Фауна крупных млекопитающих из плейстоценовых отложений Чагырской пещеры (Северо-Западный Алтай) по материалам раскопок 2007-2011 годов
 по материалам раскопок 2007-2011 годов")
Автор: Васильев С.К.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Сибирячихинский вариант среднего палеолита Атая
Статья в выпуске: 1 (53), 2013 года.
Бесплатный доступ
В статье проанализированы результаты исследования более 100 тыс. костных остатков из Чагырской пещеры (раскопки 2007-2011 гг.). Установлено, что тафоценоз слоев 5 и 6а сформировался преимущественно за счет пищевой активности волков и пещерных гиен, использовавших пещеру как убежище или логово для выведения потомства. Напротив, костные остатки из слоев 6б и 6в в приустьевой части пещеры накапливались в основном в результате охотничьей деятельности палеолитического человека, который специализировался в охоте на бизонов (Bison priscus). В меньшей степени его добычей становились лошадь Оводова (Equus (Sussemionus) ovodovi), северный олень (Rangifer tarandus), сибирский горный козел (Capra sibirica) и архар (Ovis ammon). Остатки фоновых видов мегафауны позволяют реконструировать существование степных ландшафтов в период формирования слоя 6. Накопление отложений слоя 5 происходило, по-видимому, на фоне значительной аридизации климата. По долинам рек и в горных ущельях на протяжении всего периода осадконакопления сохранялись небольшие участки лесных массивов.
Чагырская пещера, костные остатки, тафоценоз, хищники, палеолитический человек
Короткий адрес: https://sciup.org/14522965
IDR: 14522965 | УДК: 569
Текст научной статьи Фауна крупных млекопитающих из плейстоценовых отложений Чагырской пещеры (Северо-Западный Алтай) по материалам раскопок 2007-2011 годов
Чагырская пещера расположена в среднегорном районе Северо-Западного Алтая, на участке долины р. Чарыш, дренирующей отроги северного склона Тигирекского хребта [Деревянко, Маркин, Зыкин, 2008; Деревянко и др., 2009; Маркин, Зыкин, Зыкина, 2011]. В разрезе пещеры представлены голоценовые (слои 1–4) и плейстоценовые образования, верхняя часть которых сложена субаэральными осадками двух горизонтов (слой 5 и слои 6а, 6б, 6в/1, 2) лессовидных отложений. Нижний интервал последних содержит богатейшие археологические, палеонтологические и антропологические остатки. Анализ археологического материала позволил определить единственный его аналог на Алтае – технокомплексы пещеры Окладникова [Деревянко, Маркин, 1992]. Изучение индустрий этих двух пещер дало основания для выделения нового варианта среднего палеолита региона – сиби-рячихинского (или сибирячихинской линии развития), сопоставимого с мустье Закавказья, Юго-Западной Европы и Передней Азии [Деревянко, Маркин, 2012; Derevianko, Markin, 2011]. Судя по обнаруженным в пещерах антропологическим материалам, носителями данных традиций являлись представители неандертальского антропологического типа [Viola, Markin, Zenin et al., 2011; Viola, Markin, Buzhilova et al., 2012].
Изучение костных остатков крупных млекопитающих из Чагырской пещеры началось с первого года раскопочных работ [Васильев, 2009]. За пять сезонов раскопок (2007–2011 гг.) получено немногим более 100 тыс. остатков. Из них определимые до вида, рода или же класса составляют лишь 3,4 %. Весь имеющийся костный материал чрезвычайно фрагментарен. Обломков костей крупнее 5 см всего 5,5 %. Кроме того,
Археология, этнография и антропология Евразии 1 (53) 2013
обломки диафизов трубчатых костей из плейстоценовых слоев при высыхании расслаиваются и распадаются на отдельные части, в связи с чем число фрагментов костей в исследованной коллекции несколько превышает их изначальное количество. В голоценовых слоях 1–4 зафиксировано как минимум 20 таксонов, из крупных млекопитающих – бобр, волк, бурый медведь, косуля, марал, домашняя овца, лошадь и корова. В плейстоценовых отложениях (слои 5–7) найдены остатки млекопитающих 35 видов, а также кости рыб, птиц и человека (табл. 1). Среди определимых остатков крупных млекопитающих из слоев 5 и 6 абсолютно преобладают изолированные зубы и мелкие кости дистальных отделов конечностей (табл. 2).
Тафономические особенности
Ко стный материал в слоях частично переотложен в результате норной деятельности грызунов. Четко установить границы поноров в большинстве случаев не представляется возможным. При заполнении поно-ров грунтом единичные фрагменты костей голоценовой сохранности проникали до уровня слоя 7. В свою очередь, плейстоценовые костные остатки с норными выбросами попадали в голоценовые слои. Так, если в 4-м слое почти все фрагменты костей имеют типично голоценовую сохранность, то в 3-м ок. 1/4 костного материала явно происходит из плейстоценовых горизонтов. В слое 5 преобладают обломки костей плейстоценовой сохранности, но встречаются и типично голоценовые.
По отдельным слоям степень фрагментарности костных остатков изменяется незначительно (табл. 3). Наиболее мелкофрагментированный материал содержится в слое 5. Вниз по разрезу степень его фрагментарности последовательно снижается. В слоях 6а и 6б увеличивается доля более крупных обломков, в слое 6в в 2 раза возрастает количество фрагментов больше 5 см, что может свидетельствовать об общем увеличении скорости седиментации. В свою очередь, это является косвенным подтверждением более интенсивного использования пещеры человеком и крупными хищниками в данный период. В результате их активной жизнедеятельности в пещерные отложения в большом количестве могли попадать не только органические остатки, но и грязь, наносимая извне на подошвах лап и ног.
В слое 5 (горизонты 1–6) преобладают кости со следами кислотной коррозии. Они сильно истончены, ажурны, в ряде случаев со множеством мелких сквозных отверстий. Поверхность обломков костей заполирована и мелковолниста. Эмаль зубов копытных и хищников из отрыжек гиен заметно истончена либо полностью отсутствует. Их блестящая поверх- ность производит впечатление сильной окатаннос-ти. В слое 6а (горизонты 1–3) большая часть костей также разъедена кислотной коррозией. Наиболее крупный фрагмент трубчатой ко сти подобной сохранности (слой 6в/1, горизонт 1) достигает в длину 123 мм и имеет при этом сквозное отверстие диаметром 21–28 мм. В слое 6б/3 костных остатков, разъеденных кислотной коррозией, становится меньше, чем в слое 6а, но появляются кости с признаками искусственного раскола и следами порезов. В приустьевой части пещеры (раскопки 2008 г.) в слоях 6а (горизонты 1, 2) и 6б/1 на отдельных костях отмечены слабо выраженные извилистые линии, оставленные корневой системой растений.
Всего несколько мелких (1–2 см) фрагментов костей со следами воздействия огня зафиксировано в слоях 6а, 6б/4, и четыре происходят из материалов промывки осыпи.
Следы порезов каменными орудиями отмечены главным образом на костях из слоя 6в/1 (горизонты 1–5). При разборе коллекции они обнаружены на 13 обломках в основном размером 5–10 см, в т.ч. идентифицированных: диафиза берцовой кости и тонкой части лопатки, ребер и крупных трубчатых костей бизона. В ряде случаев наблюдаются множественные порезы, параллельные и V-образные. На небольшом фрагменте дистального отдела ребра бизона зафиксировано 9 параллельных и пересекающихся неглубоких нарезок, на другом обломке ребра – 14, на фрагменте трубчатой кости – 7.
Анализ остеологического материала показал, что раскопки трех последних сезонов (2009–2011 гг.) проводились в основном за пределами крупного скопления костных остатков в приустьевой части пещеры, оставленного палеолитическим человеком. Характерные осколки трубчатых костей, расколотых в свежем состоянии рукой человека, попадаются и в глубине пещеры, но уже в значительно меньшем количестве. С места разделки и поедания частей туш бизонов в приустьевой части обломки костей рассеивались во все стороны в виде сходящего на нет шлейфа. В глубине пещеры основную часть костных остатков составляют фрагменты костей и зубов, не имеющие отношения к человеческой деятельности, прошедшие через пищеварительный тракт пещерных гиен и волков, с ясно выраженными следами воздействия кислотной коррозии. Преимущественно это истонченные, корродированные и заполированные пластинки от трубчатых и плоских костей, с неровной волнистой поверхностью, часто со множеством мелких сквозных отверстий. Отчетливые следы пищеварительной ферментации здесь имеет и большинство изолированных зубов, например, свыше 97 % всех зубов Capra/Ovis .
