Динамика экологического состава микротериофауны из плейстоценовых отложений в южной галерее Денисовой пещеры

Автор: Агаджанян А.К., Шуньков М.В., Козликин М.Б.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология каменного века палеоэкология
Статья в выпуске: т.XXIX, 2023 года.
Бесплатный доступ
На основе репрезентативной фаунистической коллекции, включающей более 16 тыс. костных остатков, дается оценка динамики экологического состава сообщества мелких позвоночных в окрестностях Денисовой пещеры во второй половине среднего верхнем плейстоцене. Палеонтологические материалы представляют тафоценозы из слоев 19.1-11 в центральной части южной галереи пещеры. Согласно составу основных экологических групп животных, а также стратиграфическому положению в разрезе, время формирования слоя 19.1 предварительно можно отнести к концу тобольского или началу самаровского времени рубежа МИС 9-8. В тафоценозах из слоев 18 и 17 основные группы мелких позвоночных обладают близкими характеристиками, указывающими на относительно теплые климатические условия, которые соответствуют, скорее всего, ширтинскому межледниковью МИС 7. Экологический состав сообщества мелких позвоночных из отложений слоев 16.2 и 16.1 свидетельствует об ухудшении природной обстановки на фоне нестабильного режима влажности, что соответствует, видимо, разным фазам тазовского оледенения МИС 6. Следующий этап в развитии природной среды в долине Ануя представляют фаунистические материалы из литологических слоев 15-13, время накопления которых в целом укладывается в рамки МИС 5. Состав тафоценоза слоя 12 свидетельствует о редукции лесной растительности и расширении площади степных биотопов при общем ухудшении природной обстановки в эпоху ермаковского похолодания МИС 4. Сочетание в животном сообществе слоя 11 белки и многочисленной популяции степных пеструшек предполагает распространение лесостепных ландшафтов в условиях относительно теплого и сухого климата, который может соответствовать эпохе каргинского межстадиала МИС 3.
Горный алтай, денисова пещера, плейстоцен, мелкие позвоночные, тафоценоз, таксономический состав, экологические группы
Короткий адрес: https://sciup.org/145146623
IDR: 145146623 | УДК: 569+903.32 | DOI: 10.17746/2658-6193.2023.29.0007-0011
Dynamics of ecological composition of microtheriofauna from the Pleistocene deposits in the south chamber of Denisova Cave
This article discusses dynamics of ecological composition of small vertebrate communities near Denisova cave in the second half of the Middle-Upper Pleistocene using a representative faunal collection of over 16,000 bone remains. Paleontological evidence represents taphocenoses of layers 19.1-11 in the central part of the South Chamber in the cave. According to the composition of main ecological groups of animals as well as stratigraphic position in cross-section, layer 19.1 emerged in the Late Tobol or Early Samarovo period at the boundary of MIS 9 and 8. Main groups of small vertebrates had similar features in taphocenoses of layers 18 and 17, indicating relatively warm climate which most likely corresponds to the Shirta interglacial MIS 7. Ecological composition manifested by the community of small vertebrates from the deposits of layers 16.2 and 16.1 indicates deterioration of natural environment and unstable humidity conditions, which may correspond to different phases of the Taz glaciation MIS 6. The next stage in the development of natural environment in the Anui River valley is manifested by the faunal evidence from lithological layers 15-13, accumulated in MIS 5. Composition of taphocenosis in layer 12 shows reduced forest vegetation and expansion of the area of steppe biotopes with general deterioration of natural environment during the Ermakovo cooling MIS 4. The combination of squirrels and large population of steppe voles in the animal community of layer 11 suggests the spread offorest-steppe landscapes in relatively warm and dry climate which may correspond the Kargin interstadial MIS 3.
Текст научной статьи Динамика экологического состава микротериофауны из плейстоценовых отложений в южной галерее Денисовой пещеры
В ходе комплексных исследований плейстоценовой толщи в южной галерее Денисовой пещеры получена представительная коллекция костных остатков мелких позвоночных, включающая более 30 тыс. экз., по которым был установлен таксономических состав тафоценозов из слоев 19.1–11 [Агаджанян, Шуньков, Козликин, 2022]. Дальнейшее изучение этих материалов позволило уточнить видовую принадлежность некоторых таксонов, климатостратиграфическую позицию вмещающих отложений и природные условия их формирования. Для оценки динамики экологического состава микротериофауны в высоком разрешении при по строении графиков из анализа были исключены костные остатки летучих мышей, птиц и рыб, общая доля которых по слоям составляет от 25 до 60 %. В итоге структура выборочного состава ископаемых сообществ мелких позвоночных проанализирована по 16 669 образцам (см. рисунок,таблицу ).
