Динамика природных популяций Henosepilachna vigintioctomaculata Motchulsky, 1857 (Coleoptera: Coccinellidae) в Приморском крае
 в Приморском крае")
Автор: Мацишина Наталия Валериевна, Фисенко Птр Викторович, Ермак Марина Владимировна, Собко Ольга Абдулалиевна
Журнал: Овощи России @vegetables
Рубрика: Агрохимия, агропочвоведение, защита и карантин растений
Статья в выпуске: 1 (69), 2023 года.
Бесплатный доступ
Актуальность. Динамика популяции (волны жизни) - неотъемлемое свойство популяционных систем. Эти колебания могут индуцироваться различными факторами - условиями и ресурсами среды или элементами ценоза того же или более высокого уровня - конкурентами, паразитами, хищниками, болезнями. Малоизученная до сих пор в условиях Дальнего Востока России динамика популяции H. vigintioctomaculata необходима для выяснения причин, по которым картофельная коровка стала вредителем интродуцированных культурных растений. Материалы и методы. Для учета имаго все участки в районе исследования обследовались с интервалом в восемь дней в течение всего периода исследования. Все участки обследовались в течение одного дня. Все имаго, взятые руками на растениях-хозяевах, были помечены индивидуально при первой поимке. Маркировка обычно проводилась путем прокалывания надкрылий швейной иглой (0,4 мм в диаметре) по Hirano. Жуки нового поколения (через два-три дня после появления) были помечены маникюрным лаком, поскольку трудно было пометить иглой без травм. При повторном отлове этих жуков снова помечали иглой. Ни один из этих методов не оказал вредного влияния на выживание помеченных имаго. Жуков выпускали на то же растение, на котором они были пойманы. Учетные данные были проанализированы по методу, описанному Jolly. Результаты. Перезимовавшие имаго появились на картофельных полях и начали откладывать яйца в конце мая. Имаго первого поколения появились в конце июня - начале июля. Имаго второго поколения появились в конце июля - начале августа, часть которых мигрировала в места зимовки без яйцекладки примерно в конце августа. Выход перезимовавших жуков отмечался на дубе монгольском (Quercus mongolica), чистотеле большом (Chelidónium május), черемухе обыкновенной (Prunus padus). Типичный первичный агробиотоп заселения картофельной коровкой представлял собой посадки картофеля в фазе 5-7 листьев, окруженные межевой полосой с пыреем (Elytrigia repens), одуванчиками (Taraxacum officinale) и осотом (Sonchus arvensis). Маркирование перезимовавших и имаго нового поколения показало, что биотопическая приуроченность картофельной коровки зависит от агрометеорологических условий
Картофельная коровка, динамика популяции, зимовка, картофель, приморский край
Короткий адрес: https://sciup.org/140297782
IDR: 140297782 | УДК: 632.763.79(571.63) | DOI: 10.18619/2072-9146-2023-1-80-86
The dynamics of wild populations of Henosepilachna vigintioctomaculata Motch. (Coleoptera:Coccinellidae) in Primorsky krai
Relevance. Population dynamics (population waves) is an intrinsic quality of population systems. These fluctuations can be induced by various factors, e.g. environmental conditions and resource availability, elements of a food chain from the same or a higher level (competitors, parasites, predators), and diseases. Few researchers have addressed the issue of the population dynamics of Henosepilachna vigintioctomaculata in the Russian Far East. This paper investigates the reasons why the potato ladybird beetle became a pest of introduced cultivated plants. Materials and methods. To collect data on imagines, all research sites were examined at intervals of eight days during the entire period of the study. The examination of the research sites was conducted within one day. All the imagines taken from host plants were marked at the first encounter by puncturing their elytra with a sewing needle (0.4 mm in diameter) according to Hirano. The next generation of beetles was marked with nail polish (in two-three days after emergence) to avoid traumatizing the insects. At the second encounter, these beetles were also marked by puncturing their elytra with a needle. The methods implemented did not have an adverse effect on the survivability of the studied imagines. The beetles were released to the same plants where they had been taken from. The research data were analyzed by Jolly's method. Results. Overwintered imagines appeared in the fields and began ovipositing at the end of May. The first generation of imagines appeared at the end of June - the begging of July. The second generation of imagines could be observed at the end of July - the beginning of August. A part of the imagines migrated to overwintering sites without ovipositing approximately at the end of August. The overwintered beetles were observed to emerge on the Mongolian oak Q uercus mongolica, the greater celandine Chelidonium majus, and the bird cherry Prunus padus. A typical primary biotope of the potato ladybird beetle was a potato field at the stage of vegetative growth (5-7 leaves) surrounded by the couch grass Elymus repens, the common dandelion Taraxacum officinale, and the perennial sow-thistle Sonchus arvensis. Marking the overwintered and new generations of imagines showed that the presence of the potato ladybird beetle in an ecosystem depended on agricultural and agrometeorological conditions.
