ДНК-генотипирование на основе SSR-Mapkepob - апробирование и подбор оптимальных условий для Glycine max (L.) Merr
 Merr")
Автор: Рамазанова С.А., Гучетль С.З., Антонова Т.С.
Статья в выпуске: 2 (137), 2007 года.
Бесплатный доступ
Оптимизирована методика выделения ДНК сои. Подобраны оптимальные условия амплификации ДНК и электрофоретического разделения продуктов ПЦР для 10 микросателлитных праймеров. Выявлен 31 аллельный вариант, пригодный для ДНК-генотипирования Glycine max (L.) Merr.
Короткий адрес: https://sciup.org/142150832
IDR: 142150832 | УДК: 633.853.52:34.23.37
DNA genotyping on the SSR Markers base - approbation and selection of optimal conditions
The technique of a soybean DNA extraction is optimized. Optimal conditions of DNA amplification and electrophoretic separation of PCR products for 10 microsatellite primers are selected. 31 allelic variant, suitable for DNA-genotyping of Glycine max (L.) Merr are revealed.
Текст научной статьи ДНК-генотипирование на основе SSR-Mapkepob - апробирование и подбор оптимальных условий для Glycine max (L.) Merr
Соя в России – динамично развивающаяся культура, площади под которой год от года систематически увеличиваются. Новые сорта этой культуры разного пищевого назначения, созданные во ВНИ-ИМК в последние годы, занимают на юге РФ свыше 90 % посевных площадей. Соя – самоопылитель, но, несмотря на это у нее богатый генетический потенциал, который трудно выявить традиционными методами. Ряд исследований отечественных и зарубежных ученых посвящен разработке маркерных систем на основе ПЦР для выявления генетического разнообразия и последующей идентификации дикорастущей ( G. soja L.) и культурной ( G. max L.) сои [1, 3]. Наиболее высокий уровень полиморфизма у сортов культурной сои выявлен по микросателлитным локусам [4, 6]. Для генетического материала из разных селекцентров аллельный полиморфизм одних и тех же локусов может быть неодинаков. С этих позиций отечественные сорта сои являются совершенно неизученными.
Целью данной работы было выявление полиморфных амплифицированных фрагментов ДНК сои для последующего создания системы молекулярных маркеров, пригодных в идентификации и оценке генетического разнообразия отечественных генотипов этой культуры.
Материалы и методы. Материалом исследований послужили 13 сортов сои селекции ВНИИМК (Лань, Лира, Фора, Валента, Дельта, Парма, Рента, Веста, Вилана, Лакта, Ника, Дива, РВБ), два иностранных сорта (Вильямс, Алдана) и один вид дикорастущей сои G. gracilis (К-4947). ДНК выделяли из зеленых трехлопастных листьев сои по методу Saghai-Maroof et аl. (1984), модифицированному нами. Навеску 0,2 г растительной ткани растирали с 0,9 мл СТАВ буфера, выдерживали в течение часа при температуре 65 оС. Добавляли ½ объема смеси хлороформа и изоамилового спирта (24:1), перемешивали, центрифугировали. Субнатант, перенеся в чистые пробирки, смешивали с равным объемом хлороформ-изоамиловой смеси. ДНК осаждали охлажденным до -20 оС изопропанолом. Подсушенную ДНК растворяли в 100-200 мкл воды в течение 1215 часов. Концентрацию ДНК в полученном препарате определяли по интенсивности свечения пробы объемом 10 мкл в ультрафиолетовом свете после 78
электрофореза в 1 % агарозном геле, содержащем бромистый этидий.
Для проведения полимеразной цепной реакции брали 25 мкл реакционной смеси следующего состава: 67 мМ трис-HCl, рН 8,8, 16,6 мM сульфата аммония; 1,5-3,0 мM MgCl2, 0,01 % Tween 20, по 0,2 мM дезоксирибонуклеозидфосфатов, по 10 пМ праймеров, 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК-полимеразы. Амплификацию проводили в приборе Терцик. Для реакций с SSR-праймерами сои использовали терморежим с начальной денатурацией при 96 оС в течение 2 мин, затем 32 цикла: денатурация при 94 оС – 30 сек, отжиг при 45-60 оС в течение 40 сек в зависимости от праймера, элонгация – 1 мин при 70 оС, финальная элонгация при 70 оС – 2 мин.