Удельный вес о статков хищников в Чагырской пещере составляет 23,9 % в слое 5 и 18,5 % в слое 6, что
Таблица 1. Видовой состав и количество костных остатков
|
Таксон |
Слой |
||||||||||
|
1 |
2 |
3 |
4 |
5 |
6а |
6б |
6в |
7 |
Отвал |
Всего |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
Canis familiaris |
– |
– |
– |
– |
– |
– |
1 * |
– |
– |
– |
1 |
|
Equus caballus |
– |
– |
2 |
– |
– |
– |
– |
– |
– |
– |
2 |
|
Bos taurus |
– |
– |
2 |
– |
2 * |
– |
– |
– |
– |
– |
4 |
|
Capra/Ovis (дом.) |
4 |
– |
9 |
5 |
8 * |
3 * |
5 * |
– |
– |
4 |
38 |
|
Asioscalops altaica |
– |
– |
1 |
– |
15 |
20 |
7 |
4 |
– |
3 |
50 |
|
Chiropthera gen. indet. |
– |
– |
4 |
– |
– |
4 |
1 |
– |
– |
1 |
11 |
|
Ochotona sp . |
– |
– |
1 |
– |
1 |
– |
– |
– |
– |
– |
2 |
|
Lepus timidus |
– |
3 |
2 |
6 |
– |
– |
– |
– |
– |
– |
11 |
|
L. tanaiticus |
– |
– |
1 ** |
– |
8 |
6 |
6 |
– |
– |
3 |
24 |
|
L. tolai |
– |
– |
3 |
1 |
11 |
5 |
6 |
6 |
– |
2 |
34 |
|
Tamias sibiricus |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
1 |
|
|
Citellus sp. |
– |
5 |
13 |
7 |
93 |
50 |
32 |
12 |
– |
8 |
220 |
|
Marmota baibacina |
1 |
– |
3 |
– |
11 |
6 |
2 |
2 |
– |
– |
25 |
|
Castor fiber |
– |
– |
1 |
1 |
– |
– |
1 |
– |
– |
– |
3 |
|
Allactaga sp. |
– |
– |
1 ** |
– |
6 |
2 |
– |
– |
– |
– |
9 |
|
Cricetus sp. |
– |
12 |
11 |
9 |
5 |
7 |
– |
– |
– |
1 |
45 |
|
M. myospalax |
– |
7 |
9 |
1 |
101 |
63 |
32 |
16 |
– |
12 |
241 |
|
Arvicola terrestris |
– |
2 |
4 |
5 |
2 |
2 |
4 |
– |
– |
2 |
21 |
|
Rodentia gen. indet. |
– |
7 |
10 |
16 |
123 |
65 |
28 |
14 |
– |
11 |
274 |
|
Canis lupus |
– |
1 ** |
2 |
– |
22 |
19 |
31 |
28 |
2 |
4 |
109 |
|
Vulpes vulpes |
1 |
1 |
2 (1 ** ) |
2 |
26 |
21 |
34 |
33 |
1 |
9 |
130 |
|
V. corsak |
– |
– |
– |
3 |
12 |
15 |
21 |
26 |
1 |
3 |
68 |
|
Cuon alpinus |
– |
– |
– |
– |
11 |
6 |
2 |
6 |
– |
2 |
27 |
|
Ursus arctos |
– |
– |
2 |
– |
5 (1 ** ) |
1 |
– |
2 |
– |
– |
10 |
|
Martes zibellina |
– |
– |
– |
1 |
3 |
– |
1 |
– |
– |
– |
5 |
|
M. nivalis |
– |
– |
– |
– |
1 |
– |
– |
– |
– |
– |
1 |
|
M. erminea |
– |
– |
– |
– |
1 |
– |
– |
1 * |
– |
– |
1 |
|
M. altaica |
– |
1 |
– |
– |
3 (1 ** ) |
– |
– |
2 |
– |
– |
6 |
|
M. eversmanni |
– |
– |
– |
– |
2 |
– |
– |
– |
1 |
3 |
|
|
C. crocuta spelaea |
– |
– |
2 ** |
– |
19 |
14 |
12 |
8 |
– |
5 |
60 |
|
Panthera spelaea |
– |
– |
– |
– |
2 |
– |
– |
2 |
– |
– |
4 |
|
Mammuthus primigenius |
– |
– |
– |
– |
25 |
4 |
16 |
9 |
– |
4 |
58 |
|
Equus (E.) ferus |
– |
– |
– |
– |
5 |
4 |
4 (1 * ) |
2 |
– |
1 |
16 |
|
E. (Sussemionus) ovodovi |
– |
– |
– |
– |
6 |
23 |
30 |
26 |
– |
4 |
91 |
|
E. ovodovi/ferus |
– |
– |
– |
1 ** |
32 |
49 |
19 |
15 |
– |
3 |
119 |
|
Coelodonta antiquitatis |
– |
– |
– |
– |
7 |
5 |
5 |
1 |
– |
– |
18 |
|
Cervus elaphus |
– |
4 |
15 |
9 |
11 (5 * ) |
13 (3 * ) |
12 (1 * ) |
10 |
– |
7 (6 * ) |
82 |
|
Alces alces |
– |
– |
– |
– |
1 * |
2 * |
– |
– |
– |
1 * |
4 |
|
Caprolus pygargus |
1 |
– |
13 |
2 |
21 * |
3 * |
– |
1 * |
1 * |
1 * |
43 |
|
Rangifer tarandus |
– |
– |
– |
2 |
2 |
4 |
5 |
– |
1 |
14 |
|
|
Bison priscus |
– |
– |
3 ** |
1 ** |
29 |
117 |
158 |
313 |
12 |
37 |
669 |
|
Saiga tatarica borealis |
– |
– |
– |
– |
4 |
4 |
– |
2 |
– |
– |
10 |
Окончание табл. 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
Saiga/Procapra |
– |
– |
– |
– |
20 |
13 |
9 |
– |
– |
1 |
43 |
|
Capra sibirica |
– |
– |
5 ** |
– |
94 |
68 |
57 |
64 |
– |
10 |
298 |
|
Ovis ammon |
– |
– |
4 ** |
– |
38 |
31 |
24 |
24 |
1 |
8 |
131 |
|
Capra/Ovis |
– |
5 ** |
– |
– |
60 |
57 |
40 |
27 |
– |
9 |
81 |
|
Pisces |
– |
– |
16 |
– |
4 |
1 |
1 |
– |
– |
2 |
24 |
|
Aves |
2 |
19 |
17 |
12 |
51 |
27 |
18 |
13 |
– |
11 |
170 |
|
Неопределимые обломки |
31 |
64 |
1 645 |
434 |
22 946 |
18 729 |
25 993 |
21 564 |
759 |
4 460 |
96 634 |
|
Всего |
40 |
131 |
1 805 |
517 |
22 839 |
19 467 |
26 629 |
22 236 |
777 |
4 632 |
100 072 |
-
*Кости голоценовой сохранности в плейстоценовых слоях 5–7.
-
**Кости плейстоценовой сохранности в голоценовых слоях 1–4.
Таблица 2. Соотношение остатков видов крупных млекопитающих в плейстоценовых отложениях, %
|
Элементы скелета |
Лисицы |
Волки |
Гиена |
Мамонт |
Носорог |
Лошади |
Бизон |
Сайгак/ дзерен |
Козлы/ бараны |
|
Изолированные зубы |
61,2 |
52,7 |
71,2 |
100 |
100 |
90,8 |
76,3 |
67,9 |
64,5 |
|
Обломки черепа, верхней и нижней челюстей |
9,5 |
6,9 |
3,5 |
– |
– |
0,4 |
3,3 |
– |
1,4 |
|
Кости дистальных отделов конечностей |
19,9 |
33,6 |
20,3 |
– |
– |
8,4 |
13,6 |
32,1 |
29,7 |
|
Обломки крупных трубчатых костей, лопатки, таза |
5,0 |
3,8 |
5,1 |
– |
– |
0,4 |
1,6 |
– |
4,2 |
|
Позвонки, ребра |
4,5 |
3,1 |
– |
– |
- |
– |
5,1 |
– |
0,2 |
|
Всего остатков, ед. |
201 |
131 |
59 |
57 |
19 |
238 |
668 |
53 |
589 |
Таблица 3. Распределение фрагментов костей крупных млекопитающих по размерным классам
(12,7 %), чем на большинстве «археологических» пещерных местонахождений [Природная среда…, 2003; Васильев, Оводов, 2013]. Такой непропорционально большой удельный вес остатков Carnivora никогда не наблюдается не только в естественных биоценозах, но и в довольно точно отображающих их аллювиальных тафоценозах, где доля костей хищников, как правило, не поднимается выше 1–2 %.
Гиены и волки известны не только как сборщики падали, но и как весьма успешные коллективные охотники. Их добычей становились копытные среднего размерного класса – лошади, олени, сайгаки, горные козлы и бараны. Крупные хищники являются гораздо более универсальными коллекторами костных остатков, чем человек, охотничья деятельность которого часто носила черты специализации. Пещерные гиены собирали со всех ближайших окрестностей пещеры любые доступные им останки – от бобра до мамонта. В итоге подобное собрание костей, занесенных крупными хищниками в пещеру, наиболее полно и точно отражает особенности существовавшего некогда биоценоза.
