Двустороннее распространение нервных импульсов, открытое гистологом А.И. Бабухиным - основа ретикулярной теории

Автор: Сотников Олег Семенович, Кокурина Татьяна Николаевна, Подольская Людмила Абрамовна
Журнал: Морфологические ведомости @morpholetter
Рубрика: Обзорные и общетеоретические статьи
Статья в выпуске: 2 т.25, 2017 года.
Бесплатный доступ
Ретикулярная теория организации нервной системы предполагает электрическую и тинкториальную взаимосвязь нейронов с помощью синцитиальных мостиков в сетевых образованиях мозга и ганглиев. Это означает, что импульсы распространяются во все стороны по путям нервных ветвей. Нейронная же теория предполагает полную цитоплазматическую изоляцию смежных нейронов. Основное положение нейронной доктрины предусматривает последовательный переход нервного импульса в одном направлении от одного нейрона к другому по рефлекторной дуге. Этот закон динамической поляризации сформулирован одновременно в 1891 г. двумя великими нейрогистологами Рамон-и-Кахалем и Ван Гехухтеном. В дальнейшем этот закон был, как казалось, блестяще подтвержден открытием медиаторных синапсов с помощью электронной микроскопии. Электронная микроскопия химических синапсов позволила считать нейронную теорию Рамон-и-Кахаля единственно верной и абсолютно доказанной. Нейронная теория уверенно вошла во все учебники и руководства по неврологии, а теория ретикуляризма была признана ошибочной и неверной...
Двунаправленность нервного импульса, щелевые контакты, электрические синапсы, синцитиальные перфорации
Короткий адрес: https://sciup.org/143177189
IDR: 143177189 | DOI: 10.20340/mv-mn.17(25).02.01
The two-way distribution of nervous pulses open by hystologist AI Babukhin - the basis of the reticular theory
Reticular theory of the nervous system presupposes the electrical and tinctorial interrelation of neurons in the nervous system with the help of syncytial bridges in the brain and ganglion network formations. This means that the impulses spread in all directions along the path of the nervous branches. The neuronal theory presupposes complete cytoplasmic isolation of adjacent neurons. The basic position of the neural doctrine provides for a successive transition of the nerve impulse in one direction from one neuron to another, along a reflex arc. This «law of dynamic polarization» was formulated simultaneously in 1891 by two great neurohistologists Ramon y Cajal and Van Gehuchten. Later, this law seemed to be brilliantly confirmed by the discovery of mediator synapses using electron microscopy. Electron microscopy of chemical synapses allowed us to consider Cajal's neuronal theory as the only true and absolutely proven one. Neuronal theory confidently entered in the all textbooks and neurology manuals, and the theory of reticularism was recognized as erroneous and incorrect...
Текст научной статьи Двустороннее распространение нервных импульсов, открытое гистологом А.И. Бабухиным - основа ретикулярной теории
Сотников О.С., Кокурина Т.Н., Подольская Л.А. Двустороннее распространение нервных импульсов, открытое гистологом А.И. Бабухиным – основа ретикулярной теории// Морфологические ведомости.- 2017.- Том 25.- № 2.- С. 8-13. (25).02.01
Sotnikov OS, Kokurina TN, Podolskaya LA. The two-way distribution of nervous pulses open by hystologist AI Babukhin - the basis of reticular theory. Morfologicheskie Vedomosti – Morphological Newsletter. 2017 Jun 30;25(2):8-13. (25).02.01
Введение . Основные положения ретикулярной теории организации нервной системы предполагают широкую взаимосвязь нейронов с помощью нервных волокон, объединённых в форме сетей. Принципиальная особенность этой теории состоит в том, что волокна нервной сети считаются связанными между собой как электрическими, так и цитоплазматическими синцитиальными взаимосвязями. Предполагается, что нервные импульсы могут распространяться во все стороны по направлению нервных ветвей. Другие важные постулаты этой теории являются производными. Противоположная, или нейронная доктрина, отстаивает полную цитоплазматическую изоляцию нейронов и строгую последовательность перемещения нервного импульса от дендритов к нейросоме, а затем к мышце по рефлекторной дуге. Этот закон динамической поляризации нейрона был одновременно сформулирован испанским нейроморфологом Рамон-и-Кахалем [1] и французским гистологом Ван Гехухтеном (1891) [2]. Он стал основой физиологических доказательств нейронистов. Эта незыблемая идея прочно закрепилась в руководствах нашего времени. Казалось, что наконец морфологические и физиологические идеи обрели единую основу, хотя у известных морфологов – ретикуляристов [3], да и нейронистов [4], было достаточно доказательств, демонстрирующих синцитиальные, то есть цитоплазматические, взаимосвязи нейронов. Главным препятствием в разрешении дискуссии было несовершенство методов морфологического исследования.