Текст научной статьи Динамика природных популяций Henosepilachna vigintioctomaculata Motchulsky, 1857 (Coleoptera: Coccinellidae) в Приморском крае
Relevance. Population dynamics (population waves) is an intrinsic quality of population systems. These fluctuations can be induced by various factors, e.g. environmental conditions and resource availability, elements of a food chain from the same or a higher level (competitors, parasites, predators), and diseases. Few researchers have addressed the issue of the population dynamics of Henosepilachna vigintioctomaculata in the Russian Far East. This paper investigates the reasons why the potato ladybird beetle became a pest of introduced cultivated plants.
Materials and methods. To collect data on imagines, all research sites were examined at intervals of eight days during the entire period of the study. The examination of the research sites was conducted within one day. All the imagines taken from host plants were marked at the first encounter by puncturing their elytra with a sewing needle (0.4 mm in diameter) according to Hirano. The next generation of beetles was marked with nail polish (in two-three days after emergence) to avoid traumatizing the insects. At the second encounter, these beetles were also marked by puncturing their elytra with a needle. The methods implemented did not have an adverse effect on the survivability of the studied imagines. The beetles were released to the same plants where they had been taken from. The research data were analyzed by Jolly’s method.
Results. Overwintered imagines appeared in the fields and began ovipositing at the end of May. The first generation of imagines appeared at the end of June – the begging of July. The second generation of imagines could be observed at the end of July – the beginning of August. A part of the imagines migrated to overwintering sites without ovipositing approximately at the end of August. The overwintered beetles were observed to emerge on the Mongolian oak Q uercus mongolica , the greater celandine Chelidonium majus , and the bird cherry Prunus padus . A typical primary biotope of the potato ladybird beetle was a potato field at the stage of vegetative growth (5-7 leaves) surrounded by the couch grass Elymus repens , the common dandelion Taraxacum officinale , and the perennial sow-thistle Sonchus arvensis . Marking the overwintered and new generations of imagines showed that the presence of the potato ladybird beetle in an ecosystem depended on agricultural and agrometeorological conditions.
Д инамика популяции (волны жизни) — неотъемлемое свойство популяционных систем [1, 2]. Сообщается, что эти колебания могут индуцироваться различными факторами — условиями и ресурсами среды (температура, осадки, доступность и качество пищи, т.н. “bottom-up effects”) или элементами ценоза того же или более высокого уровня — конкурентами, паразитами, хищниками, болезнями (т.н. “top-down effects”) [3] и их можно классифицировать в соответствии с самыми разными принципами. Так, выделяют динамику многолетнюю и сезонную; стабильную, флуктуирующую и взрывную (эруптивную); колебания закономерные, т.е. повторяющиеся с определенной периодичностью, и случайные, которые не имеют явно выраженной периодичности [4]. Среди фитофагов рода Henosepilachna полевые популяционные исследования были впервые проведены для H. vigintioctomaculata [5] и H. pustulosa [6] в климатических условиях Японии. Малоизученная до сих пор в условиях Дальнего Востока России динамика популяции H. vigintioctomaculata необходима для выяснения причин, по которым картофельная коровка стала вредителем интродуцированных культурных растений.
Материалы и методы
Модельный участок, на котором проводились исследования популяционной динамики, представляет собой четырехугольник с углами, находящимися в координатах 43.860944, 131.974475; 43.857302, 131.987698; 43.842997, 131.975218; 43.846855, 131.962886 (рис.1).