По мнению Г.Ф. Барышникова [Природная среда…, 2003], большинство мелких фрагментов костей со следами пищеварительной ферментации попадало в пещерные отложения из распавшихся копролитов гиен. Часть более крупных остатков, включая изолированные зубы, происходит, несомненно, из отрыжек этих животных. Наблюдения за поведением современных пятнистых гиен ( C. crocuta ) показали, что они регулярно освобождают желудок, отрыгивая непереваренные остатки пищи, состоящие в основном из шерсти и по-лурастворившихся фрагментов костей. Обычно гиены, предварительно выкатавшись как следует на этом волосяном комке, вновь старательно выискивают в нем кусочки костей, разгрызая и заглатывая их [Лавик-Гу-долл, 1977]. С учетом всего вышесказанного, а также принимая во внимание очень медленный процесс осадконакопления в пещере (вероятно, не более 5–10 см за 1 тыс. лет), не приходится удивляться крайней скудости и фрагментарности сохранившихся в пещерных слоях остатков крупных млекопитающих.
Изучение остеологических материалов показало, что на определенном этапе (слои 6б, 6в/1) аккумуляция костных остатков в Чагырской пещере происходила в том числе и в результате более или менее продолжительного обитания здесь палеолитического человека, который специализировался в охоте на крупных стадных копытных, в первую очередь бизона. Однако большую часть времени эта пещера, как и другие пещерные местонахождения Алтая, служила убежищем или логовом для выведения потомства пещерным гиенам, волкам и лисицам. Материалы 2009–2011 гг. указывают на постепенное выклинивание по направлению в глубину пещеры основного тела костеносной линзы, накопившейся в результате охотничьей деятельности человека. На других участках, за пределами этого антропогенного скопления, аккумуляция остатков мегафауны в пещерных отложениях происходила главным образом за счет пищевой активности крупных хищников. Можно предположить, что в периоды отсутствия человека волки и гиены проводили в пещере тщательную ревизию оставленных отбросов, поедая или растаскивая все сколько-нибудь ценное в пищевом отношении. Так, остатки бизона представлены почти исключительно изолированными зубами, гораздо реже – короткими костями дистальных отделов конечностей и их фрагментами. Крупные обломки эпифизов трубчатых костей бизона или лошадей полностью отсутствуют. Обильно представлены лишь уплощенные и небольшие (до 5–10 см) толстостенные фрагменты диафизов, которые, в отличие от более мягких, насыщенных костным мозгом тканей эпифизов, почти не привлекали хищников. Кроме того, они легче попадали в захоронение благодаря своим размерам и конфигурации.
По костным остаткам следы присутствия человека уверенно прослеживаются в слое 6в, несколько слабее они выражены в слое 6б, а в слоях 6а и 5 пока не зафиксированы, хотя в слое 6а/3 найдены единичные ко сти человека. К слоям 6б и 6в приурочены также практически все палеоантропологические находки. Выпавший резец ребенка (слой 6б) свидетельствует о достаточно продолжительном периоде обитания в пещере людей, включая женщин и детей.
Палеоэкологические реконструкции
В материалах слоя 5 определимых ко стей крупных млекопитающих значительно (в 3,6 раза) меньше, чем в коллекции из слоя 6. По сравнению с 6-м слоем в 5-м относительное количество остатков бизона резко (в 5,7 раза) сокращается, а сибирского горного козла и архара, напротив, существенно (в 1,8 раза) увеличивается. В 1,4 раза уменьшается число костей благородного оленя. Удельный вес остатков лошади в слоях 5 и 6 приблизительно одинаков (10,0 и 11,1 % соответственно). Кости сайгака/дзерена встречаются в 5-м слое более чем в 3 раза чаще, чем в 6-м (рис. 1). Чем же вызваны столь контрастные изменения в относительном обилии фоновых видов мегафауны? Наиболее вероятны два объяснения. Первое – ариди-зация и похолодание климата в период накопления слоя 5, что обусловило существенную перестройку биоты. Второе объяснение предполагает отсутствие крупных изменений в структуре данных биоценозов. В этом случае накопление костных остатков в период формирования слоя 5 происходило в основном за счет пищевой активности пещерных гиен и волков, акку- муляция которыми костных остатков носила универсальный характер. Их добычей становились копытные среднего размерного класса – лошади, олени, горные козлы, бараны, а по случаю и молодняк крупных копытных и толстокожих, таких как бизон, мамонт и шерстистый носорог. Значительную роль в формировании тафоценоза слоя 5 играл также сбор этими хищниками любой доступной падали в окрестностях пещеры.
В период накопления слоя 6 процесс «естественного» формирования тафоценоза был нарушен наложением антропогенного фактора, в результате чего заметно исказилось соотношение фоновых видов мегафауны. Палеолитический человек, посещавший пещеру, специализировался в охоте на крупных стадных копытных, прежде всего бизона, обломки костей которого с этого момента в больших количествах начали захоронять-ся в пещерных отложениях, преимущественно в приустьевой части. Примечательно, что по мере продвижения в глубь пещеры (раскопки 2009–2011 гг.) их чис- ло в слое 6 заметно сокращается – с 52 (2007–2008 гг.) до 32 % (2009–2011 гг.). Одновременно с этим увеличивается количество остатков Capra/Ovis и лошадей – соответственно в 2,2 и 1,7 раза. Значительная часть костей из раскопок 2009–2011 гг. отмечена признаками деструкции от кислотной коррозии. Скорее всего, это связано с постепенным выклиниванием в глубь пещеры основного тела костеносной линзы, накопившейся по преимуществу в результате охотничьей деятельности человека. Таким образом, в составе тафоценоза слоя 6 в глубине пещеры просматриваются черты, общие с тафоценозом слоя 5, формировавшимся, по-видимому, без сколько-нибудь заметного участия человека.
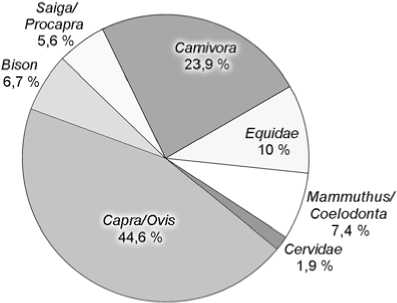
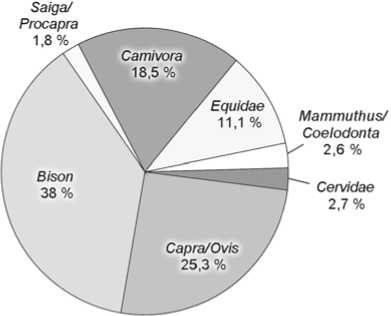
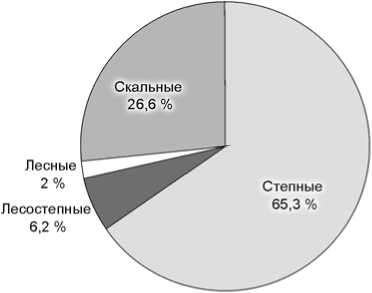
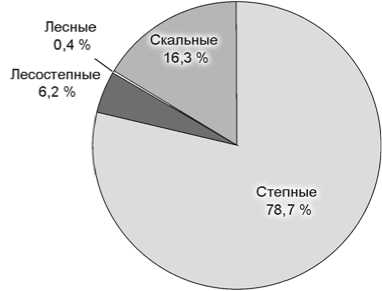
Не считая костей представителей интразональных видов-убиквистов (таких как волк, лисица, мамонт), остатки мегафауны из слоев 5 и 6 Чагырской пещеры в основном принадлежат обитателям открытых, степных пространств (рис. 2). Доля костей млекопитающих степных биотопов составляет для 5-го слоя 65,3 %,
Слой 5 (431 кость)
Слой 6 (1 547 костей)
Слой 5 (357 костей)
Рис. 2. Соотношение костных остатков крупных млекопитающих различных биотопических групп в плейстоценовых слоях.
Рис. 1. Соотношение костных остатков крупных млекопитающих различных видов в плейстоценовых слоях.
Слой 6 (1 157 костей)
6-го – 78,7 %, лесостепных – соответственно 6,2 и 4,7 %, лесных – 1,2 и 0,4, скальных – 26,6 и 16,3 %. Более чем трехкратное увеличение остатков сайгака/дзе-рена и почти двухкратное – сибирского горного козла в слое 5 объясняется, возможно, аридизацией климата, нарастанием остепнения и расширением площадей слабозадернованных скальных выходов – основных биотопов Capra sibirica . Почти в 3 раза возрастает в слое 5 число остатков таких представителей относительно холодолюбивых видов, как мамонт и шерстистый носорог (рис. 2).
Первобытный бизон был обитателем открытых ландшафтов. Его пространственно-временные формы отличались значительной экологической пластичностью. Бизоны населяли лесостепные, степные, тундростепные ландшафты. На Алтае в позднем плейстоцене обитал Bison priscus – очень крупная горностепная форма [Васильев, Оводов, 2009]. В слое 5 Ча-гырской пещеры доля костей бизона (6,7 %) близка к аналогичным показателям других пещерных местонахождений Алтая, а в слое 6 – резко увеличена (38 %), вероятно, благодаря специализации палеолитических охотников на добыче этого зверя.
