Динамика старовозрастных еловых лесов Московской области

Автор: Беляева Н.Г., Черненькова Т.В., Маслов А.А.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Биология
Статья в выпуске: 2 т.28, 2025 года.
Бесплатный доступ
Исследована динамика еловых (Picea abies) лесов на территории Московской области за прошедшие четыре десятилетия. Отслежены основные тренды после распада старовозрастных ельников, а также изменение экологических условий местообитаний. Выявлены две разнонаправленных формы вторичных сукцессий: с восстановлением елового подроста и с активным развитием кустарников.
Московская область, еловые леса, сукцессии, долговременные исследования, заповедные лесные участки
Короткий адрес: https://sciup.org/143185069
IDR: 143185069 | УДК: 574.42:502.72:630(470.311) | DOI: 10.31433/2618-9593-2025-28-2-39-43
Dynamics of old-growth spruce forests in the Moscow region
The authors have studied the dynamics of spruce (Picea abies) forests in the Moscow region over the past four decades, and have traced main trends after the decay of old-growth spruce forests, as well as changes in ecological conditions of habitats. It was revealed the two multidirectional forms of secondary successions: with restoration of spruce undergrowth and with active development of shrubs.
Текст научной статьи Динамика старовозрастных еловых лесов Московской области
Старомонетный переулок 29 стр. 4, г. Москва, 119017, e-mail: , ; e-mail: , ;
Советская 21, п/о Успенское, Московская обл., 143030, e-mail: ,
Ель обыкновенная ( Picea abies (L.) Karst.) по своему распространению и хозяйственному значению является одной из основных лесообразующих пород в европейской части России. В зоне хвойно-широколиственных лесов, к которой относится бóльшая часть Московской области, древостои с преобладанием ели рассматриваются во многих случаях как сообщества, сформировавшиеся в результате многовековой деятельности человека. В этой связи большой интерес представляет динамика еловых массивов, сохранившихся с конца XIX века. Исследования проводились на территории заповедных лесных участков Московской области (рис. 1).
Цель работы – выявление изменений состава и структуры старовозрастных ельников в зоне хвойно-широколиственных лесов путем повторных учетов с интервалом в 40 лет. Полученные результаты дадут возможность идентифицировать
динамические тренды ельников естественного происхождения и позволят спрогнозировать их дальнейшее развитие.
В системе лесного хозяйства в соответствии с Лесным кодексом «заповедный лесной участок» имеет официальный природоохранный статус. Режим, установленный в заповедных участках, позволил во многих случаях сохранить старовозрастные еловые леса и производные от них сообщества после массовой гибели ели в результате вспышек короеда типографа в 2011–2013 гг. [1].
В середине 1980-х гг. были выполнены лесо-водственно-геоботанические описания на 56 пробных площадках. Малонарушенные старовозрастные еловые леса представляли собой сомкнутые леса из ели с примесью других пород с преобладающим возрастом древостоев 80–100 лет. В 2024 г. на тех же площадках были выполнены повторные описания. В данной работе проанализированы из-
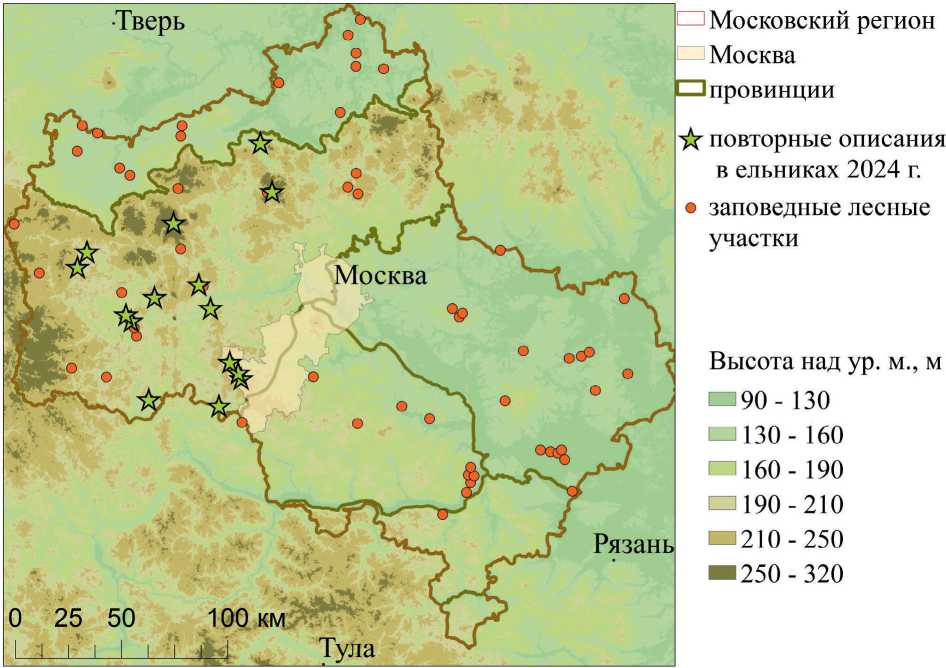
Рис. 1. Район исследований и расположение заповедных лесных участков [2] и повторных описаний в ельниках в 2024 г.