Электрофорез продуктов амплификации проводили в 2 % агарозном геле (ТАЕ-буфер) в камере для горизонтального электрофореза в течение 60-120 мин при силе тока 50-58 mA и напряжении 80-100 V. Документировали результаты электрофореза при помощи трансиллюминатора и видеосистемы с программным обеспечением Gel Imager-2.
Результаты. Выполнение ДНК-генотипирования требует проведения предварительных экспериментов по оптимизации методики выделения ДНК, подбору оптимальных температурных режимов амплификации, скринингу SSR-праймеров, выявляющих полиморфизм фракций ДНК, оптимизации физикохимических параметров электрофореза.
Выделение ДНК сои вначале проводили из 5-10дневных этиолированных проростков. Она была непригодна для амплификации. Пригодной оказалась ДНК, выделенная из зеленых трехлопастных листьев, без использования активированного угля.
Использовали 10 пар микросателлитных праймеров, описанных в работах зарубежных авторов [2, 4]. Для них вычисляли теоретическую температуру отжига. Теоретические значения температуры отжига являются приблизительными, и требуется их экспериментальная оптимизация. В таблице 1 показано сравнение теоретических и экспериментальных температур отжига для выбранных праймеров. Известно, что чем выше температура отжига, тем меньше вероятность неспецифичного связывания праймера с матричной ДНК, т. е. снижение температуры отжи-
Таблица 1 – Оптимизация температуры отжига для ПЦР анализа микросателлитных локусов
ДНК сои
|
Название локуса |
Температура отжига (ºС) |
Количество фракций |
||
|
расчетная |
экспериментальная |
продуцируемых |
полиморфных |
|
|
45 |
5 |
3 |
||
|
SATT 1 |
60 |
50 |
4 |
4 |
|
60* |
4 |
4 |
||
|
45 |
4 |
2 |
||
|
SATT 2 |
63 |
50 |
2 |
2 |
|
60* |
2 |
2 |
||
|
45 |
5 |
3 |
||
|
SATT 5 |
58 |
50 |
5 |
4 |
|
55* |
4 |
4 |
||
|
45 |
7 |
2 |
||
|
SOYPR 1 |
68 |
50 55 |
5 3 |
2 2 |
|
60* |
3 |
2 |
||
|
45 |
2 |
2 |
||
|
SOYGY |
66 |
50 |
2 |
2 |
|
2 |
55 |
2 |
2 |
|
|
60* |
2 |
2 |
||
|
45 |
5 |
2 |
||
|
SAT 1 |
70 |
50 55* |
4 6 |
2 4 |
|
60 |
6 |
4 |
||
|
50* |
7 |
2 |
||
|
SAT 43 |
64 |
55 |
5 |
2 |
|
60 |
4 |
2 |
||
|
SOYHSP 176 |
67 |
50 55 60* |
5 5 4 |
3 3 3 |
- оптимальная температура отжига для получения фракций ДНК с чёткой визуализацией
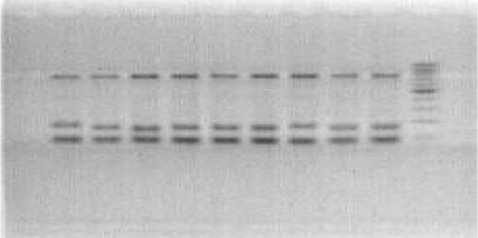
а

b
Рисунок 1 – Электрофоретические спектры продуктов амплификации ДНК сортов сои по локусу SAT5, при температурах отжига: а – 50 оС, b – 55 оC га ведет к гибридизации праймера с не полностью комплементарной ему последовательностью нуклеотидов. При этом могут появляться дополнительные, невоспроизводимые фракции ДНК. На рисунке 1 представлены продукты амплификации с праймером SAT5 при температурах отжига 50 и 55оС. Очевидно, что при температуре 55 оС (рис. 1b) в информативной зоне от 100 до 200 пар нуклеотидов фракции чётко различаются. А неинформативные фракции (700 п.н. и менее 100 п.н.) почти не видны.