Относительно длинные метаподии, узкие и высокие копытные фаланги указывают на связь лошади Оводова ( Equus ovodovi ) с умеренно-аридными ландшафтами и твердыми грунтами. По этим признакам она уступает только плейстоценовым ослу и кулану – обитателям аридных степей и полупустынь. Второй вид лошади, чьи остатки были обнаружены в пещере, – E. ferus . Она обладала массивными метаподиями и широкими копытными фалангами. Эта крупная тяжелая лошадь или ее близкородственные формы ( E . ex. gr. gallicus ) находили свой экологический оптимум и были наиболее многочисленны в лесостепной зоне либо на севере степной, где преобладали относительно мягкие (зачастую увлажненные, как в современной Барабе) грунты. Тот факт, что по количеству остатков E. ovodovi почти в 9 раз превосходит E. ferus, является прямым указанием на господство умеренно-аридных степных ландшафтов в момент формирования слоя 6.
К остаткам типично лесных видов относятся лишь четыре находки из слоев 5 и 6 – кости и зубы соболя. Небольшие лесные участки, по-видимому, были приурочены к долинам рек и горным ущельям. На существование приречных лесов указывает также найденный в слое 6б изолированный зуб бобра. Присутствие единичных остатков бурого медведя едва ли можно использовать в качестве индикатора лесных ландшафтов. Плейстоценовый Ursus arctos Южной Сибири по своей экологии, вероятнее всего, был сходен с современным медведем-пищухоедом (Ursus arctos pruinosus), населяющим ныне нагорные степи Северного Тибета. Входившие в состав мамонтовой фауны благородный олень и лось, в отличие от современных, также не имели облигатной связи с лесными местообитаниями и находили вполне приемлемые условия для своего существования в степных и лесостепных ландшафтах. Наглядным свидетельством этого служат морфофункциональные особенности строения их нижней челюсти [Васильев, 2005б, 2011]. Глубина снежного покрова, судя по наличию остатков сайгака, скорее всего, не превышала 15–20 см. Присутствие костей северного оленя позволяет предположить формирование слоев 5 и 6 в холодные интервалы позднего плейстоцена.
По палинологическим данным лесостепь реконструируется для времени накопления слоя 5 и верхней половины слоя 6а. Нижняя его толща, слои 6б и 6в/1 формировались в условиях господства степных ландшафтов. Для слоев 6в/2 и 7 отмечено наличие лесной растительности с участием холодных листопадных лесов [Рудая, 2011]. Однако почти все определимые костные остатки крупных млекопитающих из этих слоев (не считая пяти костей волка и лисицы – видов-убиквистов) принадлежат обитателям степных (66 %) и скальных (29,8 %) биотопов, лишь две кости – северному оленю, который условно может быть отнесен к тундровому (4,2 %).
Таким образом, остатки крупных млекопитающих позволяют реконструировать для периода накопления слоя 6 (в т.ч. 6в/2, а также слоя 7) господство открытых степных ландшафтов. Время формирования слоя 5 было отмечено, по-видимому, существенной аридиза-цией климата. Наряду с этим на протяжении всего периода осадконакопления в позднем плейстоцене по долинам рек, в горных ущельях, а возможно, местами и на склонах гор северной экспозиции могли сохраняться относительно небольшие участки лесных массивов.
Систематический обзор
Бобр ( Castorfiber ). Изолированный правый М1 найден в слое 6б/1. Длина коронки 6,2 мм, ее ширина 7,2, высота зуба 24,5 мм. В голоценовом слое 3 найдена целая плюсневая кость крупной особи, в слое 4 – венечный отросток нижней челюсти. Единичные остатки бобра (преимущественно зубы) отмечены на большинстве плейстоценовых пещерных местонахождений Алтая.
Волк ( Canis lupus ). Его остатки встречаются во всех плейстоценовых слоях. В количественном отношении волк лишь незначительно уступает лисице и почти вдвое превосходит пещерную гиену. Для промеров и сравнения подходит лишь небольшое число изолированных зубов и два фрагмента нижней челюсти. Несмотря на незначительную величину выборки, можно констатировать большое сходство в размерах зубов с современным волком Алтая (табл. 4). Чагыр-ская пещера служила весьма удобным логовом для этого хищника. В других пещерах Горного Алтая ос-
Таблица 4. Размеры зубов Canis lupus
|
Промеры, мм |
Чагырская пещера |
Алтай, современность (C. lupus altaicus) |
||||
|
n |
Lim |
M |
n |
Lim |
M |
|
|
Длина P2 |
1 |
– |
14,30 |
7 |
13,7–15,5 |
14,34 |
|
Ширина P2 |
1 |
– |
6,10 |
6 |
5,6–7,2 |
6,37 |
|
Длина P3 |
1 |
– |
15,70 |
6 |
15,5–16,4 |
15,97 |
|
Ширина P3 |
1 |
– |
7,20 |
6 |
6,7–7,7 |
7,25 |
|
Длина P4 |
5 |
22,0–27,2 |
24,38 |
8 |
24,0–27,4 |
25,39 |
|
Ширина P4 |
5 |
11,0–15,5 |
12,94 |
8 |
12,1–14,8 |
13,26 |
|
Длина M1 |
2 |
15,4–18,0 |
16,70 |
8 |
15,5–17,4 |
16,46 |
|
Ширина M1 |
2 |
18,2–20,3 |
19,25 |
8 |
20,3–23,0 |
21,78 |
|
Длина M1 – M3 |
1 |
– |
42,50 |
10 |
45,0–48,8 |
46,28 |
|
Длина P2 |
1 |
– |
13,40 |
9 |
11,0–13,1 |
12,29 |
|
Ширина P2 |
1 |
– |
5,70 |
9 |
5,2–7,0 |
6,13 |
|
Длина P3 |
2 |
13,6–15,0 |
14,30 |
10 |
13,0–14,2 |
13,78 |
|
Ширина P3 |
2 |
5,8–7,0 |
6,40 |
9 |
5,4–8,0 |
6,58 |
|
Длина P4 |
3 |
14,8–17,2 |
16,10 |
10 |
15,2–16,3 |
15,90 |
|
Ширина P4 |
3 |
7,5–8,5 |
8,03 |
10 |
6,2–8,6 |
7,87 |
|
Длина M1 |
2 |
27,0–29,7 |
28,35 |
10 |
26,7–31,5 |
28,66 |
|
Ширина M1 |
2 |
11,1–12,7 |
11,90 |
10 |
9,9–13,2 |
11,42 |
|
Длина M2 |
4 |
11,0–12,4 |
11,53 |
10 |
11,0–13,3 |
12,17 |
|
Ширина M2 |
4 |
8,0–9,0 |
8,58 |
10 |
8,1–9,6 |
8,91 |
|
Длина M3 |
2 |
5,2–6,7 |
5,95 |
7 |
5,6–6,5 |
6,14 |
|
Ширина M3 |
2 |
5,3–5,7 |
5,50 |
7 |
5,2–6,0 |
5,67 |
татки пещерной гиены, как правило, значительно преобладают над волчьими [Васильев, Оводов, 2013].
Лисица ( Vulpes vulpes ). По числу остатков лисице принадлежит первое место среди хищных. Преобладают изолированные зубы и мелкие ко сти дистальных отделов конечностей. Длина 6 экз. М1 16,1– 18,3 ( М 16,98) мм, ширина – 5,3–7,0 ( М 6,50) мм. Ширина нижнего конца берцовой кости 13,3 мм, поперечник – 9 мм. Длина пяточной кости 33,2 мм. Часть костей и зубов несет признаки кислотной коррозии. Пещерные гиены и волки могли утилизировать останки погибших по тем или иным причинам лисиц, как находившиеся в пещере, так и занесенные ими извне. Судя по тому, что молочных зубов лисиц (в т.ч. и корсака) в отложениях не отмечено, пещера в основном использовалась ими в качестве временного убежища.
Корсак (Vulpes corsak). Остатки этой мелкой степной лисицы встречаются в отложениях пещеры почти в 2 раза реже, чем V. vulpes. Сохранились четыре целых верхних хищнических зуба (Р4). Длина коронки составляет 12,0–12,3 (М 12,15) мм, ширина – 4,3–4,7 (М 4,53) мм. Два обломка нижней челюсти имеют альвеолярную длину зубного ряда 29,0 и 29,6 мм. Длина 4 экз. М1 13,2–14,0 (М 13,73) мм, ширина – 5,1–5,6 (М 5,33) мм. Две нижние половины лучевой кости имеют ширину диафиза 6,7 и 6,8 мм, поперечник – 3,6 и 4,6, ширину нижнего конца 11, поперечник – 6,2 и 6,4 мм. Длина трех разъеденных кислотной коррозией пяточных костей 22,4; 24,3 и 27,5 мм.