В сообществе мелких млекопитающих из слоя 19.1 присутствуют землеройки рода Sorex, крот Asioscalops, рыжие полевки Clethrionomys, бурундук Cricetus crisetus и белка-летяга Pteromys – показатели относительно благоприятных природных условий. В то же время в тафоценозе самой представительной является популяция скальных полевок Alticola, отмечены узкочерепная полевка Stenocranius gregalis, суслик Spermophilus и сурок Marmota. Два последних вида впадают на зиму в спячку, снижая температуру тела и интенсивность обмена веществ, что позволяет им дожить до весны и быть менее зависимыми от низких температур и промерзания почвы. Благодаря этим особенностям суслики по долинам рек распространяются на север до полярного круга, а сурки заселяют высокогорные плато. В силу биологических и анатомических особенностей они не могут жить в условиях высокотравных лугов с плотной дерновиной, предпочитая открытые ландшафты с разреженным травостоем в условиях холодного климата. Следует отметить также находки костей лемминга рода Lemmus – бореального компонента фауны, относительно низкую численность цокора Myospalax и отсутствие слепушонки Ellobius – грызуна, живущего под землей и питающегося подземными частями растений. Слепушонка и цокор активны круглый год и не могут существовать в условиях долговременного и глубокого промерзания почвы. Их отсутствие или малое количество в тафоценозе отражает интенсивные криогенные процессы в грунтах. В целом, согласно составу основных экологических групп микротериофауны, а также стратиграфическому положению в разрезе, время формирования слоя 19.1 предварительно можно отнести к концу тобольского или началу самаровского времени, т.е. к рубежу МИС 9–8.
В тафоценозах из слоев 18 и 17 основные группы мелких позвоночных обладают близкими характеристиками. В их составе отмечены бурозубки Sorex , крот, суслики, слепушонка. Стабильна численность степных пеструшек Lagurus и полевок – плоскоче-
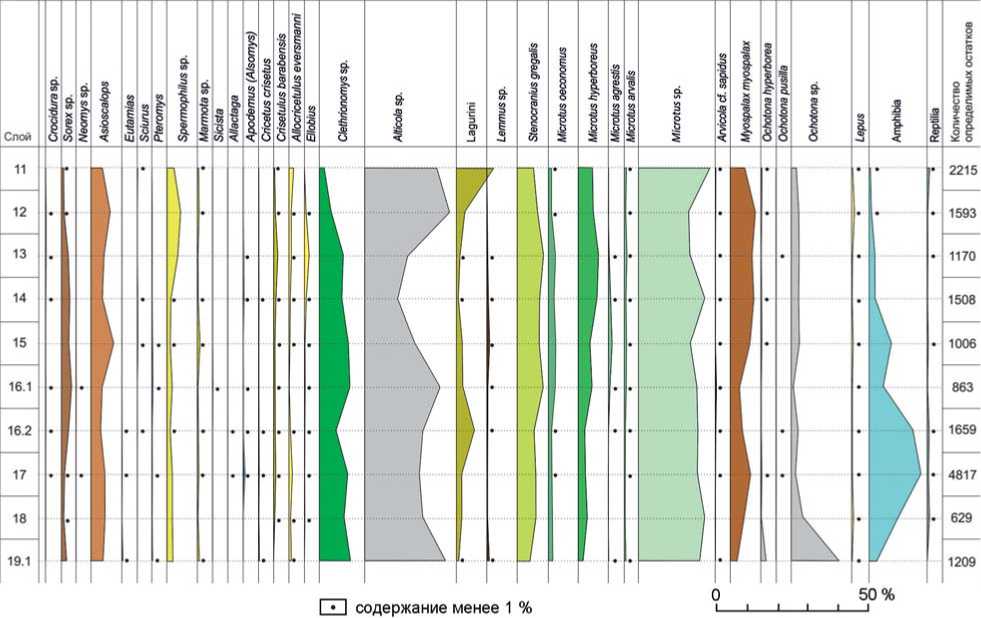
Диаграмма выборочного состава мелких позвоночных из плейстоценовых отложений в южной галерее Денисовой пещеры.