Температура элонгации зависит от типа используемой ДНК-полимеразы и является постоянной от 70 до 72 оС. В программах для амплификации SSR локусов мы использовали температуру 70 оС.
Наши исследования показали, что изученные локусы являются полиаллельными (табл. 2). У локуса SATT9 выявлено наибольшее число аллелей 5. У трех локусов SATT1, SATT5, SAT1 обнаружено по 4 аллеля. Два локуса SATT36 и SOYHSP 176 имеют по 3 аллеля, и четыре локуса SATT2, SOYPR1, SOYGY2 и SAT43 – по 2 аллеля.
ПЦР-анализ 16 генотипов сои, изученных по этим локусам, позволил нам выявить 31 аллельный вариант – от 2 до 5 аллелей на локус, что в среднем составляет 3,1 аллель на один микросателлитный маркер.
Таким образом, все использованные для ПЦР пары праймеров выявляли полиморфизм фракций ам-плифицированной ДНК, что говорит об их пригодности для молекулярно-генетической характеристики сортов сои.
Время проведения электрофореза подбирали для каждого праймера индивидуально. Для этого электрофорез продуктов каждой реакции проводили дважды при разной продолжительности для того, чтобы обнаружить полиморфные фракции. Влияние продолжительности электрофореза на разделение продуктов реакции было разным в зависимости от используемого праймера. Например, праймер SOYHSP176 давал продукты амплификации, различающиеся небольшим числом пар нуклеотидов, поэтому при длительности электрофореза 90 мин различия между фракциями визуально не выявлялись (рис. 2a). Чтобы получить четкие различия между фракциями, время электрофореза увеличивали до 120 минут (рис. 2b).
Таблица 2 – Характеристика исследованных микросателлитных локусов сои
|
Локус |
Повтор |
Оптимальная температура отжига (оС) |
Количество фракций |
|
|
проду-цируе-мых |
полиморфных |
|||
|
SATT 1 |
(ATT)24 |
60 |
4 |
4 |
|
SATT 2 |
(AAT)18 |
60 |
2 |
2 |
|
SATT 5 |
(TAA)21 |
55 |
4 |
4 |
|
SATT 9 |
(AAT)12 |
45 |
5 |
5 |
|
SOYPR 1 |
(TAT)20 |
60 |
3 |
2 |
|
SOYGY 2 |
(AT)9(ATT)6 |
55 |
2 |
2 |
|
SAT 1 |
(AT)17 |
60 |
6 |
4 |
|
SAT 36 |
(AT)19 |
55 |
3 |
3 |
|
SAT43 |
(AT)20 |
55 |
5 |
2 |
|
SOYHSP176 |
(AT)15 |
60 |
4 |
3 |
а
b
Рисунок 2 – Электрофоретические спектры продуктов амплификации ДНК сортов сои по локусу SOYHSP176 при продолжительности электрофореза: а – 90 мин; b – 120 мин.
Праймеры SAT43, SAT36, SATT1, SATT2 давали продукты реакции, визуально легко различимые при длительности электрофореза 90 мин. Было также установлено, что продолжительность электрофореза более 120 мин, при силе тока 55 ma и напряжении 70-80 V не выявляла дополнительно полиморфных фрагментов ДНК.
Таким образом, в результате выполненной работы оптимизирована методика выделения ДНК сои. Подобраны оптимальные условия амплификации ДНК и продуктов ПЦР для 10 микросателлитных прайме- ров. В локусах ДНК изученных генотипов сои выявлен 31 аллельный вариант, пригодный для генотипирования.
Исследования выполнены при финансовой поддержке РФФИ и Администрации Краснодарского края, грант № 06-04-96757.