В начале семидесятых годов XIX-го столетия русский гистолог профессор А.И. Бабухин решил исследовать загадочное биологическое явление – генерацию электричества живыми тканями и органами некоторых пресноводных рыб. В этой статье путешествие Александра Ивановича Бабухина вдоль берегов Нила [6] авторам можно было бы описать как увлекательное «Путешествие натуралиста...», подобно Чарльзу Дарвину [5]. А.И. Бабухину, как микроскописту, удалось подробно изучить историю развития электрических органов животных, доказав их происхождение из мышечной, а не из нервной ткани; описать развитие, строение и сродство физиологии электрических и псевдоэлектрических органов, внести значительный вклад в дискуссию Ч. Дарвина об эволюции биоэлектрических органов. Увлечённый электрофизиологическими опытами Э.Г. Дюбуа-Реймона, не имея никакой электрической аппаратуры, не найдя у местных жителей ни кусочка металлического провода для электродов, буквально голыми руками и скальпелем он сделал важнейшее для всей нейрофизиологии открытие на органах электрического сома Malopterurus electricus – открыл явление двунаправленного движения нервного импульса. Известный физиолог Н.Е. Введенский высоко оценил находки автора [7]. А.И. Бабухин, разделив электрический орган на две изолированные части, имеющие множество нервных волокон, отходящих от единого толстого отростка гигантского нейрона, перерезал часть этих ветвей и раздражал их (рис. 1). Наблюдая, как местные рыбаки по неосторожности получают настоящий электрический шок при прикосновении к телу электрического сома, он решил использовать часть органа в качестве электро-стимулятора, накладывая его на центральные культи перерезанных нервов. Центральная часть органа удалялась, так, что две оставшиеся половины были полностью изолированы. При этом в опытах он безотказно получал ответ интактной части электрического органа. Открытие состояло в том, что импульсы от раздраженных центральных нервных культей вначале распространялись ретроградно, в направлении к телу нейрона (рис. 1), и только затем переходили в нисходящие соседние неперерезанные волокна, то есть в антероградном направлении. Следовательно, ток мог перемещаться в двух противоположных направлениях. В зависимости от условий опытов направление «животного» тока можно было изменять на противоположное. Опыты Бабухина, таким образом, противоречили главному закону нейронной доктрины - закону динамической поляризации нейронов. Это было уникальное открытие нашего соотечественника.
Доказательство способности нервных импульсов распространяться по волокнам в разные стороны свидетельствовало об их возможности перемещаться по нервным сетям, что и предполагали ретикуляристы. Строго говоря, это открытие противоречило нейронной доктрине в принципе. Подобные данные были подтверждены затем в опытах S.N. Langley и H.K. Anderson [8]. Большую поддержку выводам Бабухина оказали многочисленные работы C.S. Sherrington [9]. Сразу же после опытов Бабухина он опубликовал работу «Двойное (антидромное) проведение в центральной нервной системе», в которой цитировал его работы и показал естественную антидромную проводимость вдоль восходящих путей спинного мозга. Его с соавторами знаменитый труд «Рефлекторная деятельность спинного мозга» был опубликован с предисловием академика И.П. Павлова [10]. В этой работе авторами антидромное раздражение, открытое Бабухиным, использовано как механизм изучения ослабления центрального возбуждения и образования центрального возбуждения в синапсах (цит. по [10], с. 44). На основании открытия Бабухина был сформулирован также закон двустороннего проведения [11].
Важно подчеркнуть, что основоположник нейронной теории Рамон-и-Кахаль, хорошо знавший Шеррингтона, никак не отреагировал ни на открытия А.И. Бабухина, ни на их множественные доказательства Ч.С. Шеррингтоном с соавторами. Камилло Гольджи, как глава сторонников ретикулярной теории, также не интересовался физиологией нейронов. Приведенные данные свидетельствуют о том, что наш соотечественник, гистолог, первооткрыватель антидромного распространения нервного тока А.И. Бабухин, вместе с Ч.С. Шеррингтоном должны считаться основоположниками экспериментального доказательства ретикулярной теории. К сожалению, во многих современных руководствах это открытие недостаточно оценено как важное основание ретикулярной теории. В многочисленных длительных дискуссиях сторонников нейронной и ретикулярной теорий нередко возникал фундаментальный вопрос о том, каким образом в естественных условиях нервный импульс может менять направление своего движения в нервной сети при переходе из одного нервного волокна к другому. В этом и состоит смысл дальнейшего продолжения исследований по ретикулярной теории, в том числе и цель настоящего исследования.