Выборочный состав мелких позвоночных из плейстоценовых отложений в южной галерее Денисовой пещеры
|
>s о и |
Ol |
1 |
1 |
о |
я |
о |
1 |
eq |
0 |
1 |
04 |
8 |
0 |
1 |
1 |
1 |
1 |
40 |
о |
1 |
Я |
Я |
40 ОО |
Я |
04 ОО |
О |
40 |
1 |
0 |
OI |
01 |
1 |
0 |
||||
|
Я |
Ol |
in |
Ol |
я |
eq |
Я |
я |
co |
ОО |
я |
40 |
40 |
04 |
о |
OI |
OO |
co |
01 |
40 |
00 04 |
01 |
||||||||||||||||
|
eq >s о t; U |
04 |
о |
oo |
s |
0 |
ОО ОО |
eq 00 |
о |
40 |
о |
of |
о |
О |
0 |
40 Ol^ |
40 |
04 40 |
eq |
|||||||||||||||||||
|
2 |
OO |
Ol |
о |
eq 04 |
40 |
n |
2 |
Я |
ОО |
04 |
01 |
eq 40 |
04 OI |
01 |
Я |
О |
'—1 |
^r |
|||||||||||||||||||
|
>s о ч U |
^ |
о |
гр |
40 |
04 |
о |
о |
я |
of |
40 |
eq 01^ оГ |
S |
04 04. ОО |
О |
0 |
s |
О |
OO OO |
40 of |
40 |
0 |
0 |
|||||||||||||||
|
40 |
Ol |
я |
я |
eq |
я |
04 |
ОО |
я |
Я |
40 eq |
40 40 |
я |
Ol |
—1 |
l> |
—1 |
OO 04 |
О |
Я |
^Г- |
04 |
||||||||||||||||
|
>s о t; U |
П |
о |
о |
40 40 |
04 |
OO |
0 |
0 |
о |
о |
О |
$ |
eq 04 |
04 04 |
ОО |
О |
0 |
Я |
О |
40 ОО |
я |
О |
|||||||||||||||
|
2 |
Ol |
о |
Ol |
OO |
eq |
04 |
—1 |
04 |
ш |
04 |
04 eq |
Я |
Я |
о |
Ol |
01 |
^r- |
О |
ОО Ol |
04 ОО |
О1 |
in |
|||||||||||||||
|
>s о ч U |
2 |
о |
04 |
40 04 40 |
Я |
0 |
о |
04 04 |
s |
я |
04 of |
О |
ОО ОО |
40 04 |
о |
0 |
s |
0 |
О |
40 04 40 |
ОО 40 of |
ОО |
04 О!^ |
s |
|||||||||||||
|
Ol |
co |
40 |
О |
eq |
—1 |
^ |
О |
40 |
OI |
OI |
OI |
я |
Я |
04 |
S |
о |
co |
40 |
l> |
l> |
О |
Я |
40 |
Я |
40 |
||||||||||||
|
40 >s о U |
'—1 |
Ol |
Ol |
Ol |
of |
Ol |
2 |
Я |
О |
eq |
Ol |
я |
я |
of |
04 |
ОО |
ОО ОО |
40 |
0 |
OO |
00 |
Ol |
Я oo |
of |
04 О!^ |
ОО |
40 |
||||||||||
|
о |
—1 |
—1 |
eq eq |
—1 |
Я |
Ol |
04 |
—1 |
OI |
OI |
01 |
Я |
о |
40 |
40 |
co |
>n |
l> |
—1 |
я |
01 |
Я |
in |
^r |
|||||||||||||
|
eq 40 >s о t; U |
^ |
04 |
о |
Ol |
s |
о |
Я of |
о |
0 |
40 40 |
Ol |
О |
О |
о |
о |
40 40 |
ОО |
О1 |
о. |
^ 40 |
eq of |
04 40 |
01 40 |
О |
я^ |
01 |
я |
01 |
Ol |
||||||||
|
oo |
Ol |
о |
oo |
^ |
'—1 |
Ol |
—1 |
—1 |
ш |
'—1 |
m |
2 |
1> |
я |
о |
01 |
OO Ol |
^r- |
s |
—1 |
я |
01 |
я |
01 |
Ol |
||||||||||||
|
>s о ч U |
^ |
r~ |
о |
3 |
о |
3 |
40 40 |
о |
0 |
о |
0 |
0 |
О |
Я |
О |
о |
о |
40 |
О |
0 |
of |
^ |
40 |
п |
0 of |
Я |
0 |
04 |
я |
я |
of |
О |
|||||
|
40 |
Ol |
’Г. |
Ol |
Ol |
co |
04 |
^ |
—1 |
^ |
04 |
'—1 |
—1 |
|> |
ш |
eq eq |
ОО |
^Г- |
О |
я |
40 О1 |
eq 00 |
я |
^ |
OO |
Ol 00 01 |
Ol |
0 |
CO |
|||||||||
|
oo >s о t; U |
^ |
^ |
04 |
V |
eq |
04 |
40 |
40 |
04 |
40 40 |
я |
oo |
04 |
00 00 |
of |
||||||||||||||||||||||
|
^ |
n |
40 eq |
eq |
n |
—1 |
^Г- |
—1 |
О |
о |
Я |
00 |
co |
40 |
я |
|> |
^ |