Участок был разделен на шесть типичных биотопов питания и размножения, представляющих собой частные дачные угодья, где преобладающим видом является Solanum tuberosum. Биотопы обозначались арабскими цифрами для удобства обследования (1-6) (табл. 1). Для учета имаго все участки в районе исследования обследовались с интервалом в восемь дней в течение всего периода исследования. Все участки обследова- лись в течение одного дня. Все имаго, взятые руками на растениях-хозяевах, были помечены индивидуально при первой поимке. Маркировка обычно проводилась путем прокалывания надкрылий швейной иглой (0,4 мм в диаметре) по Hirano (7.). Жуки нового поколения (через два-три дня после появления) были помечены маникюрным лаком, поскольку трудно было пометить иглой без травм. При повторном отлове этих жуков снова помечали иглой. Ни один из этих методов не оказал вредного влияния на выживание помеченных имаго. Жуков выпускали на то же растение, на котором они были пойманы.Учетные данные были проанализированы по методу, описанному Jolly [8] и Seber [9].

Рис.1. Расположение модельного участка на карте
Figure 1. The position ofthe modelsite on a map
Таблица 1. Описание модельных биотопов Table 1. The description of the model biotopes
|
Порядковый номер |
Площадь |
Типичная растительность |
|
1 |
100 м2 |
Solanum tuberosum, Elytrígia répens, Ambrósia artemisiifólia, Taraxacum officinale, Cirsium setosum, Sonchus arvensis, Solanum nigrum, Solánum melongéna, Capsicum annuum, Solánum lycopérsicum, Cucumis sativus, Cucurbita pepo |
|
2 |
250 м2 |
Solanum tuberosum, Convolvulus arvensis, Hibiscus trionum, Humulopsis scandens, Elytrígia répens, Ambrósia artemisiifólia, Taraxacum officinale, Solánum melongéna, Capsicum annuum, Solánum lycopérsicum, Cucumis sativus, Cucurbita pepo |
|
3 |
150 м2 |
Solanum tuberosum, Solánum melongéna, Capsicum annuum, Solánum lycopérsicum, Cucumis sativus, Elytrígia répens, Ambrósia artemisiifólia, Pisum sativum, Glycine max, Brassica oleracea, Acalypha australis |
|
4 |
320 м2 |
Solanum tuberosum, Acalypha australis, Elytrígia répens, Ambrósia artemisiifólia, Taraxacum officinale, Cirsium setosum, Sonchus arvensis, Solanum nigrum, Echinochloa crus-galli, Trifolium pretense, Equisetum arvense |
|
5 |
200 м2 |
Solanum tuberosum, Eriochloa villosa, Chenopodium album, Cirsium arvense var. integrifolium, Commelina communis, Zea mays |
|
6 |
100 м2 |
Solanum tuberosum, Ambrósia artemisiifólia |
Результаты и обсуждение
В наших исследованиях,по среднемноголетним данным, перезимовавшие имаго появились на картофельных полях и начали откладывать яйца в конце мая.Имаго первого поколения появились в конце июня -начале июля. Имаго второго поколения появились в конце июля – начале августа,часть которых мигрировала в места зимовки без яйцекладки примерно в конце августа. Время выхода жуков с мест зимовки зависит от климатических особенностей местности и погодных условий года. А.Н.Иванова [10],исходя из многолетних наблюде-ний,пришла к выводу,что вылет жуков картофельной коровки с мест зимовки происходит при температуре не ниже 13-15ºС. По данным О.Н. Горелова и Л.Ф. Ламеко [11] выход жуков из зимовки наблюдается,когда сумма положительных температур в среднем достигнет 340°,а средняя температура воздуха 13-14ºС (10-20 мая).Эти температуры обеспечивают начало развития вида. Обычно время выхода жуков с мест зимовки длится 2-3 недели.Это объясняется тем,что стации зимовки,распо-ложенные в лесу, прогреваются неодинаково.Первое время жуки держатся на различных деревьях и кустарниках. В наших исследованиях выход перезимовавших жуков отмечался надубе монгольском ( Quercus mongolica ), чистотеле большом ( Chelidónium május ), черемухе обыкновенной ( Prúnus pádus ) (рис.2).В целом,зимовоч-ные биотопы картофельной коровки представляют собой широколиственные лесные насаждения с кустарниковым подлеском и разнотравьем,сухие и хорошо прогреваемые солнцем (рис.2).А.И.Куренцов [12] отмечал,что жуки картофельной коровки держатся в большом количестве на цветущем в это время растении селезеночнике волосистом ( Chrysosplechnium pilosum Maxim .). Они сидят по нескольку экземпляров на одном растении, выедая цветы,почки. Жуки встречаются на тех экземплярах этого растения, которые растут по краям лесосек, по сторонам лесных дорог,там,где больше солнечного света.Весенние колонии жуков часто встречаются на опушках тех лесов,которые вплотную подходят к огородам и полям. Жуки вначале встречаются на цветущих в это время древесных породах в основном розоцветных (Rosaceae): черемухе азиатской ( Padus asiatica ),яблоне маньчжурской ( Malus manshurica ), боярышнике Максимовича ( Crategus maximowiczii ),груше уссурийской ( Pyrrus ussuriensis ), лесных травах и затем переходят на более характерные и часто встречающиеся сорные растения полей: осот полевой ( Sonchus arvensis L.),пас-лен черный ( Solarium nigrum L.).
