Faunal remains from the early complexes of the Sagan-Zaba II multilayered habitation site (9120–7880 cal bp) in the Cis-Baikal: planigraphy, subsistence patterns, and seasonality
 in the Cis-Baikal: planigraphy, subsistence patterns, and seasonality")
Author: Nomokonova T.Y., Goriunova O.I., Novikov A.G., Losey R.J., Weber A.W.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Paleoenvironment, the stone age
Article in issue: 3 т.44, 2016.
Free access
Short address: https://sciup.org/145145271
IDR: 145145271 | DOI: 10.17746/1563-0102.2016.44.3.037-046
Text of the review article Faunal remains from the early complexes of the Sagan-Zaba II multilayered habitation site (9120–7880 cal bp) in the Cis-Baikal: planigraphy, subsistence patterns, and seasonality
The multilayered habitation site Sagan-Zaba II is represented by several complexes characteristic of CisBaikal ancient cultures during the Holocene. The period over which the site was occupied has been determined on the basis of extensive radiocarbon dating, which indicates site use from ~9000 to 900 cal BP (Nomokonova et al., 2013). Various archaeological and faunal materials from the site allow tracing of the special features of the Olkhon regional subsistence development in different chronological intervals. Archaeological complexes of a number of cultural layers have already been introduced into scientific use (Dolganov et al., 2011, 2013; Goriunova et al., 2012; Novikov, Goriunova, Weber, 2014; Nomokonova et al., 2010). However, numerous faunal remains obtained in the course of excavations in the northeastern part of the bay, and also the subsistence aspects, were not considered earlier. This article is the first publication devoted to analysis of faunal remains from early complexes of this part of the Sagan-Zaba II site (layers VII and VI of the 2007–2008 excavation areas), including analysis of the species composition, radiocarbon dating of animal bones, planigraphy of their finding in the layers, selection of animals, and seasonality of this site.
The site is situated in the Sagan-Zaba bay, on the western shore of Lake Baikal (Fig. 1), 154 km to the northeast of Irkutsk and 13.5 km to the southeast of the Elantsy settlement (Olkhonsky District of the
Irkutsk Region). It was discovered in 1972 by a team of the North Asian Expedition of the Institute of History, Philology and Philosophy of the SB USSR Academy of Sciences, headed by A.P. Okladnikov. Excavations were carried out by the same team under the supervision of A.P. Okladnikov and I.V. Aseyev in 1974–1975. Five cultural layers were distinguished (Okladnikov, 1975; Aseyev, 2003: 51). Excavations aimed at complex multidisciplinary studies of the site were resumed in 2006–2008 by the Sagan-Zaba team of the Russian-Canadian Archaeological Expedition (Irkutsk Laboratory of Archaeology and Paleoecology of the Institute of Archaeology and Ethnography SB RAS, and Department of Anthropology of the University of Alberta, Edmonton, Canada) (Goriunova, Novikov, Vorobieva et al., 2007; Goriunova, Novikov, Weber et al., 2008). The studies conducted in the northeastern part of the bay (the most promising for excavations) have resulted in 11 cultural layers being distinguished (from VII to I, taking into account that some of them are subdivided into lower and upper).
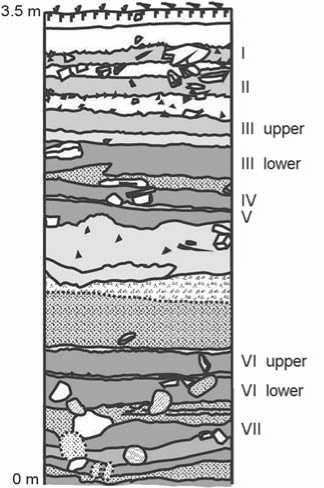
The stratigraphic sequence in the excavation area is a series of humic sandy loam soils separated by interlayers of light rubbly sandy loam and coarse deposits of proluvial and diluvial genesis (Fig. 2) (Goriunova, Novikov, Weber et al., 2008; Vorobyeva, 2010). Complexes VII and VI of the lower and upper cultural horizons are stratigraphically tied to the grayish-brown (up to brown) and dark grayish-brown layers of humic sandy loam 0.12–0.18 m thick. Their division is well defined at the front wall of the
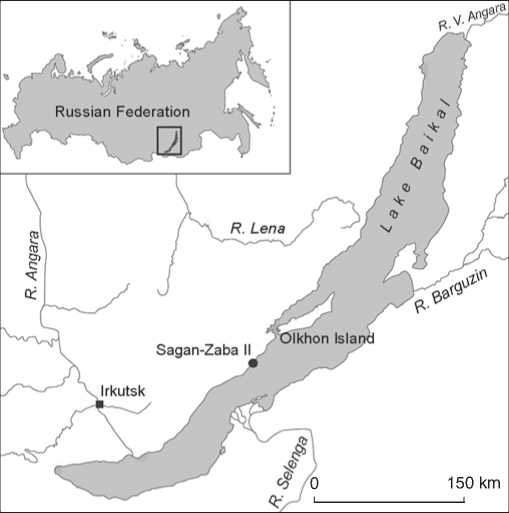
Fig. 1. Schematic map showing location of the Sagan-Zaba II site on the shore of Lake Baikal.