Красный волк ( Cuon alpinus ). Зубы и мелкие кости дистальных отделов конечностей зафиксированы от слоя 5 (горизонт 3) до слоя 6в/1 (горизонт 3). Большая их часть повреждена кислотной коррозией. Три экземпляра М2 имеют длину 9,5; 9,7 и 10,1 мм, ширину 5,2; 5,9 и 6,1 мм. Остатки красного волка повсеместно встречаются в отложениях пещер Алтая, но обычно в 2–3 раза реже, чем серого волка.
Бурый медведь (Ursus arctos). В голоценовом слое 3 найдены резец и обломок сильно стертого коренного зуба, в слое 5 (горизонты 1, 4, 5) – три первые фаланги (одна из них голоценовой сохранности), резец и фрагмент коренного зуба, в слое 6а/1 – обломок верхнего конца первой фаланги, в слое 6в/1 (горизонт 1) – третья задняя фаланга. Данные остатки слишком скудны для морфометрического анализа. Материалы пещеры Страшной показывают, что в позднем плейстоцене в Горном Алтае обитал бурый медведь, превосходящий по размерам костей и зубов самых крупных голоценовых представителей этого вида [Васильев, Гребнев, 2009]. Наряду с его остатками в гораздо меньшем количестве в алтайских пещерах найдены зубы и кости малого пещерного (Ursus savini – Денисова и Страшная) и пещерного (Ursus spelaeus – Страшная) медведя [Васильев, Зенин, 2009; Васильев, Шуньков, Цыбанков, 2008; Knapp et al., 2009].
Соболь ( Martes zibellina ). В слоях 5 (горизонт 4) и 6б/1 обнаружены два клыка верхней челюсти длиной 22,0 и 20,3 мм, в голоценовом слое 4 – аналогичный клык плейстоценовой сохранности, поврежденный кислотной коррозией, длиной более 19,5 мм, в слое 5 (горизонты 4, 5) – две проксимальные половины плюсневых костей. Единичные остатки соболя отмечены в плейстоценовых отложениях большинства пещерных местонахождений Алтая. Они свидетельствуют о том, что на фоне господства открытых ландшафтов по долинам рек и в горных ущельях местами сохранялись более или менее значительные участки лесной растительности.
Степной хорь (Mustela eversmanni). Левая ветвь нижней челюсти с неполным М1 и не затронутый стиранием М1 другой особи найдены в слое 6а/1. Длина челюсти от резцов до суставного отростка более 40,5 мм, альвеолярная длина зубного ряда (Р2 – М2) 19,2 мм. Высота челюсти за М1 8,8 мм, толщина под ним 4,9 мм. В материалах промывки отвала обнаружена целая плечевая кость плейстоценовой сохранности. Ее длина от головки 43,4 мм, ширина верхне- го конца, диафиза и нижнего конца соответственно 9,8; 3,5 и 11,8 мм. Остатки степного хоря являются хорошим индикатором палеосреды, указывая на господство открытых ландшафтов.
Пещерная гиена ( C. crocuta spelaea ). Значительная часть костных остатков со следами пищеварительной ферментации происходит из отрыжек и распавшихся копролитов, по всей видимости, именно этого хищника. Присутствие молочных зубов указывает на то, что пещера использовалась кланами гиен для выведения потомства. В слое 6а/3 найден погрызенный фрагмент нижней челюсти с Р2 – М1. Из костей посткраниального скелета сохранилось семь первых и третьих фаланг и их обломков, центральная кость заплюсны, фрагменты верхнего конца лучевой и локтевой костей, обломок нижнего конца метаподии. Нижний отдел малой берцовой кости (слой 6б/1) имеет ширину 18,3 мм, поперечник 10,8 мм. В слое 6в/1 (горизонт 1) найден неполный копролит гиены диаметром 24 мм. Промеры изолированных зубов и нижней челюсти приведены в табл. 5. Пещерная гиена – основной аккумулятор ко стных остатков в пещерных тафоценозах Западной Европы и гор Южной Сибири. Ее роль в этом качестве подробно рассмотрена в ряде статей [Васильев, Оводов, 2013; Diedrich, 2010, 2011; Diedrich, Žak, 2006; Kuch, Berger, Skinner, 2010].
Пещерный лев ( Panthera spelaea ). В слое 5 (горизонт 1) найден превосходно сохранившийся слабостертый М1. Его длина 32,6 мм, ширина – 15,7 мм.
Таблица 5. Размеры зубов C. crocuta spelaea
|
Промеры, мм |
Чагырская |
Денисова |
||||
|
n |
Lim |
M |
n |
Lim |
M |
|
|
Длина Р3 |
1 |
– |
24,50 |
3 |
24,3–27,7 |
25,80 |
|
Ширина Р3 |
1 |
– |
19,30 |
3 |
17,9–20,3 |
18,93 |
|
Длина Р4 |
1 |
– |
43,00 |
1 |
– |
40,50 |
|
Ширина Р4 |
1 |
– |
23,50 |
1 |
– |
23,00 |
|
Длина от задней стенки клыка до М1 |
1 |
– |
92,50 |
2 |
91,5–99,5 |
95,50 |
|
Длина диастемы |
1 |
– |
3,00 |
9 |
3,2–8,8 |
5,68 |
|
Альвеолярная длина Р2 – Р4 |
1 |
– |
58,00 |
7 |
55,0–63,0 |
58,50 |
|
Коронарная длина Р2 – Р4 |
1 |
– |
61,00 |
6 |
57,6–64,3 |
61,05 |
|
Толщина горизонтальной ветви под Р3 |
1 |
– |
21,00 |
8 |
17,8–22,3 |
20,63 |
|
Длина Р2 |
2 |
17,2–17,2 |
17,20 |
15 |
15,7–17,8 |
16,80 |
|
Ширина Р2 |
2 |
12,4–13,1 |
12,75 |
16 |
10,8–13,1 |
12,03 |
|
Длина Р3 |
2 |
21,1–22,0 |
21,55 |
11 |
21,5–25,5 |
23,45 |
|
Ширина Р3 |
2 |
17,0–17,2 |
17,10 |
11 |
15,4–18,3 |
16,84 |
|
Длина Р4 |
2 |
24,0–24,9 |
24,45 |
9 |
21,7–24,6 |
23,34 |
|
Ширина Р4 |
2 |
14,5–15,0 |
14,75 |
9 |
14,6–15,5 |
15,07 |
|
Длина М1 |
1 |
– |
36,30 |
9 |
31,3–34,4 |
32,58 |
|
Ширина М1 |
1 |
– |
14,60 |
13 |
12,3–14,6 |
13,42 |
Соответствующие промеры нижнего хищнического зуба у пещерных львов Евразии ( n = 31) – 23,7–33,2 ( М 28,41) и 11,5–16,3 ( М 14,11) мм [Верещагин, 1971; Алексеева, 1980]. Зуб из Чагырской пещеры принадлежал, таким образом, очень крупной особи. Из слоя 5 (горизонт 1) происходит также разъеденный кислотной коррозией характерно изогнутый диафиз первой фаланги. Его ширина посередине 13,6 мм. Поврежденная желудочной ферментацией вторая фаланга передней конечности (предположительно третья левая) обнаружена в слое 6в/1 (горизонт 1). Ее общая длина 37,3 мм, сагиттальная 36,5, ширина верхнего конца 18, поперечник – 20, ширина диафиза 11, поперечник – 12,1, ширина нижнего конца более 15 мм. В этом же слое найден обломок нижнего отдела метапо-дии с поперечником более 21 мм. Единичные остатки P. spelaea встречаются почти на всех пещерных местонахождениях Алтая [Оводов, Мартынович, 2008]. Судя по всему, львы, в отличие от пещерных гиен и волков, редко использовали пещеры в качестве убежища или логова для выведения потомства. Их остатки попадали в пещерные тафоценозы в основном в результате пищевой активности пещерных гиен.
Мамонт ( Mammuthus primigenius ). Его остатки представлены исключительно мелкими обломками молочных зубов и некрупных бивней. Длина 41 фрагмента пластинок зубов первых смен (pd 3–4) варьирует от 17 до 63 мм, а 16 обломков пластинок бивня – от 20 до 65 мм. Целиком сохранились три зуба мамонтят (pd 2), верхний и два нижних. Длина и ширина pd2 13,5 и 11,2 мм, pd2 – 10,5 и 8,5; 11,0 и 8,9 мм при количестве пластин в зубе 4, 4 и 3 соответственно. Все зубы мамонтят, как целые, так и обломки, несут признаки растворения кислотной коррозией в пищеварительном тракте крупных хищников и происходят из их отрыжек или распавшихся копролитов. Совершенно очевидно, что для попадания таких остатков в пещерные отложения гиенам совсем не обязательно было непосредственно заносить головы мамонтят под своды пещеры. У крупных трубчатых костей взрослых мамонтов, в силу их размеров и большой прочности, пещерные гиены могли лишь частично выгрызать губчатую ткань эпифизов. Трупы же мамонтят первых лет жизни, по-видимому, утилизировались ими целиком, и зубы заглатывались вместе с кусками альвеол. Присутствие в пещерных тафоценозах почти исключительно обломков зубов мамонтят первых трех смен (pd 2–4) характерно и для других пещерных памятников Южной Сибири, таких как Денисова пещера, Страшная, Логово Гиены или грот Проскурякова [Васильев, Оводов, 2013].