Fig. 1. Location of the strict scientific forest reserves in the study area [2] and repeated relevès in spruce forests in 2024
менения древесного и кустарникового ярусов еловых насаждений за два временных среза.
Спустя 40 лет при сравнении состава и структуры растительности верхних ярусов ельников заметны кардинальные изменения. Главные – это уменьшение сомкнутости древесного яруса (А), в составе которого доминировала ель, и увеличение покрытия кустарникового яруса, в котором начала преобладать лещина ( Corylus avellana ). В частности, среднее проективное покрытие древесного яруса в старовозрастных ельниках сократилось более чем в два раза – с 72 до 31% (рис. 2), в то время как проективное покрытие подлеска (В2) увеличилось с 16 до 40%, а подроста (B1) – c 9 до 24%.
Столь сильные изменения отражают факт полной или частичной гибели материнского елового древостоя, в том числе в результате массовых вспышек короеда типографа 1999–2003 и 2010– 2013 гг. При этом снижение общей сомкнутости древесного яруса происходило не только за счет выпадения ели, но и за счет выпадения других видов: березы (Betula sp.), осины (Populus tremula), сосны (Pinus sylvestris). Участие клена (Acer platanoides) и дуба (Quercus robur), напротив, несколько возросло.
В отличие от древесного яруса, покрытие почти всех видов подроста за 37 лет увеличилось. В старовозрастных ельниках подрост деревьев в 1987 г. был представлен незначительно. На первом месте (с покрытием 5.4%) был подрост ели, в меньших количествах были представлены липа ( Tilia cordata ) и клен. В 2024 г. у подроста ели средние значения покрытия выросли почти в 3 раза (с 5 до 13%), что говорит о потенциально успешном восстановлении данного вида на месте распавшихся древостоев и демонстрирует первый вариант развития растительных сообществ после выпадения древесного полога.
В подлеске самое существенное изменение – сильнейшее развитие лещины, среднее по-

Рис. 2. Бокс-плоты варьирования покрытия древесного яруса (А), подроста (B1) и подлеска (В2) в разные периоды измерения. На рисунке: средние значения и 0.95 доверительный интервал
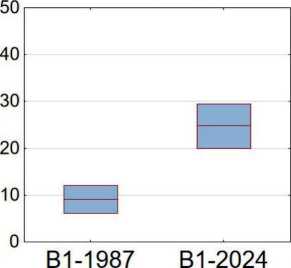
Fig. 2. Boxplots of variation in tree layer cover (A), undergrowth (B1) and understorey (B2) at different measurement periods. In the figure: mean values and 0.95 confidence interval крытие которой увеличилось впятеро (с 5.8 до 27.6%) и в отдельных сообществах доходило до 80–90%. Таким образом, на месте старовозрастных ельников во многих случаях сформировались лещинники, что характеризует другой тип вторичных сукцессий.