Ретикулярная теория предполагала, что между нейронами в нервных сплетениях, помимо электрической, существует синцитиальная цитоплазматическая связь. Синцитиальные цитоплазматические мостики описаны у клеток многих тканей, так что наличие этих структур в нервной системе можно считать вполне закономерным явлением, тем более что синцитии нейронов были неоднократно обнаружены и в до электронно-микроскопический период [4]. К сожалению, такие находки при световой микроскопии редки, неизвестны причины и закономерности их появления. Так что в качестве концепции ретикуляризм ранее трудно было обосновать и представления о ретикулярной гипотезе постепенно исчезли из руководств и учебников. Однако память о таком удивительном явлении, как синцитиальное слияние нейронов, осталась у неврологов, тем более что, согласно клеточной теории Шванна, нейроны не должны быть исключением. В связи с вышеуказанным, мы провели экспериментальное электронно-микроскопическое исследование сплетений вегетативной нервной системы.
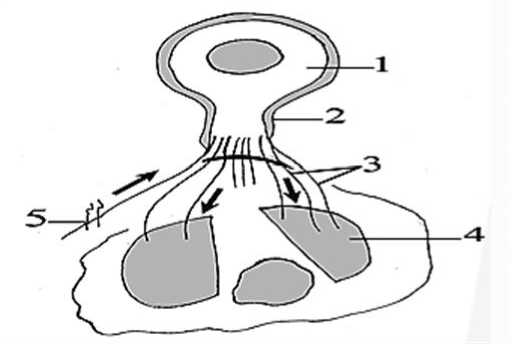
Рис. 1. Схематическое изображение опытов А.И. Бабухина с изменением направления импульсов нервного тока волокон электрического органа (схема). 1 – гигантский нейрон электрического сома; 2 – крупный аксон; 3 – многочисленные тонкие эфферентные ветви аксона; 4 – ампутированные фрагменты электрического органа; 5 – электрический стимулятор в виде фрагмента электрического органа, помещенного в область одного из перерезанных нервных отростков; стрелки – целлюло-петальное и целлюло-фугальное направления распространения электрических импульсов.
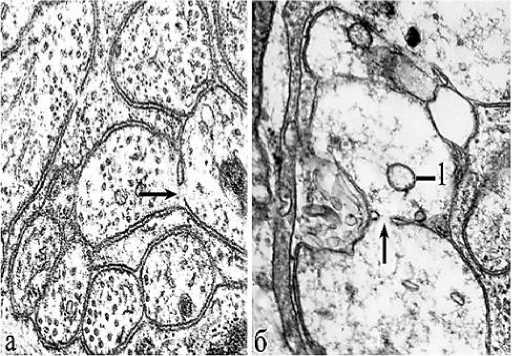
Рис. 2. Электронное микрофото. Несколько синцитиальных перфораций в области нервного сплетения; а – общий вид препарата; б – синцитиальная пора между двумя волокнами, резко различающихся по диаметру; в – эффект разрыхления контуров двух контактирующих мембран без образования чётко выделенной поры; г –синцитиальная связь препарата на краю сплетения и коллагеновых волокон; 1 – шванновская клетка; 2 – коллаген; стрелки – синцитиальные связи. Ув.: х10000.