Крупная кабаллоидная лошадь ( Equus ferus ). Представлена в основном обломками зубов, не пригодными для морфометрического анализа. Неполная задняя копытная фаланга имеет ширину 75,6 мм, ширину и высоту суставной поверхности 48 и 26 мм.
Ширина аналогичной третьей фаланги Equus ex. gr. gallicus из Тараданово (Предалтайская равнина) 72,3– 88,5 ( М 78,79) мм ( n = 47), ширина и высота суставной поверхности – 45,0–58,2 ( М 50,82) ( n = 57) и 23,5–31,5 ( М 27,75) мм ( n = 62). В позднем плейстоцене на Алтае обитала особая, очень крупная и массивная форма кабаллоидной лошади. По строению метаподий она не может быть отождествлена с обитавшей на равнинной части юга Западной Сибири близкородственной E. ex. gr. gallicus . Г.Ф. Барышниковым крупная форма лошади, кости которой найдены в пещерах Алтая, предварительно была обозначена как E. ferus [Природная среда…, 2003].
Лошадь Оводова (Equus (Sussemionus) ovodovi). В позднеплейстоценовых отложениях пещер Алтая и Хакасии повсеместно присутствуют остатки двух форм лошади – крупной кабаллоидной и мелкой, которая ранее отождествлялась с куланом [Галкина, Оводов, 1975] или плейстоценовым ослом [Природная среда..., 2003; Васильев, Оводов, Мартынович, 2006]. Проведенные биомолекулярные и морфологические исследования показали: эта мелкая форма не может быть отнесена ни к плейстоценовому ослу или кулану, ни к представителям кабаллоидных лошадей, а является совершенно новой отчетливо выраженной монофилетической группой [Orlando et al., 2009; Eisenmann, Vasiliev, 2011]. Ранее считалось, что все представители подрода Sussemionus, распространенные в раннем плейстоцене от Северной Америки и Евразии до Африки, вымерли приблизительно 0,5 млн л.н. На основе материалов из пещер Хакасии и Алтая был описан новый вид, названный в честь его первооткрывателя лошадью Оводова [Eisenmann, Vasiliev, 2011; Васильев, 2013]. По высоте в холке (137–160 см, М 146) эта лошадь приблизительно соответствовала современному монгольскому кулану E. h. hemionus, однако была более тяжелой и массивной, на что указывают ее менее стройные метаподии. Кости E. ovodovi присутствуют на большинстве пещерных местонахождений Алтая, Хакасии, а также в аллювиальных отложениях Предалтайской равнины [Васильев, 2005а; Косинцев, Васильев, 2009]. В позднем плейстоцене, от казанцевского (R–W) до сартанского (W-3) времени включительно, они практически повсеместно отмечены в пещерных отложениях Алтая и Хакасии. На различных местонахождениях относительное обилие остатков E. ovodovi существенно различается. Они составляют свыше половины всех остатков лошадей в гроте Проскурякова, около половины – в Денисовой пещере, ок. 1/3 – в Страшной, наиболее многочисленны в пещерах Логово Гиены и Чагырской, где встречаются в 5–9 раз чаще, чем E. ferus. В Чагырской пещере остатки лошади Оводова обнаружены во всех плейстоценовых слоях (кроме слоя 7, где определимые остатки единичны). Для морфометрического анализа оказались пригодными изолированные зубы и первая передняя фаланга. По размерам зубов и их морфологии (в частности, относительной длине протокона и постфлекси-да) E. ovodovi из Чагырской пещеры мало отличается от лошади из Логова Гиены (табл. 6). Размеры и про- порции первой фаланги также весьма близки к таковым
E. ovodovi из Логова Гиены и Тараданово (табл. 7).
Шерстистый носорог (Coelodonta antiquitatis).
Из 18 небольших фрагментов зубов 13 принадлежали
Таблица 6. Размеры зубов Equus (Sussemionus) ovodovi
|
Промеры, мм |
Верхние зубы |
Нижние зубы |
||||||||||
|
Чагырская |
Логово Гиены |
Чагырская |
Логово Гиены |
|||||||||
|
n |
Lim |
M |
n |
Lim |
M |
n |
Lim |
M |
n |
Lim |
M |
|
|
Длина зуба |
1 |
– |
35,30 |
80 |
P 2 32,3–40,8 |
35,62 |
4 |
30,0–31,7 |
31,00 |
66 |
26,8–37,5 |
31,93 |
|
Длина протокона / постфлексида |
1 |
– |
6,70 |
95 |
5,2–10,5 |
8,02 |
4 |
14,3–15,2 |
14,73 |
64 |
11,3–18,8 |
14,54 |
|
Ширина зуба |
1 |
– |
23,70 |
92 |
20,4–27,7 |
24,48 |
4 |
14,7–16,0 |
15,10 |
68 |
13,4–18,0 |
15,90 |
|
Индекс протокона / постфлексида |
1 |
– |
18,98 |
80 |
15,9–29,8 |
22,52 |
4 |
45,1–49,0 |
47,52 |
61 |
35,5–64,8 |
45,54 |
|
Длина зуба |
1 |
– |
27,00 |
153 |
P 3–4 23,0–29,8 |
26,93 |
5 |
24,0–26,7 |
25,60 |
162 |
22,5–30,8 |
27,01 |
|
Длина протокона / постфлексида |
1 |
– |
8,90 |
154 |
7,2–13,7 |
10,83 |
5 |
8,1–13,0 |
11,24 |
157 |
7,2–16,0 |
11,96 |
|
Ширина зуба |
1 |
– |
25,40 |
137 |
23,7–29,6 |
26,60 |
5 |
15,6–17,7 |
16,48 |
158 |
14,7–19,8 |
17,14 |
|
Индекс протокона / постфлексида |
1 |
– |
32,96 |
152 |
27,3–51,5 |
40,22 |
5 |
33,8–50,0 |
43,91 |
156 |
30,0–57,6 |
44,28 |
|
Длина зуба |
2 |
26,2–28,5 |
27,35 |
216 |
M 1–2 20,2–28,3 |
24,27 |
4 |
23,3–27,5 |
24,70 |
186 |
21,6–29,2 |
25,46 |
|
Длина протокона / постфлексида |
2 |
10,0–11,0 |
10,50 |
225 |
8,5–13,5 |
11,42 |
5 |
7,0–10,3 |
8,62 |
177 |
6,3–14,0 |
9,57 |
|
Ширина зуба |
2 |
26,0–26,7 |
26,35 |
203 |
23,5–30,0 |
26,36 |
5 |
13,2–15,7 |
14,34 |
173 |
11,7–18,0 |
14,76 |
|
Индекс протокона / постфлексида |
2 |
38,2–38,6 |
38,39 |
213 |
33,9–60,2 |
47,05 |
4 |
29,4–39,3 |
34,90 |
176 |
26,5–48,3 |
37,59 |
|
Длина зуба |
– |
– |
– |
103 |
M3 24,5–32,0 |
27,31 |
2 |
31,3–33,0 |
32,15 |
67 |
28,5–34,2 |
31,44 |
|
Длина протокона / постфлексида |
– |
– |
– |
105 |
10,2–15,4 |
12,72 |
2 |
9,1–10,2 |
9,65 |
66 |
7,0–13,4 |
9,25 |
|
Ширина зуба |
– |
– |
– |
105 |
21,0–25,9 |
23,13 |
2 |
13,2–14,5 |
13,85 |
66 |
11,3–15,0 |
12,83 |
|
Индекс протокона / постфлексида |
– |
– |
– |
97 |
39,0–59,2 |
46,58 |
2 |
29,1–30,9 |
30,02 |
65 |
20,6–42,1 |
29,42 |
Таблица 7. Размеры первых передних фаланг Equus (Sussemionus) ovodovi
|
Промеры, мм |
Чагырская ( n = 1) |
Логово Гиены |
Тараданово |
||||
|
n |
Lim |
M |
n |
Lim |
M |
||
|
Наибольшая длина |
83,2 |
3 |
83,2–88,2 |
85,03 |
4 |
84,0–85,5 |
85,00 |
|
Сагиттальная длина, min |
78,0 |
3 |
78,1–81,7 |
79,83 |
4 |
78,7–80,2 |
79,45 |
|
Ширина верхнего конца |
45,3 |
3 |
43,8–45,5 |
44,83 |
4 |
44,7–48,3 |
46,25 |
|
Поперечник » » |
32,7 |
3 |
32,0–33,3 |
32,73 |
4 |
30,5–35,0 |
33,05 |
|
Ширина диафиза, min |
29,2 |
5 |
26,1–29,8 |
27,88 |
4 |
27,6–30,4 |
29,03 |
|
Ширина нижнего конца |
37,0 |
3 |
38,1–39,8 |
39,07 |
4 |
40,2–42,0 |
41,25 |
|
Поперечник » » |
21,6 |
3 |
21,0–22,7 |
21,90 |
4 |
21,2–23,0 |
21,93 |
молодым (juven.) особям. Все зубы, кроме двух, несут признаки кислотной коррозии. Из слоя 5 (горизонт 4) происходит неповрежденный правый Р2 взрослого животного, его длина 23,5 мм, ширина 15 мм. Присутствие остатков шерстистого носорога (равно как и мамонта) в пещерных отложениях не имеет, скорее всего, никакого отношения к деятельности палеолитического человека.