Оценка фитоценотических связей, рассчитанная на основе гамма-корреляции проективных покрытий основных лесообразующих видов двух временных срезов, продемонстрировала разную степень взаимозависимости видов. Сомкнутость ели в древостое 2024 г. имеет значимую отрицательную связь с показателями 1987 года (r=-0.344) (табл.), что может быть связано с распадом пре- имущественно более сомкнутых еловых древостоев. Связь подроста ели (r=0.388) и лещины (r=0,423) – положительная, что означает схожую картину их распространения в 1987 и 2024 гг. и лишь активизацию роста этих видов после распада древостоев. В целом в еловых сообществах достоверно сменилось распределение ели в составе древостоя, в то время как проективные покрытия сопутствующих видов за 37 лет остались положительно связанными.
Изменение экологических условий местообитаний за период с 1987 по 2024 гг. оценивалось с помощью экологических шкал Элленберга в пространстве ординационных осей на основе неме-
Гамма-корреляция проективного покрытия основных лесообразующих видов древесного и кустарникового ярусов в ельниках в разные годы исследований
Таблица
Table
Gamma-correlation of main forest-forming species of tree and shrub layers cover in spruce forests in different measurement periods
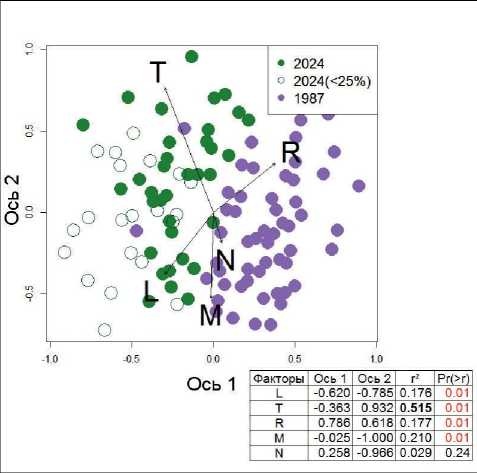
Рис. 3. NMDS ординация и корреляция распределения описаний с ординационными осями и квадраты коэффициентов корреляции (r2) Обозначение факторов: L – освещенность, Т – температура, R – кислотность почв, M – увлажнение почв, N – содержание азота в почве. Обозначение: <25% – проективное покрытие древостоя в описании менее 25% (распавшиеся ельники).
Fig. 3. NMDS ordination and correlation of the relevès distribution with ordination axes and squares of correlation coefficients (r2) Factors: L – lightness, T – temperature, R – soil reaction, M – soil moisture, N – soil nitrogen richness. Denotation: <25% – stand coverage is less than 25% (decayed spruce forests) in the relevès ров в точках описаний, повышение значений оценок по шкале температуры связано главным образом с выпадением ели (с более низким баллом по данному фактору) и увеличением обилия лещины и подроста клена (с более высокими баллами).
В работе были рассмотрены изменения структуры ценопопуляций видов деревьев и кустарников в малонарушенных старовозрастных ельниках за последние четыре десятилетия с учетом экологических условий местообитаний и межвидовых связей. В результате сравнения характеристик растений верхнего полога за два срока наблюдений выявились два разнонаправленных вида вторичных сукцессий, связанных с распадом древостоя. Во-первых, это демутационный процесс с восстановлением молодого поколения ели в окнах полога. Во-вторых, активное развитие лещины. Общий тренд неморализации состава еловых лесов связан как с внутренней динамикой сообществ, так и с влиянием изменений внешних (климатических) факторов.
Массовое усыхание темнохвойных лесов в регионе является следствием климатических изменений с особо катастрофическими последствиями периодически повторяющихся засух. Увеличение средней температуры на 2°С за 40 лет наблюдений является критичным для еловых лесов, находящихся на своей южной границе распространения. Наблюдения спонтанной динамики сообществ еловых лесов Центрально-Лесного заповедника (Тверская обл.) подтвердили сходную тенденцию к распаду в течение двух лет в результате повторяющихся метеорологических аномалий [3], дальнейшего ослабления деревьев и вспышек насекомых-вредителей.
Работа выполнена при поддержке гранта РНФ № 24-17-00120 «Леса Русской равнины: тренды последних десятилетий» по части формулировки проблемы и сбора полевых данных, а также ГЗ Института географии РАН (FMWS-2024-0007 No. 1021051703468-8) по обработке данных и анализу результатов.