Жуки проходят дополнительное питание пыльцой на цветках черемухи,яблони,боярышника.С последних растений коровка переходит на картофель, но может питаться листьями огурцов,томатов, баклажана. Перелет жуков с опушек леса на картофельные поля происходит с конца третьей декады мая [13,14].В наших исследова-ниях,переход на картофельные поля проходил в течение всего периода выхода имаго из диапаузы (рис. 3). Дополнительного питания пыльцой нами зафиксировано не было.Картофельные поля заселялись при этом нерав-номерно,в зависимости от преобладающей погоды.
Спустя 3-5 дней после выхода коровки из зимовки,на растениях картофеля отмечались спаривающиеся жуки (рис.3-1) и первые повреждения (рис.3-2).Типичный первичный агробиотоп заселения картофельной коровкой представлял собой посадки картофеля в фазе 5-7 листь-ев,окруженные межевой полосой с пыреем (Elytrigia repens),одуванчиками (Taraxacum officinale) и осотом (Sonchus arvensis).
Принято считать,что перезимовавшие имаго и имаго первого поколения являются находящимися в репродуктивной стадии [15].Некоторые из них доживают до периода появления следующего поколения,и не маркированных невозможно отличить от новых.Кроме того, необходимо отметить, что места зимовки для картофельной коровки достоверно не установлены.
По результатам наших исследований,исследований Л.А.Михайловой [16] выделяются три фенологические группы коровок:
-
1) Жуки, массово мигрирующие осенью к определенным участкам леса, где они, вероятно,остаются в под-стилке,что доказать не удалось. Однако,по данным Л.А. Михайловой,жуки этой группы образуют значительные скопления в местах зимовки.Указанная группа наиболее тесно связана с лесными насаждениями;
-
2) Менее многочисленная, мигрирующая в лес незначительными партиями группа коровок,которая уходит на зимовку позднее и распределяется в лесной подстилке рассеянно;
-
3) Не мигрирующая группа,зимующая в местах пита-ния.Это жуки последнего периода отрождения,питаю-щиеся на огородах до заморозков.
2. Henosepilachna vigintioctomaculata на растениях чистотела
Поскольку первые две группы представлены в популяции коровки наиболее широко,полагаем уместным считать зимовочными стациями лесные,а места зимовки – в пределах тех растений,на которых жук обнаруживается весной.