Марал ( Cervus elaphus sibiricus ). Фрагменты костей и зубов благородного оленя прослеживаются по всему разрезу плейстоценовых отложений пещеры. Часть из них имеет голоценовую сохранность. Дистальный отдел пястной кости из слоя 5 (горизонт 2) принадлежал некрупной особи, вероятно, самке – его ширина 45,2 мм, поперечник 33,5 мм, что находится на уровне минимальных значений у современного и голоценового марала Алтая [Васильев, 2005б]. Две запястные лучевые кости из слоев 5 (горизонт 3) и 6б/1 имеют длину в проекции 44,5 и 37,3 мм, ширину 25 и 20, высоту в переднем отделе 31,8 и 30,3 мм. Эти размеры находятся близ верхней и нижней границ изменчивости carpi radiale крупного голоценового марала Алтая, длина c. radiale которого 36,5–45,9 ( М 41,06) мм ( n = 46). Наибольшая длина третьей фаланги (слоя 6в/1, горизонт 2) 54,3 мм, дорзальная – 53 при высоте 37,3 мм. В слое 6б/1 найден роговой отросток длиной 192 мм. Позднеплейстоценовый марал юга Западной и Средней Сибири отличался очень крупными размерами тела. Особенно выделялись в этом отношении олени каргинско-го времени. В строении нижней челюсти позднеплейстоценового марала отмечены признаки, указывающие на его адаптацию к обитанию в условиях открытых – степных и лесостепных – ландшафтов [Там же].
Лось ( Alces alces ). Три обломка зубов и фрагмент второй фаланги, найденные в слоях 5, 6 и в материалах промывки отвала, имеют голоценовую сохранность. В позднеплейстоценовых отложениях Алтая остатки лося встречаются крайне редко. Они отмечены в пещерах Каминной и Страшной [Васильев, Деревянко, Маркин, 2006; Васильев, Зенин, 2009, 2010].
Косуля ( Capreolus pygargus ). Все остатки косули из плейстоценовых отложений имеют типично голоценовую сохранность и попали туда, несомненно, из слоев 1–4. В слое 5 (горизонт 1) в одном из квадратов помимо обломков центральнокубовидной и лучевой костей, пяти фрагментов 1–3-й фаланг, найдено десять целых астрагалов C. pygarus . Несомненно, что данные кости, не имеющие признаков обработки, были оставлены здесь человеком. В позднеплейстоценовых отложениях немногочисленные остатки косули отмечены в пещерах Денисовой и Каминной [Васильев, Деревянко, Маркин, 2006; Васильев, Шуньков, Цыбанков, 2008; Васильев, Шуньков, 2009].
Бизон ( Bison priscus ). Его остатки доминируют (34,4 %) в тафоценозе Чагырской пещеры. Преобладают изолированные зубы и их обломки (76,3 %). Посткраниальный скелет представлен целыми ко стями дистальных отделов конечностей и их обломками – фалангами, запястными и заплюсневыми, но имеются также единичные фрагменты подъязычных костей, ребер, позвонков (преимущественно хвостовых). Среди изолированных щечных зубов и резцов бизона к молодым (juven.) особям относится 4,3 %, полувзрослым (subad.) – 58, взрослым (ad.) – 30,4, старым (sen.) – 7,2 %. Немногие пригодные для измерений кости запястья, заплюсны и фаланги находятся на уровне минимальных значений промеров соответствующих костей Bison priscus Алтая и Верхнего Приобья (табл. 8). Все они принадлежали некрупным животным, скорее всего, самкам. Зубы нижней челюсти (Р2–4, М3), напротив, в средних и крайних значениях промеров мало отличаются от таковых Bison priscus Алтая и Верхнего Приобья (табл. 9). Число фрагментов костей и зубов бизона с признаками кислотной коррозии, в отличие от остатков других представителей мегафауны, весьма невелико. Наряду с этим следы порезов каменными орудиями встречаются исключительно на обломках
Таблица 8. Размеры костей конечностей Bison priscus
|
Промеры, мм |
Чагырская |
Логово Гиены, W-2 |
Тараданово, W-1–2 |
Красный Яр, R–W |
||||||||
|
n |
Lim |
M |
n |
Lim |
M |
n |
Lim |
M |
n |
Lim |
M |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
Наибольшая длина |
2 |
71,8–77,0 |
74,40 |
Ph 9 |
alanx I (пер 71,0–87,0 |
едние) 78,07 |
115 |
70,0–93,0 |
80,07 |
40 |
72,5–91,6 |
79,68 |
|
Сагиттальная длина, min |
2 |
63,3–67,2 |
65,25 |
9 |
63,0–76,2 |
69,49 |
120 |
61,0–82,7 |
71,65 |
40 |
63,5–82,0 |
71,32 |
|
Ширина верхнего конца |
2 |
37,2–38,0 |
37,60 |
10 |
39,0–51,5 |
45,44 |
115 |
34,7–52,7 |
45,22 |
40 |
36,5–49,5 |
43,94 |
|
Поперечник верхнего конца |
2 |
40,0–43,8 |
41,90 |
4 |
42,0–50,2 |
45,38 |
114 |
38,5–55,2 |
46,89 |
38 |
38,0–56,0 |
46,23 |
|
Ширина диафиза, min |
2 |
33,0–33,9 |
33,45 |
11 |
34,0–48,0 |
41,08 |
119 |
33,0–51,2 |
41,75 |
40 |
34,0–47,0 |
40,97 |
|
Ширина нижнего конца |
2 |
37,3–38,6 |
37,95 |
9 |
37,8–49,7 |
44,30 |
115 |
35,4–56,0 |
45,13 |
37 |
35,3–49,0 |
43,82 |
|
Поперечник нижнего конца |
2 |
27,4–28,4 |
27,90 |
2 |
27,0–33,0 |
30,00 |
103 |
27,2–39,2 |
32,31 |
34 |
25,5–35,0 |
31,00 |
|
Наибольшая длина |
1 |
– |
49,30 |
Ph 11 |
alanx II (пер 47,8–57,2 |
едние) 52,55 |
142 |
46,0–62,0 |
54,37 |
34 |
46,0–58,8 |
53,34 |
|
Сагиттальная длина, min |
1 |
– |
42,30 |
11 |
40,0–47,0 |
43,67 |
147 |
38,5–50,9 |
44,36 |
34 |
39,5–49,0 |
43,82 |
|
Ширина верхнего конца |
1 |
– |
36,20 |
9 |
41,0–49,4 |
45,10 |
139 |
34,3–52,4 |
46,10 |
31 |
39,3–54,0 |
44,94 |
|
Поперечник верхнего конца |
1 |
– |
37,50 |
4 |
48,3–54,0 |
52,00 |
114 |
35,0–55,5 |
48,07 |
31 |
38,0–55,0 |
46,41 |
|
Ширина диафиза |
1 |
– |
30,70 |
11 |
32,0–42,8 |
36,42 |
142 |
28,5–44,2 |
36,76 |
32 |
31,5–44,8 |
37,21 |
|
Поперечник диафиза, min |
1 |
– |
28,30 |
5 |
30,8–35,5 |
33,66 |
145 |
26,4–39,5 |
33,90 |
34 |
28,7–40,0 |
33,42 |
|
Ширина нижнего конца |
– |
– |
– |
6 |
32,0–42,5 |
38,25 |
135 |
30,7–49,0 |
40,63 |
32 |
33,7–44,8 |
38,98 |
|
Поперечник нижнего конца |
– |
– |
– |
4 |
38,0–42,7 |
41,10 |
138 |
32,0–47,5 |
41,38 |
32 |
35,0–44,3 |
39,76 |
|
Наибольшая длина |
1 |
– |
P 76,00 |
halanx 1 |
III (передни – |
е + зад 85,50 |
ние) 124 |
74,7–113,0 |
95,00 |
45 |
75,0–104,5 |
90,62 |
|
Дорзальная длина |
1 |
– |
61,00 |
3 |
60,7–69,0 |
64,73 |
124 |
61,6–93,0 |
73,85 |
45 |
58,0–80,8 |
70,43 |
|
Высота кости |
1 |
– |
43,50 |
4 |
44,0–56,0 |
49,75 |
131 |
47,2–77,5 |
59,35 |
45 |
45,0–68,3 |
56,21 |
|
Высота суставной поверхности |
1 |
– |
26,70 |
1 |
– |
42,00 |
131 |
36,0–54,7 |
43,57 |
46 |
36,0–51,0 |
42,76 |
|
Ширина суставной поверхности |
1 |
– |
35,70 |
4 |
28,5–34,0 |
31,33 |
136 |
27,3–43,0 |
