менее продуктивным является вид и сорт растения и тем меньше мембранных комплексов электротранспортной сети содержится в его клеточных структурах [13].
Наши расчеты показали, что в 1–8-летних березняках подрост ели, несмотря на низкое содержание пигментов, имеет самую высокую интенсивность фотосинтеза. Отношения ССК:ХБК у подроста в насаждениях такого возраста не превышают 1,05–1,60. Начиная с 13-летнего возраста березы в связи с ухудшением световых условий отношения между светособирающим и хлорофилл-белковыми комплексами у подроста повышаются до 1,77–2,10, что может свидетельствовать об утрате подростом лидирующих позиций. В 53–80-летних березняках интенсивность фотосинтеза у подроста ели, несмотря на высокое содержание пигментов, самая низкая. Отношения между светособирающим и хлорофилл-белковыми комплексами самые высокие (2, 17). Фонд каротиноидных пигментов у подроста ели при онтогенезе березняка черничного меняется мало. Это связано, видимо, с необходимостью по- стоянной защиты хлорофилла от неблагоприятных внешних воздействий: от яркого солнечного освещения в летний период и низких температур в зимний период.
Обусловленность жизненного состояния от световых условий у подроста ели проявилась и в скорости роста главного и бокового побегов (рис. 1).
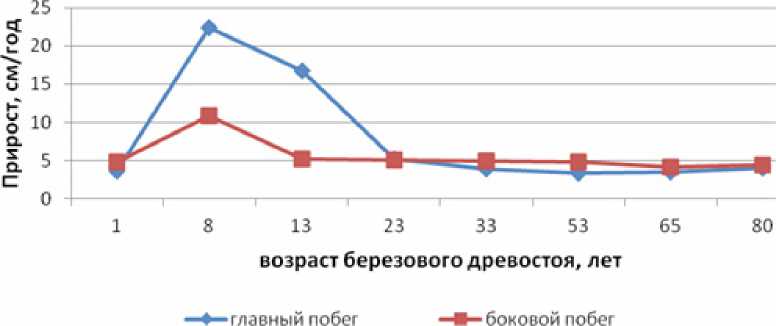
Рисунок 1. Прирост главного и бокового побегов у подроста ели в березняках черничных разного возраста.
Известно, что появившись под пологом насаждений, молодые елочки с раннего возраста приспосабливаются к ограниченному режиму почвенного и светового питания и в соответствии с ними формируют свой прирост. Об этом свидетельствуют и наши исследования. Как видно из рисунка 1, обусловленность состояния светового режима в изучаемых насаждениях от их возрастного состояния у подроста ели проявилась в скорости роста в высоту и величине биометрических характеристик. Результаты исследований показали, что у подроста средней высоты прирост в высоту в березовом насаждении в зависимости от его возраста изменяется от 22,4±0,3 см до 3,5±0,5 см. Величина прироста боковых побегов также изменяется, но при более низких абсолютных значениях (от 10,9±3,6 см до 4,4±0,5 см). Максимальный прирост в высоту подрост ели имеет в 8-летнем возрасте. В 13-летнем березняке, несмотря на относительно благоприятные для ели экологические условия, прирост верхушечного и боковых побегов у подроста ели по сравнению с 8-летним насаждением оказывается несколько замедленным. Но особенно медленно подрост ели растет под пологом спелых и перестойных насаждений, где конкуренция за свет и элементы питания со стороны лиственных пород наиболее выражена. Уже при достижении березняком 23-летнего возраста ежегодный прирост верхушечного побега у ели не превышает 5,6 см. Дальнейшее увеличение возраста березы уже слабо влияет на рост ели. В 65-летнем березняке черничном у среднего подроста среднепериодический прирост в высоту снижается до 3,5±0,18 см, прирост боковых побегов до 4,1±0,16 см.