Материалы и методы исследования . Исследования проведены на 5-ти половозрелых интактных крысах Wistar и 10 крысах Wistar, испытавших гипоксию в течение 50 минут. Для выведения животных из опыта использовали этиловый эфирный наркоз. У животных извлекали и фиксировали сплетения каудального брыжеечного ганглия. Все манипуляции с животными производились согласно этическим принципам, изложенным в Европейской конвенции по защите позвоночных животных, используемых в экспериментах. Кровеносную систему животного перфузировали сначала раствором Рингера, а затем 2,5% раствором глутарового альдегида, приготовленного на 0,1 М фосфатном буфере (рН 7,2―7,4). Извлечённый препарат делили на части и фиксировали в охлажденном 2,5% растворе глутарового альдегида в течение 1,5 часов, затем в 1% растворе четырехокиси осмия в течение 2 часов при 4° С. Далее кусочки ганглия обезвоживали в 70%, 96% (дважды) и 100% этаноле (три смены по 20 мин). После этого материал заливали в аралдит и помещали в термостат при 37°С, а затем при 56°С. Срезы, приготовленные на ультратоме Leica EM UС7 (Германия), контрастировали цитратом свинца и уранил-ацетатом по Рейнолдсу. В работе использован микроскоп Jeol JEM-1400 (Jeol, Япония).
Результаты исследования и обсуждение . Синцитиальные перфорации между нейритами сплетения действительно встречаются редко и имеют различную форму. Естественные синцитиальные прободения спаренных мембран располагаются как между волокнами одного размера, так и между волокнами, резко различающимися по размеру (рис. 2, «б»). Перфорации располагаются на волокнах внутри мелких пучков или у их краев на границе с коллагеновыми волокнами (рис. 2-г). У крупных сплетений может встречаться несколько синцитиев и одиночные шванновские ядра (рис. 2-а). Все нейриты покрыты глиальной цитоплазмой, но не имеют выраженных мезаксонов. Синцитии чаще имеют субмикроскопические параметры, но могут принимать и размеры микроскопических пор (рис. 3), причём они не отличаются друг от друга, даже если на одном из препаратов отмечены признаки реактивного набухания при гипоксии (рис. 3-б). Синцитиально соединёнными могут быть как обычные нейриты, так и синапсы (рис. 4). По нашим представлениям, перфорации не имеют строгой топографической локализации и встречаются как в области сплетений отростков, так и при контакте тел нейронов.
Хотя исследования показывают, что синцитии в нервных сплетениях встречается редко, но всё-таки этого достаточно, чтобы рассматривать их в качестве морфологического феномена, обладающего особенными функциями. Теперь ясно, что закон двустороннего проведения, обнаруженный А.И. Бабухиным, может реализоваться не только в одном волокне. Импульсы способны передаваться с помощью синцитиев и на многих смежных волокнах, причём не способом эфапса, а как прямая электрическая немедиаторная проводимость. Гипотеза Камилло Гольджи подтверждается. Таким образом, ретикулярная теория имеет такие же основания, как и нейронная теория, и нуждается в дальнейшей разработке. Это тем более целесообразно, что в последнее время всё более широкое распространение получают исследования электрических синапсов, которые также способны проводить электрические импульсы в противоположные стороны и передавать в области контакта двух мембран низкомолекулярные вещества (например, многие красители). Такие электрические синапсы получили название щелевых контактов (рис. 5). По своим свойствам они занимают промежуточное положение между прилежащими нейролеммами и контактами с суженной межклеточной щелью, обладающей электрической и тинкториальной проницаемостью. Сравнивая различные щелевые контакты при гипоксии, удается установить, что величина их щели может резко сужаться вплоть до полного исчезновения, то есть слияния прилежащих мембран.
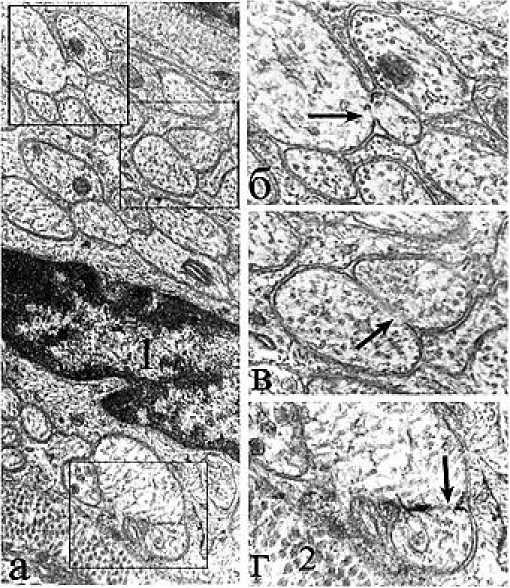
Рис. 3. Электронное микрофото. Синцитиальные поры нервных волокон в области сплетения; а - синцитий двух интактных волокон; б - синцитиальная перфорация двух реактивно изменённых нервных волокон (гипоксия); 1 - вакуолеподобная остаточная структура; стрелки - поры синцитиума. Ув.: в -х25000, г - х15000.