33,94 |
49 |
27,0–41,5 |
32,12 |
|
Наибольшая длина |
3 |
73,2–76,3 |
74,23 |
P 4 |
halanx I (за 78,0–83,0 |
ние) 80,28 |
94 |
73,7–93,0 |
82,56 |
28 |
74,0–90,0 |
82,04 |
|
Сагиттальная длина, min |
3 |
63,4–69,4 |
65,93 |
4 |
70,7–76,7 |
73,73 |
96 |
64,8–83,7 |
73,82 |
29 |
66,7–88,0 |
72,87 |
|
Ширина верхнего конца |
5 |
32,8–37,1 |
35,08 |
4 |
35,0–39,7 |
38,30 |
89 |
33,0–48,5 |
40,89 |
28 |
33,2–47,0 |
40,39 |
|
Поперечник верхнего конца |
5 |
38,1–44,2 |
40,60 |
2 |
43,5–45,5 |
44,50 |
91 |
38,0–54,0 |
46,30 |
28 |
39,5–52,5 |
46,50 |
|
Ширина диафиза, min |
4 |
29,3–31,6 |
30,33 |
4 |
30,7–37,8 |
34,88 |
93 |
29,0–43,1 |
36,70 |
31 |
30,0–43,0 |
36,59 |
Окончание табл. 8
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
Ширина нижнего конца |
3 |
33,2–37,6 |
35,00 |
4 |
34,3–40,4 |
38,00 |
94 |
31,2–48,1 |
40,32 |
28 |
34,2–44,7 |
40,04 |
|
Поперечник нижнего конца |
3 |
24,0–26,3 |
25,53 |
– |
– |
– |
83 |
25,0–36,7 |
29,67 |
22 |
26,0–35,0 |
29,74 |
|
Латеральная длина |
1 |
– |
81,20 |
86 |
Astragalu 78,2–98,2 |
s 89,90 |
162 |
76,2–104,0 |
88,62 |
77 |
82,0–102,5 |
91,72 |
|
Сагиттальная » |
1 |
– |
64,20 |
95 |
60,0–77,0 |
70,38 |
169 |
58,5–80,0 |
69,24 |
77 |
64,2–79,4 |
72,47 |
|
Медиальная » |
1 |
– |
75,50 |
90 |
72,4–90,0 |
83,31 |
160 |
70,3–92,3 |
82,11 |
76 |
77,4–95,1 |
84,85 |
|
Ширина нижнего конца |
1 |
– |
54,30 |
75 |
50,8–69,4 |
61,01 |
146 |
47,5–68,1 |
57,24 |
76 |
53,2–72,8 |
60,87 |
|
Медиальный поперечник |
1 |
– |
43,30 |
78 |
45,0–59,3 |
52,18 |
112 |
42,5–55,3 |
49,67 |
67 |
46,5–60,0 |
52,32 |
|
Латеральный поперечник |
1 |
– |
44,30 |
88 |
42,5–56,2 |
50,95 |
155 |
41,7–58,0 |
49,08 |
75 |
46,0–57,5 |
51,74 |
|
Наибольшая длина |
4 |
39,5–45,5 |
42,23 |
6 |
Os malleol 43,0–50,2 |
are 45,87 |
6 |
45,8–53,3 |
48,70 |
6 |
46,2–49,0 |
47,25 |
|
Наибольший поперечник |
4 |
19,5–21,5 |
20,80 |
6 |
20,5–28,3 |
24,05 |
5 |
21,2–30,8 |
26,50 |
6 |
21,7–27,7 |
24,62 |
|
Высота |
4 |
28,8–34,3 |
31,15 |
6 |
31,0–41,8 |
35,55 |
– |
– |
– |
– |
– |
– |
|
Длина |
1 |
– |
45,00 |
– |
Os tarsale – |
IV – |
4 |
49,2–54,2 |
50,78 |
8 |
48,0–53,5 |
51,44 |
|
Ширина |
1 |
– |
28,00 |
– |
– |
– |
4 |
31,5–33,7 |
32,75 |
8 |
30,7–34,8 |
33,01 |
|
Высота |
1 |
– |
16,00 |
– |
– |
– |
4 |
14,8–16,0 |
15,63 |
8 |
16,2–22,0 |
18,60 |
|
Ширина нижнего конца |
1 |
– |
66,50 |
19 |
Metatarsa 69,7–88,7 |
le 78,54 |
50 |
67,5–84,0 |
76,36 |
47 |
68,3–86,7 |
77,49 |
|
Поперечник нижнего конца |
1 |
– |
40,70 |
16 |
39,6–49,5 |
45,58 |
44 |
40,7–50,4 |
45,88 |
39 |
42,1–50,9 |
46,45 |
Таблица 9. Размеры зубов нижней челюсти Bison priscus
Сайгак ( Saiga tatarica borealis ). Часть трудноопределимых костей и зубов мелких представителей семейства Bovidae , возможно, относится к дзерену ( Procapra gutturoza ). Остатки этой антилопы, наряду с сайгаком, отмечены в плейстоценовых отложениях расположенной в нескольких десятках километрах пещеры Страшной [Васильев, Зенин, 2009, 2010]. Все единичные достоверно определенные остатки из Чагырской пещеры принадлежат сайгаку В слое 6а/1 найдена первая задняя фаланга с разрушенным кислотной коррозией верхним отделом. Длина ко сти в проекции 45 мм, сагиттальная - 42, ширина диафиза 7,3, ширина нижнего конца 9,8, поперечник -10,2 мм. Два характерного строения обломка третьей фаланги обнаружены в слоях 5 (горизонт 6) и 6в/1 (горизонт 5), два изолированных зуба нижней челюсти - в слое 6а/2. Подавляющее число остатков сайгака (и дзерена?) несет признаки деструкции пищеварительными ферментами.
Сибирский горный козел (Capra sibirica). Это второй по численности остатков вид после бизона. Число костей и зубов, пригодных для измерений, невелико. Длина М3 26,5-30,0 (М 28,63) мм (n = 12), ширина 8,0-11,3 (М 9,51) мм (n = 10). Ширина нижнего конца лучевой кости 40,3 мм, его поперечник -26,7 мм. Центральнокубовидная кость имеет ширину 33,5 мм, поперечник 29,7, ширину верхней суставной поверхности 26,2 мм. Ширина и поперечник нижнего отдела плюсневой кости 35,2 и 24,8 мм. Судя по тому, что большинство зубов и фрагментов костей горного козла имеет признаки кислотной коррозии, их аккумуляция в отложениях Чагырской пещеры происходила почти исключительно за счет пищевой активности пе- щерных гиен и волков. Гиены, очевидно, в основном подбирали трупы павших зверей в окрестностях пещеры, а волки могли использовать также активную охоту на этих полорогих. Наибольший урон от них несут во второй половине зимы самцы старше 4-5 лет. Ослабевших во время гона зверей волки загоняют по снегу, отрезая от скальных отстоев [Егоров, 1955; Гептнер, Насимович, Банников, 1961].
Заключение
Тафоценоз плейстоценовых слоев Чагырской пещеры, как и других пещерных местонахождений Алтая, в значительной степени сформировался за счет пищевой активности крупных хищников. Волки и пещерные гиены почти непрерывно использовали пещеру в качестве убежища или логова для выведения потомства.
Во время накопления слоев 6б и 6в пещера периодически служила стоянкой для палеолитического человека. Основной его добычей были бизоны, главным образом полувзрослые особи или же самки. В результате разделки и поедания частей туш животных в приустьевой части постепенно сформировалось достаточно мощное скопление фрагментарных костных остатков с признаками человеческой деятельности. В виде сходящего на нет шлейфа оно прослеживается также и в глубину пещеры. Охота на бизона, возможно, носила сезонный характер и была приурочена к маршруту его ежегодных миграций по долине Чарыша. Гораздо реже добычей палеолитического человека становились, вероятно, лошадь Оводова, архар и сибирский горный козел, северный олень.
Преобладание в тафоценозе слоя 6 остатков Bison priscus и Equus ovodovi заставляет предполагать господство степных умеренно-аридных ландшафтов. В слое 5 доля остатков бизона сокращается в 5,7 раза, а сибирского горного козла и архара увеличивается в 1,8 раза, что, вероятно, является следствием значительной аридизации климата в это время. Присутствие единичных костей соболя и бобра в слоях 5 и 6 Чагыр-ской пещеры свидетельствует о наличии небольших участков лесной растительности по берегам рек и в горных ущельях.