|||||||||||||||||||||
|
04 >s о t; U |
^ |
m |
40 |
Я |
0 |
0 |
eq |
О |
о |
о |
40 40 |
я |
04 04 |
^ |
ОО 04. |
04 04 |
0 |
О |
OO |
04 oo |
04 04 |
Ol |
0 |
0 |
|||||||||||||
|
2 |
Ol |
2 |
Ol |
—1 |
40 |
n |
OI |
ОО |
ОО |
Я |
О |
ОО ОО |
Ol eq |
—1 |
^t- |
l> |
я |
Ol |
40 |
01 |
^r |
||||||||||||||||
|
о H |
Рн Q 7§ о 6 |
О О p 1 |
3 3 i |
3 к p О |
Ph 1 |
Ph t |
1 8 Я |
8 i |
3 s |
t p я |
3 в ^ g |
Ph ,3 1 g Cl |
Рн G О g i |
G Я |
G b/j Л Я |
t 0 н о |
5 |
3 3 0 |
8 р -В 0 |
В К Q 1 > ^ Я |
р t 0 и |
3 к р ,о р' t 0 и |
Рн t 0 и |
я о -Id р я я |
Ph -5 Я |
p b/j 3 s b/j Q О |
8 •2 8 p 3 s b/j Q О |
Ph 3 s b/j Q О |
Ph 3 0 |
b/j р b/j ■i р о |
3 8 0 к р о 3 р i| |
3 р -1 3 р i| |
р b/j Q 3 р i| |
Q 3 p |
|||
Окончание таблицы
Экологиче ский состав сообщества мелких позвоночных из отложений слоев 16.2 и 16.1 включает землероек Soricidae, крота, а среди древесных беличьих – бурундука, белку Sciurus vulgaris и летягу. Зарегистрирован тушканчик Allactaga , что предполагает нарушение сплошного травяного покрова и появление локальных участков сухих степей. Нестабильна доля лесных полевок и степных пеструшек, а скальные полевки достигают очередного пика численности. Вновь появляется лемминг Lemmus – представитель бореальных биотопов. Об ухудшении природной обстановки свидетельствует также относительно низкая численность цокора и пищух Ochotona . Характер фаунистических остатков из этих отложений в целом отражает обстановку относительно холодного климата, с нестабильным режимом влажности, что соответствует, видимо, разным фазам тазовского оледенения МИС 6.
Следующий этап в развитии природной среды в долине Ануя представляют фаунистические материалы из литологических слоев 15–13. Для тафоцено-за из слоя 15 характерны стабильно высокая численность лесных полевок, присутствие белки и летяги, а среди насекомоядных – максимально высокая доля крота. Относительно низкое количество барабинского хомячка и хомяка Эверсманна указывает на редукцию злакового разнотравья за счет, скорее всего, увеличения площади лесных массивов. Уменьшается доля сусликов и полевок – плоскочерепной, узкочерепной и северосибирской – обитателей скальных или сухих биотопов. Распространение среди Microtini полевки-экономки, пашенной и обыкновенной полевок отражает достаточно влажные условия, а присутствие лемминга предполагает развитие ассоциаций зеленых мхов. О достаточно благоприятной климатической обстановке свидетельствует возросшая численность цо-кора и лягушек. В целом состав мелких позвоночных из этого слоя отражает условия относительно теплого и влажного климата, отвечающие, скорее всего, казан-цевскому межледниковью МИС 5е.