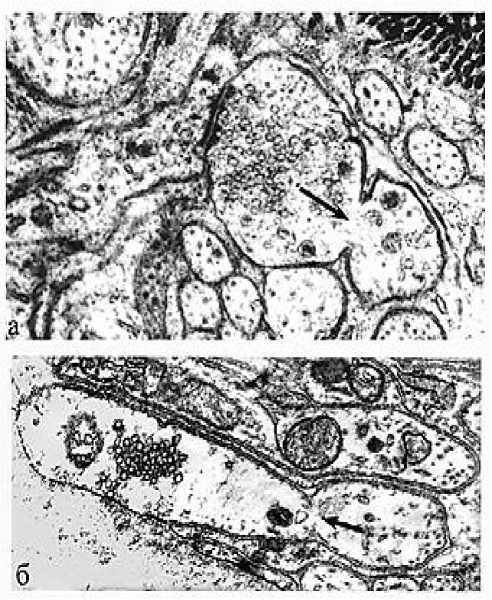
Рис. 4. Электронное микрофото. Синцитиальные перфорации, образованные в области химических синапсов; а - интактный препарат; б - агрегация синаптических пузырьков в области реактивно измененного синапса (гипоксия); стрелки – синцитиальные мембранные перфорации. Ув.: а – х30000, б - х10000.
В области слившихся наружных слоев мембран их толщина уменьшается более чем в два раза и представляет собой тонкую электронно-плотную перемычку (рис. 5-б). Этот локальный процесс, видимо, начинается с заполнения прозрачной обводненной щели концентрирующейся электронно-плотной водорастворимой субстанцией (рис. 5-а). Как известно, любые мембраны формируются и сохраняются только благодаря гидрофобно-гидрофильному взаимодействию полярных фосфолипидов. Поэтому уменьшение объёма щели контакта и воды нарушает связи гидрофильных фрагментов контактирующих мембран и других компонентов фосфолипидов, обычно организующих их билипидность (рис. 6). Естественно, такие плотно контактирующие мембраны должны обладать склонностью к механической неустойчивости и разрушению (рис. 6-б,в). Очевидно, именно этим объясняется появление синцитиальных пор и перфораций мембран именно в области щелевых контактов. На препаратах появляются частично или полностью прерванные структуры, которые чётко отражают все стадии превращения щелевых контактов в синцитии (рис. 6). Сужающиеся межмембранные щели контакта истончаются, а наружные слои сливающихся мембран становятся предельно электронно-плотными, изогнутыми терминалями-тяжами (рис. 7-а,в) или изолированными вакуолеподобными фосфолипидными липосомами (рис. 6-в; рис. 7-а,б). Прерванные контактирующие двуслойные нейролеммы способны повторно сливаться, запечатывая межклеточную перфорацию (рис. 7-а).
Заключение. Перечисленные нами данные позволяют обобщить исследования, полученные морфологами-ретикуляристами 19-20 века (Бабухиным, Шеррингтоном), и современные морфо-физиологические представления об электрических синапсах и синцитиальных перфорациях. Результаты этих исследований можно представить в качестве единого мембранного процесса, общего для всех клеток (рис. 8). Сформулированная гипотеза позволяет представить такие статичные морфологические структуры, как щелевые контакты и синцитиальные перфорации, в качестве кинетических механизмов единого нового явления в нейроморфологии и нейропатологии. В подтверждение возможности превращения щелевых контактов в синцитиальные поры можно привести множество дополнительных фактов, предполагающих появление этого феномена при альтерации нейронов.
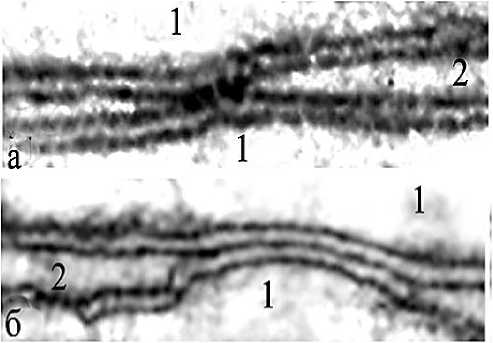
Рис. 5. Электронное микрофото. Щелевые контакты (электрические синапсы) в области спаренных волокон нервных сплетений; а – сужение межклеточной щели, заполненной электронно-плотным материалом; б – слияние наружных поверхностей контактирующих мембран, исчезновение межмембранной щели (перемычка); 1 – аксоплазмы контактирующих волокон; 2 – межмембранные щели. Ув.: х30 000.