В целом, маркирование перезимовавших и имаго нового поколения показало,что биотопическая приуроченность картофельной коровки зависит от агрометеорологических условий.В дождливом и прохладном 2019 году наибольшее количество имаго отмечено в наиболее сухих, хорошо прогреваемых биотопах №№ 1,3,6 (рис.4, 5).Соотношение полов «самка: самец» у перезимовавших имаго составило 1,14:1 для биотопа №1,1,44:1 – для биотопа №2, 2,06:1–для биотопа №3,2,13:1–для биотопа №4,2,07:1–для биотопа №5,3,75:1–для биотопа №6, идентифицировать удалось всех особей (рис.4). У имаго нового поколения это же соотношение составило 1,105:1 для биотопа №1,2,46:1 – для биотопа №2; 1,42:1 – для биотопа №3; 2,3:1 – для биотопа №4,1,57:1 – для биотопа №5 и 1,5:1–для биотопа №6.Тренд наувеличение численности самок подтверждает мнение,что самки более выносливы и адаптированы к неблагоприятным усло-виям,нежели самцы [15].Тенденция к преобладанию числа самок над числом самцов сохранилась и в 2020 году,однако общая численность перезимовавших имаго в среднем снизилась, что связано с замоканием мест зимовки и частичной гибелью.
Соотношение полов самка:самец у перезимовавших имаго в 2020 г для биотопа №1 составило 1,8:1,для биотопа №2 – 1,88:1,для биотопа №3 – 1,89:1,для биотопа №4 – 2,72:1, для биотопа №5 -1,66:1,для биотопа №6– 1,31:1 (рис.6). Сниженная численность перезимовавших имаго не привела к снижению количества имаго нового поколения. Напротив,по сравнению с 2019 г. отмечается достоверная прибавка населения для всех биотов за исключением №6(рис.7). Соотношение самок к самцов

1. Henosepilachna vigintioctomaculata на листьях дуба
(43,85326°С, 131,97147°В)
-
1. Henosepilachna vigintioctomaculata on oak leaves
(43,85326°С, 131,97147°В)
(43,85326°С, 131,97147°В)

3. 43,85326°С, 131,97147°В
Имаго картофельной коровки на листьях крапивы 3. An imago of the potato ladybird beetle on leaves of the stinging nettle
(43,85326°С, 131,97147°В)
-
2. Henosepilachna vigintioctomaculata on a perennial sow-thistle
4. 43,85326°С, 131,97147°В
5. 43,85326С, 131,97147В
6. 43,85326С, 131,97147В
7. 43,85326°С, 131,97147°В Зимовочная стация #1
8. 43,85326°С, 131,97147°В Зимовочная стация #2
Имаго картофельной коровки на листьях клёна 4. An imago of the potato ladybird beetle on maple leaves

Имаго картофельной коровки на бересклете 5. An imago of the potato ladybird beetle on Euonymus plants
Имаго картофельной коровки на черемухе 6. An imago of the potato ladybird beetle on a bird cherry

7. Overwintering site #1
8. Overwintering site#2

10. 43,85326 ° С, 131,97147 ° В Зимовочная стация #4
10. Overwintering site #4
9. 43,85326°С, 131,97147°В Зимовочная стация #3
9. Overwintering site #3
Рис. 2. Зимовочные стации картофельной коровки
Figure 2. Overwintering sites ofthe potato ladybird beetle

1. Спаривание картофельной коровки на картофеле (43,85278°С, 131,96936°В)
1. Mating potato ladybird beetles on potato (43,85278°С 131,96936°В)
2. Первые повреждения картофеля (43,85269°С, 131,96933°В)
2. The first lesions on potato plants (43,85269°С, 131,96933°В)

3. Типичный первичный биотоп картофельной коровки: первые всходы картофеля, межевая полоса с пыреем, одуванчиками и осотом (43,85280°С, 131,96926°В)
3. A typical primary biotope of the potato ladybird beetle: the first sprouts, a field boundary with the perennial sow-thistle, the common dandelion, and the couch grass (43,85280°С, 131,96926°В)
Рис.3. Стации расселения картофельной коровки
Figure 3. Colonization sites ofthe potato ladybird beetle
у нового поколения в 2020 г составило 2,62:1 для биотопа №1,3,7:1 – для биотопа №2, 1,55:1 – для биотопа №3, 3,23:1 для биотопа №4,1,78:1 – для биотопа №5,2,2:1 – для биотопа №6.
Несмотря на то,что осень 2020 года была благоприятна для ухода на зимовку,неконтролируемые палы травы и подлеска весны 2021 года привели к снижению численности зимующих имаго, а для биотопов 5 и 6– полному отсутствию весеннего заселения (рис.8). Численность нового поколения в этих биотопах складывалась из расселительной и репродуктивной активности имаго биотопов №№4 и 5,о чем свидетельствуют данные маркировки (рис.9). Соотношение полов у перезимовавших имаго составило 1,41:1 в биотопе №1,1,5:1 – в биотопе №2,1,66:1 – в биотопе №3,1,83:1 – в биотопе №4 для «самка: самец» соответственно.