0 1 m
Fig. 2. Stratigraphic sequence of the northwestern wall of trench 4c (sq. 4a).
trench (depthward into abrasion scarp); near the coastal cliff, the layer’s thickness reduces to 0.05 m (Fig. 3).
The technique of field studies included the unearthing of trenches layer by layer; thorough screening of the sediments from all cultural deposits through a 3 mm mesh sieve; recording of materials using an electronic theodolite by threedimensional indicators; detailed stratigraphic study of sequences; and selection of samples for study by natural science methods.
Radiocarbon dating of animal bones
The chronology of lower and upper layers VII and VI of Sagan-Zaba II site has been determined by means of 16 radiocarbon AMS dates (Table 1), calibrated using the Calib 7.0.1 software program (Stuiver, Reimer, 1993) and using the IntCal13 database (Reimer et al., 2013). The dates have been obtained from bones of ungulates and Baikal seals selected in equal quantities from each analyzed layer, and presented on a calibrated curve at 95 % probability. In previous publications, we
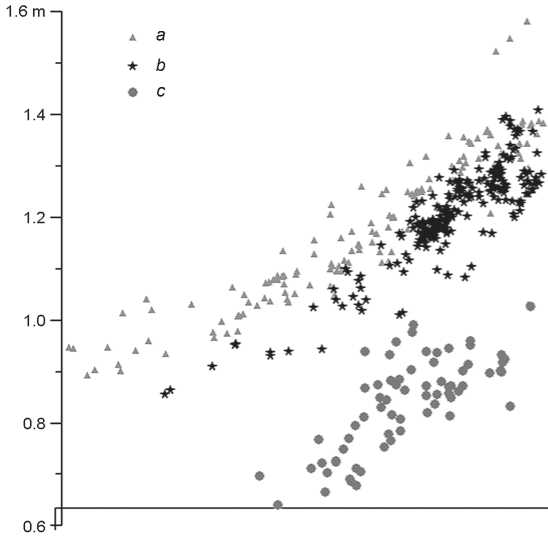
Fig. 3. Two-dimensional vertical sequence of trench 4c with the density of archaeological material found in layers VII and VI. a – upper layer VI; b – lower layer VI; c – layer VII.
Table 1 . Results of radiocarbon dating of animal bones
Considering that several radiocarbon dates have been obtained for each layer, it became possible to use an additional function available in the BCAL software program (Buck, Christen, James, 1999) under the name of ETI (elapsed time intervals). It allows a rough estimate of the length of time represented by the obtained set of dates, to make an assumption about the time period to which these dates pertain and when the site could have been used. Table 1 gives such indicators calculated for the dates obtained from bones of ungulates and Baikal seals from each of the analyzed layers. These demonstrate that, probably, the time periods when these animals were used at the site are more or less equal, except for ungulates whose remains have been found in layer VI, where this indicator is smaller (up to 80 years).
Context of finding faunal remains
Faunal remains in layers VII and VI of the Sagan-Zaba II site were mainly localized as concentrations near the areas of hearths, fireplaces, and stone structures. For example, one fireplace and six stone features, occupying a considerable part of the excavated area, have been recorded in layer VII (Fig. 5). Apparently, all of them were used as hearth-constructions. These structures with oval or circular shape are either continuous or composed of separate stones. The constructions have dimensions of 0.92–0.34 × 0.71–0.28 m. The thickness of ash pits is minor. The archaeological material of layer VII was located in several concentrations in the area of hearths.
OxA22411_Phoca
OxA22387_Cervus
OxA22419_Phoca
OxA22363_Ca pre о lus
OxA22412_Phoca
OxA22364_Ca pre о lus
OxA22358_Artio da ctyla
OxA22421_Phoca
OxA20578_Phoca
OxA20580_Phoca
OxA22420_Phoca
OxA20579_Phoca
OxA22357_Artio da ctyla
OxA22390_Artio da ctyla
OxA22356_Artio da ctyla
OxA22374_Cervidae
। . . .