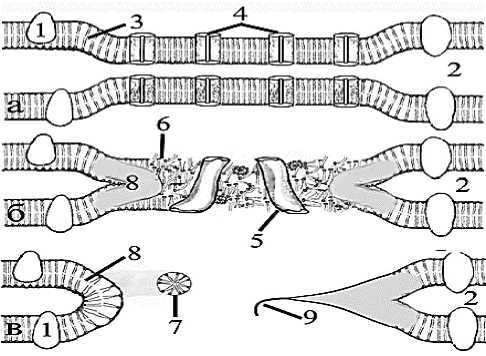
Рис. 6. Схема механизма превращения щелевого контакта в синцитиальную перфорацию; а – молекулярная организац щелевого контакта с сужением межклеточной щели; б нарушение геометрической молекулярной организац билипидных слоёв мембран при их сближении и слиянии; в формирование синцитиальной перфорации; 1 – молекул интегральных, мембранных белков; 2 – межмембранная щел 3 – билипидная организация мембраны; 4 – молекул коннексиновых полуканалов; 5 – конформационные изменен белков и расширение коннексиновых пор; 6 – нарушен упорядоченности организации фосфолипидных молекул; 7 остаточные фосфолипидные тельца (липосомы); 8 повторное слияние фосфолипидных спаренных мембра закупоривающих межклеточную щель; 9 – прерванн электронно-плотная терминаль щелевого контакта.
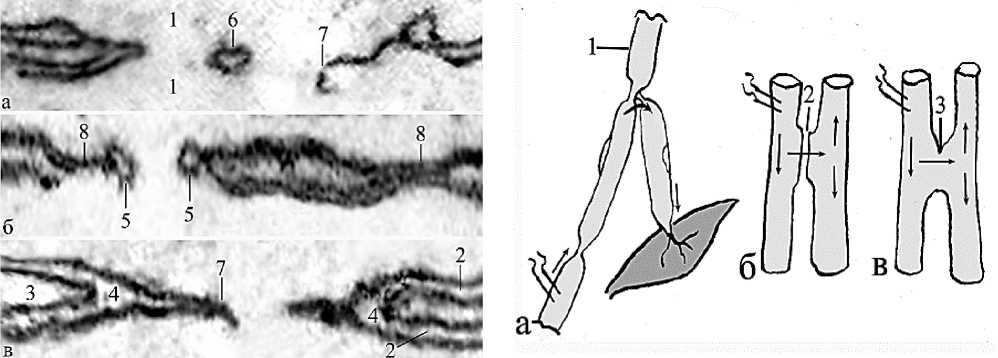
Рис. 7. Электронное микрофото. Варианты прерванных терминалей синцитиальных перфораций; а – изолированное остаточное тельце и остаток перемычки; б – формирование щелевых контактов, перфорации и отпочковывание остаточных фосфолипидных телец; в – истончение, разрыв щелевого контакта и запечатывание межклеточного пространства; 1 – цитоплазмы спаренных нейронов; 2 – мембраны смежных волокон; 3 – остатки межклеточных щелей; 4 – слияние мембран; 5 – формирующиеся остаточные тельца; 6 – изолированное остаточное тельце; 7 – остаток уплотненной перемычки. Ув.: х40 000.
Рис. 8. Варианты искусственных и естественных каналов двунаправленных импульсов нервных волокон; а – антидромная импульсация тока при искусственной ампутации и раздражении бифуркации миелинового нервного волокна; б – изменение направления электрических импульсов в области щелевого контакта; в – бидирекциональное направление импульсов в области синцитиальной перфорации; 1 – миелиновое нервное волокно; 2 – щелевой контакт; 3 – синцитиальный анастомоз между нервными волокнами; 4 – мышца.
Открытие электрических синапсов позволяет пересмотреть все важнейшие положения нейронной доктрины Рамон-и-Кахаля [14] об электрической, метаболической и механической изолированности нейронов, о распространении импульсов в одном направлении и невозможности их синцитиальной связи. Щелевые контакты доказывают реальность электрической взаимосвязи нейронов без участия медиаторных синапсов, возможность распространения импульсов во все стороны и частичный метаболический обмен между нервными клетками [15]. Ретикулярная связь огромного оркестра электрически взаимодействующих сетей нейропиля мозга способна влиять на когнитивные функции и на поведенческие перемены всего организма [16]. Щелевые контакты обладают комплексом неизвестных ранее физиологических механизмов. Они способны либо восстанавливать ритмическую частоту пачек спайков, либо обеспечивать коллективное их срабатывание с увеличением амплитуды потенциала действия нескольких нейронов [17], либо выступать в качестве фильтра низких частот [18].