Заключение. Таким образом, результаты исследований свидетельствуют, что самые низкие концентрации хлорофилла и каротиноидных пигментов в хвое подроста ели в березняке черничном содержатся в начальные периоды формирования его на вырубке. В это время подрост ели имеет наибольшую продуктивность по созданию органического вещества (углеводов). На это указывают и наиболее низкое отношение ССК:ХБК в хлоропластах. Начиная с 13-летнего возраста березы, когда световые условия для подроста постепенно начинают ухудшаться в связи с образо- ванием верхнего оттеняющего полога, в его хвое происходит достаточно быстрое возрастание концентрации, как хлорофилла, так и каротиноидных пигментов. При дальнейшем увеличением возраста березы фонд фотосинтетических пигментов в хвое подроста уже слабо увеличивается, достигая максимальных значений при достижении березой возраста спелости. Следовательно, в берёзовых насаждениях более молодого возраста (8—13 лет) ель обладает повышенной способностью к ассимиляции СО2 и созданию органического вещества, чем в насаждениях более старших возрастов. Это подтверждается более активным ростом подроста ели в высоту.
Список литературы Динамика формирования пигментного фонда и рост ели в березняке черничном при онтогенезе древостоя
- Мелехов, И.С. Леса Архангельской и Вологодской области/И.С. Мелехов, В.Г. Чертовской, Н.А. Моисеев//Леса СССР. Т.1. -М.: Наука, 1966. -С. 78-156.
- Трубин, Д.В. Динамика и перспективы лесопользования в Архангельской области/Д.В. Трубин, С.В. Третьяков, С.В. Коптев//Архангельск: АГТУ, 2000. -96 с.
- Побединский, А.В. Рациональное использование лесных богатств России/А.В. Побединский//Лесное хоз-во. -2002. -№6. -С. 2-5.
- Вялых, Н.И. Способы рубок главного пользования и лесовозобновление на Европейском Севере (Материалы финско-российского семинара по лесовосстановлению. Финляндия, Вуокатти, 28.92.10. 1998)/Н.И. Вялых//Бюллетень НИИ леса. Финляндия, 2000. -С. 25-31.
- Дружинин, Ф.Н. Надземная фитомасса подпологовой ели после комплексных рубок/Ф.Н. Дружинин//Известия высших учебных заведений. Лесной журнал. -2012. -№3. -С. 19-24.
- Бобкова, К.С. Еловые леса // Биопродукционный процесс в лесных экосистемах Севера / Бобкова К.С.// СПб. :Наука, 2001. - С. 52-68.
- Зарубина, Л.В. Влияние прореживания и азота на сезонную динамику дыхания корней сосны и ели/Л.В. Зарубина, В.Н. Коновалов//Известия высших учебных заведений. Лесной журнал. -2016. -№1. -С 100-115.
- Тужилкина, В.В. Хлорофилловый индекс и ежегодный сток углерода в еловые фитоценозы/В.В. Тужилкина, К.С. Бобкова//Биопродукционный процесс в лесных экосистемах. -СПб.: Наука, 2001. -С. 203-207.
- ОСТ 56-69-83. Пробные площади лесоустроительные. Метод закладки. -М.: ЦБПТИ Гослесхоза СССР, 1984. -60 с.
- ГОСТ 16128-70. Площади пробные лесоустроительные. Метод закладки. -М.: Изд-во стандартов, 1971. -23 с.
- Алексеев, В.А. Световой режим леса/В.А. Алексеев. -М.: Наука, 1975. -280 с.
- Шлык, А.А. Определение хлорофиллов и каротиноидов в экстрактах зеленых листьев/А.А. Шлык//Биохимические методы в физиологии растений. -М.: Наука, 1971. -С. 154-169.
- Рубин, А.Б. Регуляция первичных стадий фотосинтеза при изменении физиологического состояния растений/А.Б. Рубин, П.С.Венедиктов, Т.Е.Кренделева//Фотосинтез и продукционный процесс. -М.: Наука, 1988. -С. 29-39.
- Зарубина, Л.В. Эколого-физиологические особенности ели в березняках черничных/Л.В. Зарубина, В.Н. Коновалов//Архангельск: С(А)ФУ, 2014. 378 с.
- Цельникер, Ю.Л. Хлорофилловый индекс как показатель годичной аккумуляции углерода древостоями леса/Ю.Л. Цельникер, И.С. Малкина//Физиология растений -1994. -Т. 41. Вып. 3. -С. 325-330.
- Красовский, А.А. Преобразование энергии света при фотосинтезе. Молекулярные механизмы/А.А. Красовский//XXIX Баховские чтения. -М.: Наука, 1974. 64 с.
- Ходасевич, Э.В. Фотосинтетический аппарат хвойных/Э.В. Ходасевич//Минск: Наука и техника, 1982. -199 с.