Соотношение полов «самка: самец» для имаго нового поколения составило в биотопе №1 – 4:1,0,57:1 – в биотопе №2,2,2:1 – в биотопе №3,2,4:1 – в биотопе №4,2,4:1 – в биотопе №5,2,14:1 – в биотопе №6(рис. 9).Анализ процентного соотношения самок в изучаемых выборках показывает несостоятельность постулатов Л.Ф. Радыгиной об эффективности применения хемостерили-
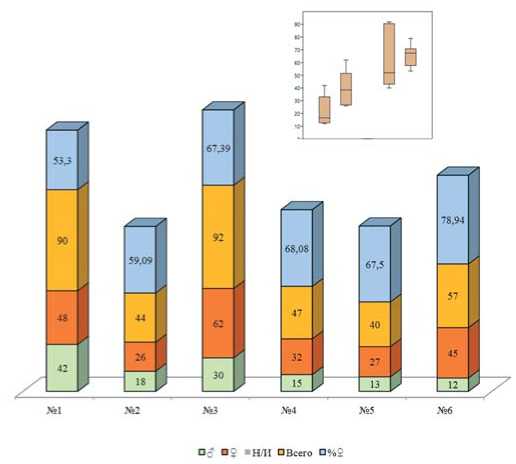
Рис. 4. Общее количество помеченных перезимовавших имаго и соотношение полов,2019 год;Н/И – не идентифицирован пол
Figure 4. The totalnumberofthe marked overwintered imagines and the proportion between sexes,2019;N/I– non-identified sex
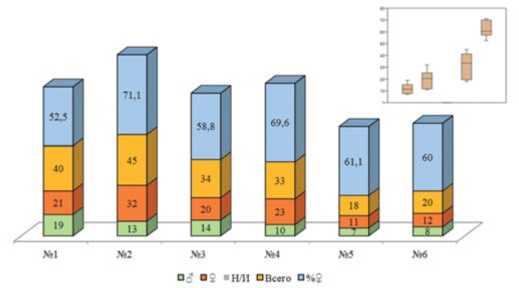
Рис.5. Общее количество помеченных имаго нового поколения и соотношение полов,2019 год. Примечание: 1-6 – номера биотопов;Н/И – не идентифицирован пол
Figure 5. The totalnumberofthe marked imagines from the new generation and the proportion between sexes,2019. Note: 1-6 – the numbers ofthe biotopes;N/I– non-identified sexes
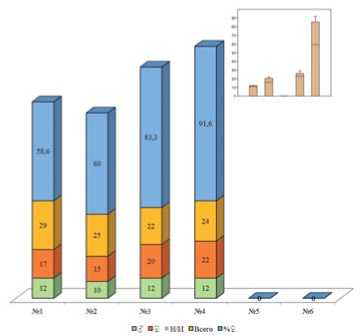
Рис.8. Общее количество помеченных перезимовавших имаго и соотношение полов,2021 год. Примечание: 1-6 – номера биотопов;Н/И – не идентифицирован пол
Figure 8. The totalnumberofthe marked overwintered imagines and the proportion between sexes,2021. Note: 1-6 – the numbers ofthe biotopes;N/I– non-identified sexes.