Таким образом, исследования профессора А.И. Бабухина о возможности двунаправленного движения нервных импульсов стали одной из основ для доказательства возможности распространения импульса между волокнами в нервном сплетении и подтвердили морфологические исследования сторонников ретикулярной теории Камилло Гольджи о межнейрональных синцитиях; обнаружение электрических синапсов открыли новые представления об организации и функционировании всей нервной системы.
Список литературы Двустороннее распространение нервных импульсов, открытое гистологом А.И. Бабухиным - основа ретикулярной теории
- Ramon y Cajal S. Significacion fisiologica de las expansiones protoplàsmicas y nerviosas de la sustancia gris. Rev. Scienc. Med. Barcel. 1891;22:1-15.
- Van Gebuchten. La Structure des Centres Nerveux. Louvain. 1891. 193pp.
- Dogiel AS. Zur Frage über den Bau der Nervenzellen und über das Verhältniss ihres Axencylinder-(Nerven-) Fortsatzes zu den Protoplasmafortsützen (Dendriten). Arh. Mikrosk. 1893;41:62-87.
- Retzius G. Biologische untersuchunden. Neue Folge. Jena: Vong Fischer; 1894. Bd. VI. 325pp.
- Darvin Ch.R. Puteshestvie naturalista vokrug sveta na korable "Bigl"'. M., L.: Izd. CK VLKSM; 1936. 399s.
- Babuhin A.I. Jelektricheskie organy u ryb. M.: Retinoidy; 2007. 87s.
- Vvedenskij N.E. Polnoe sobranie sochinenij. L.: Izd. Len. gos. univer. im. A.A. Zhdanova; 1954. T. 5. 380s.
- Langley JN, Anderson HK. On Reflex Action from Sympathetic Ganglia. J. Physiol. 1894;6(5-6):410-440.
- Sherrington CS. Doubl (antidrome) conduction in the central nervous system. Lond. Proc. R. Soc. 1897;61:243-246.
- Krid R., Denni-Broun D., Ikkls I., Liddell E., Sherrington Ch. Reflektornaja dejatel'nost' spinnogo mozga. M.-L.: Gos. izd. biol. i med. lit., 1935. 268s.
- Bykov K.M., Vladimirov G.E., Delov V.E., Konradi G.P., Slonim A.D. Uchebnik fiziologii. M.: Medgiz, 1954. 891s.
- Buzsáki G. Electrical wirng of the oscillating brain. Neuron. 2001;31(3):342-344.
- Draguhn A, Traub RD, Bibbig A, Schmitz D. Ripple (approximately 200-Hz) oscillations in temporal structures. J. Clin. Neurophysiol. 2000;17(4):361-376.
- Mazzarello P. Camillo Golgi's scientific biography. J. Histol. Neurosci. 1999;8(2):121-131. (Dalhousie Univerity. 2012).
- Jin NG, Ribelayga CP. Direct Evidence for Daily Plasticity of Electrical Coupling Between Rod Photoreceptors in the Mammalian Retina. J. Neurosci. 2016;36(1):178-184.
- DOI: 10.1523/JNEUROSCI.3301-15.2016
- Okun M, Steinmetz NA, Cossell L, lacaruso MF, Ko H, Barthó P, Moore T, Hofer SB, Mrsic-Flogel TD, Carandini M, Harris KD. Diverse coupling of neurons to populations in sensory cortex. Nature. 2015;521(7553):511-515.
- DOI: 10.1038/nature14273
- Rosa EJr, Skilling QM, Stein W. Effects of reciprocal inhibitory coupling in model neurons. Biosystems. 2015;127:73-83.
- DOI: 10.1016/j.biosystems.2014.11.002
- Dargaei Z, Colmers PL, Hodgson HM, Magoski NS. Electrical coupling between Aplysia bag cell neurons: characterization and role in synchronous firing. J. Neurophysiol. 2014;112(11):2680-2696.
- DOI: 10.1152/jn.00494.2014