В составе мелкой фауны из слоя 14 снижается доля крота, рыжих полевок и лягушек, что связано, видимо, с редукцией лесных массивов и уменьшением об- щей влагообеспеченности. В то же время сохраняется стабильная численность сусликов, хомяка Эверсманна и барабинского хомячка – типичных представителей степных биотопов, а также возрастает число северосибирской полевки, предпочитающей условия бореального климата. Эти экологические показатели свидетельствуют о некотором понижение температурного режима и уменьшении влажности климата.
Таксономический состав мелких позвоночных из литологического слоя 13 в целом достаточно близок тафоценозу из нижележащего слоя. Вместе с тем в составе мелкой фауны из отложений слоя 13 немного увеличивается численность крота и лесных полевок Clethrionomys , присутствует лесная мышь, а ба-рабинский хомячок и слепушонка достигают своего максимального количества. Возрастает доля сусликов – свидетельство расширения участков злакового разнотравья, а стабильная численность популяции серых полевок предполагает существование луговых и пойменных ассоциаций.
Близкие экологические характеристики фаунистического сообщества из толщи слоев 14 и 13 указывают на относительно теплые климатические условия в эпоху их формирования, при этом природная обстановка во время накопления слоя 13 была более благоприятной для развития лесных и луговых биотопов в рамках, видимо, второй половины МИС 5.
В тафоценозе из отложений слоя 12 снижается количество землероек до минимального значения, вдвое падает численность Clethrionomys и слепушонки, что указывает на существенное сокращение площади ле сных массивов и участков высокотравных лугов. В то же время растет число скальных полевок и степных пеструшек, относительно стабильными остаются популяции хомяка Эверсманна и барабин-ского хомячка – представителей открытых биотопов. Возрастает численность крота и цокора – активных землероев, для которых предпочтительны многоснежные зимы с неглубоким промерзанием грунта. Древесные беличьи отсутствуют, а в составе наземных беличьих увеличивается доля сусликов и сурка, отражающая распространение степных ландшафтов с разреженным травостоем. Эти данные свидетельствуют о редукции лесной растительности и расширении площади степных биотопов при общем ухудшении природной обстановки в эпоху ермаковского похолодания МИС 4.
В структуре тафоценоза из литологического слоя 11 среди древесных беличьих отмечена белка, что предполагает наличие локальных лесных участков. Увеличивается численность степной пеструшки и хомяка Эверсманна, при этом снижается количество рыжих, плоскочерепных и узкочерепных полевок. Сочетание в животном сообществе белки и многочис- ленной популяции степных пеструшек предполагает распространение лесостепных ландшафтов в условиях относительно теплого и сухого климата. На относительно аридные условия при достаточно благоприятном температурном режиме указывают также уменьшение доли крота и цокора и минимальное количество лягушек. Согласно этим данным, формирование слоя 11 может соответствовать эпохе каргинско-го межстадиала МИС 3.
Таким образом, динамика таксономического состава мелких позвоночных из плейстоценовых отложений в южной галерее Денисовой пещеры отражает изменения природных условий во второй половине среднего и на протяжении верхнего плейстоцена. Реконструированы эпохи холодного климата, которые соответствуют, возможно, началу самаровского, тазов-скому и ермаковскому похолоданиям в климатостратиграфической схеме плейстоцена Западной Сибири. Относительно теплые эпохи отвечают, возможно, финалу тобольского, ширтинскому и казанцевскому межледниковьям, а также каргинскому межстадиалу. При этом анализ разных экологических групп мелких млекопитающих позволил установить локальные особенности в развитии каждой эпохи, характерные непосредственно для окрестностей пещеры.
Исследование выполнено по проекту НИР ИАЭТ СО РАН № FWZG-2022-0003 «Северная Азия в каменном веке: культурная динамика и экологический контекст».
Список литературы Динамика экологического состава микротериофауны из плейстоценовых отложений в южной галерее Денисовой пещеры
- Агаджанян А.К., Шуньков М.В., Козликин М.Б. Динамика сообществ мелких позвоночных из плейстоценовых отложений южной галереи Денисовой пещеры // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. - Новосибирск: Изд-во ИАЭТ СО РАН, 2022. - Т. XXVIII. - С. 7-12. EDN: QIZORL