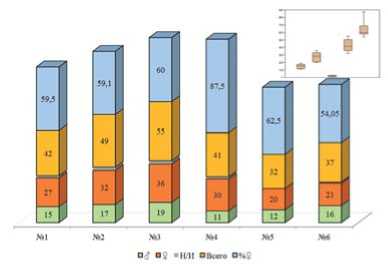
Рис.6. Общее количество помеченных перезимовавших имаго и соотношение полов,2020 год. Примечание: 1-6 – номера биотопов;Н/И – не идентифицирован пол
Figure 6. The totalnumberofthe marked overwintered imagines and the proportion between sexes,2020. Note: 1-6 – the numbers ofthe biotopes;N/I– non-identified sexes
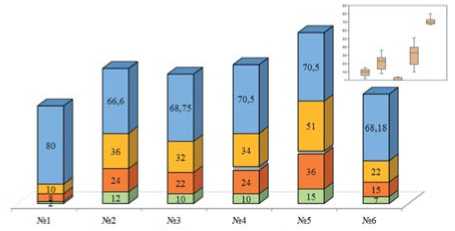
Ос OS "Н/И О Вс «о ОНх
Рис.9. Общее количество помеченных имаго нового поколения и соотношение полов,2021 год. Примечание: 1-6 – номера биотопов;Н/И – не идентифицирован пол
Figure 9. The totalnumberofthe marked imagines from the new generation and the proportion between sexes,2021. Note: 1-6 – the numbers ofthe biotopes;N/I– non-identified sexes
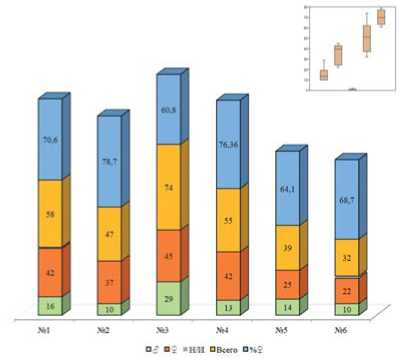
Рис.7. Общее количество помеченных имаго нового поколения и соотношение полов,2020 год. Примечание: 1-6 – номера биотопов;Н/И – не идентифицирован пол
Figure 7. The totalnumberofthe marked imagines from the new generation and the proportion between sexes,2020. Note: 1-6 – the numbers ofthe biotopes;N/I– non-identified sexes
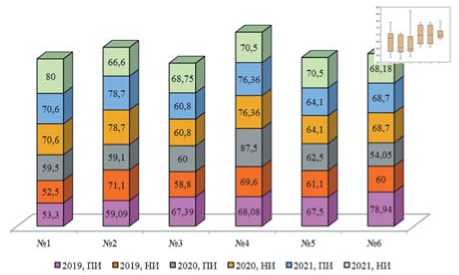
Рис.10. Количество самок (% от общей выборки) в изучаемых биотопах по годам исследований. Примечание: 1-6 номера биотопов;ПИ – перезимовавшие имаго;НИ – имаго нового поколения
Figure 10. The percentage offemale beetles in the studied biotopes overthe years ofresearch. Note: 1-6 – the numbers ofthe biotopes;OV – overwintered imagines;
NI– imagines from the new generation
заторов самцов для борьбы с картофельной коровкой (рис.10) [17,18,19,20]. В своих работах Людмила Федоровна отмечает,что самки картофельной коровки способны носить запас семени одного самца в течение всего периода жизни [19].Учитывая превосходящую численность самок над самцами в популяции,хемостерили-зация самцов не будет иметь экономической эффективности и интересна скорее в модельных исследованиях.
Таким образом, динамика природной популяции H. vigintioctomaculata зависела от агрометеорологических условий: температуры выхода из диапаузы,погод-ных условий ухода на зимовку, соотношения полов «самка:самец», по средне многолетним данным в популяции картофельной коровки преобладало число самок над самцами, а так же антропогенных факторов,которые приводили к гибели насекомых.
Об авторах:
Наталия Валериевна Мацишина – кандидат биол. наук, старший научный сотрудник лаборатории селекционно-генетических исследований полевых культур, ,автор для переписки,
Aboutthe Authors:
Nataliya V.Matsishina – (Biology), SeniorResearcher, the Laboratory of Breeding and Genetic Research on Field Crops, , Correspondence Author,
Marina V.Ermak – JuniorResearcher,the Laboratory ofBreeding and Genetic Research on Field Crops, ,
Petr V.Fisenko – (Biology),Leading researcher, the Laboratory ofBreeding and Genetic Research on Field Crops ,
Olga A.Sobko – Researcher, the Laboratory ofBreeding and Genetic Research on Field Crops, 2nd year Ph.D. student. ,
Список литературы Динамика природных популяций Henosepilachna vigintioctomaculata Motchulsky, 1857 (Coleoptera: Coccinellidae) в Приморском крае
- Чернова Н.М., Былова А.М. Общая экология. Москва: Дрофа; 2004.
- Шилов И.А. Экология. Москва: Высшая школа; 2009.
- Фролов А.Н. Динамика численности и прогноз массовых размножений вредных насекомых: исторический экскурс и пути развития. Аналитический обзор. Вестник защиты растений. 2017;4(94):521.
- Макфедьен Э. Экология животных. Цели и методы. Москва: Мир; 1965.
- Iwao S. Dynamics of numbers of a phytophagous lady-beetle, Epilachna vigintioctomaculata, living in patchily distributed habitats. In: Dynamics of Populations: Proc. Adv. Study Inst. “Dynamics of Numbers in Population”, Oosterbeek, the Netherlands, 7-18 Sept. 1970. Wageningen, Netherlands: H. Veenman & Zonen N.V.; 1971. Р.129-147.
- Nakamura K., Ohgushi T. Studies on the population dynamics of a thlstle-feedlng lady beetle, Henosepilachna pustulosa (KоNo) in a cool temperate climax forest. I. The estimation of adult population parameters by the marking, release and recapture method. Res. Popul. Ecol. 1979;(20):297-314.
- Hirano K. A new marking method using a sewing needle: effects on survival rate of twentyeight-spotted lady-beetles, Henosepilachna vigintioctopunctata Fabricius. Appl. Ent. Zool. 1981;(6):264-270.
- Jolly G.M. Explicit estimates from capture-recapture data with both death and immigrationstochastic model. Biometrika. 1965;(52):225224.
- Seber G.A.F. The estimation of animal abundance and related parameters. London: Griggin; 1973.
- Иванова А.Н. Картофельная коровка на Дальнем Востоке. Владивосток; 1962.
- Горелов О.Н., Ламеко Л.Ф. Прогноз сроков борьбы с картофельной коровкой: информ. листок. Владивосток: Приморский ЦНТИ; 1980.
- Куренцов А.И. Новые данные по биологии картофельной коровки. Труды Горнотаежной станции Дальневосточного филиала Академии наук СССР. Владивосток; 1946;(5):257-266.
- Куренцов А.И. Проблема сельскохозяйственного освоения горнотаежных районов в Приморском крае и вредные насекомые. Труды Горнотаежной станции Дальневосточного филиала Академии наук СССР. Ворошилов-Уссурийский; 1941;(4):15-97.
- Кузнецов В.Н. Эколого-фаунистический обзор кокцинеллид (Coleoptera, Coccinellidae) Дальнего Востока. Фауна и экология беспозвоночных Дальнего Востока (вредители и энтомофаги). Владивосток; 1984. С.25-36.
- Reznik S.Y., Karpun N.N., Zakharchenko V.Y., Shoshina Y.I., Dolgovskaya M.Y., Saulich A.K., Musolin D.L. To everything there is a season: phenology and photoperiodic control of seasonal development in the invasive caucasian population of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae). Insects. 2022;(13):580. https://doi.org/10.3390/insects13070580
- Михайлова Л.А. Динамика численности картофельной коровки в различных экологических условиях юга Приморья. Ботанические и зоологические исследования на Дальнем Востоке. Владивосток; 1968;(2):238-242.
- Радыгина Л.Ф. Биологическое обоснование метода химической стерилизации 28-пятнистой картофельной коровки Epilachna vigintioctomaculata Motsch. в целях снижения её численности: диссертация на соискание ученой степени кандидата биологических наук. Ленинград-Пушкин; 1984.
- Радыгина Л.Ф. Влияние хемостерилянтов на половую активность самцов 28-пятнистой картофельной коровки Epilachna vigintioctomaculata Mötsch. Фауна и экология насекомых Приморского края и Камчатки. Владивосток; 1981. С.121-128.
- Радыгина Л.Ф. Особенности размножения 28-точечной картофельной коровки в Приморском крае. Защита растений в сельском и лесном хозяйствах Дальнего Востока. Уссурийск; 1977;(48):28-30.
- Радыгина Л.Ф. Химическая стерилизация картофельной коровки в Приморском крае. Бюллетень ВНИИЗР. 1980;(47):16-21.