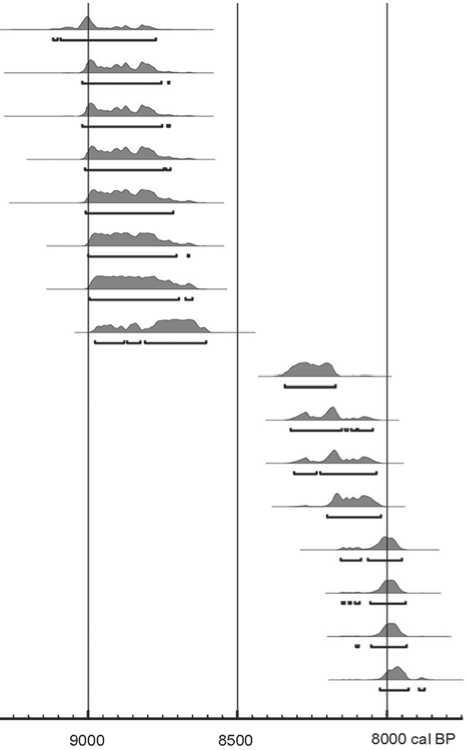
Fig. 4. Radiocarbon dates obtained from animal bones from layers VII and VI.
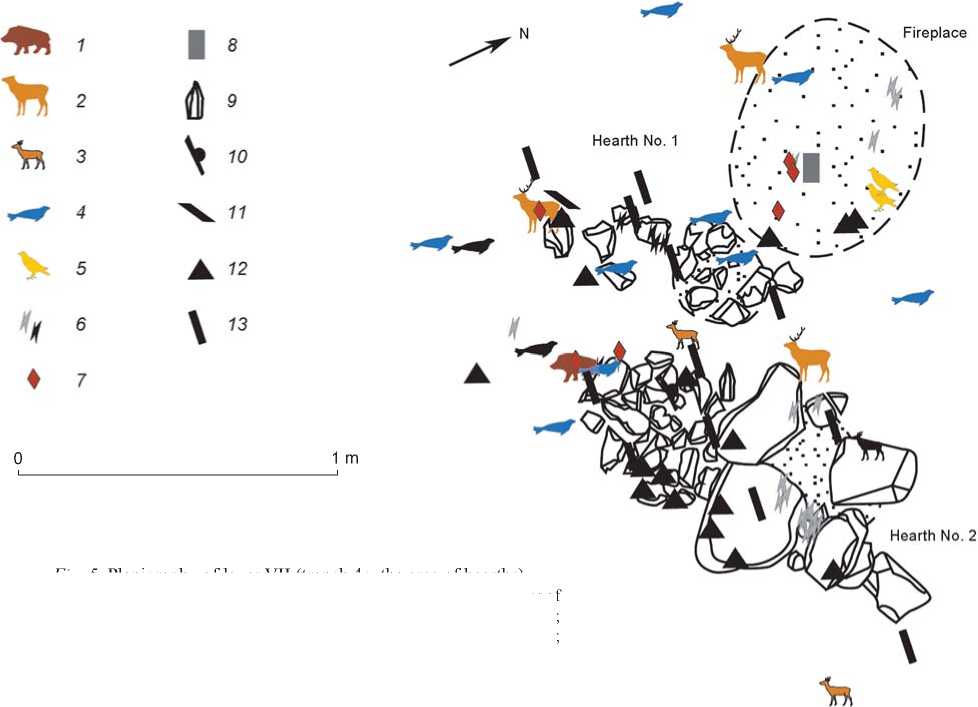
Fig. 5. Planigraphy of layer VII (trench 4c, the area of hearths).
1 – boar bones; 2 – bones of large cervids; 3 – bones of small cervids; 4 – bones of Baikal seals; 5 – bones of birds; 6 – burned indeterminate bones; 7 – bone tools; 8 – an abrader; 9 – a core; 10 – an end-scraper; 11 – a burin; 12 – a flake, spall; 13 – a prismatic blade.
These are mainly faunal remains. Together with these, stone and bone items were also present, including prismatic blades, a cutting tool, an adze, burins, a reamer, inserts and pieces of bone parts of composite tools, a harpoon, and fragments of split boar-canine (Fig. 6, 3–7 ). The compositions of these concentrations make it possible to define them as domestic waste deposits.
In general, the planigraphic placement of faunal remains in lower and upper layers VI confirms the context of the finds near hearths and stone structures. In contrast to the previous layer, pottery fragments (from more than 17 vessels) have been recorded in these complexes. The archaeological material of lower layer VI was concentrated in three accumulations, similar in their composition and typological set of goods (Goriunova et al., 2012). All of them were grouped near the hearth (eastward and westward of it). The semi-circular hearth stone feature has an oval shape, 1.1 × 0.8 m in size. The thickness of the ash pit is small. The accumulations are dominated by faunal remains; also, potsherds of several vessels, prismatic blades, inserts, burins, the shank of a composite fishhook, fragments of bone needle (Fig. 6, 10 ) and shell beads were discovered. Ashy soil is noted at the location of one accumulation.
In upper layer VI, seven stone structures are noted that, possibly, were used as hearths (Ibid). Semi-circular and continuous oval structures of 1.6–0.8 × 1.2–0.5 m in size were encountered. Degraded ash pits have been recorded in some of them. Faunal remains, potsherds from different vessels, prismatic blades, and inserts have been discovered near the hearths. The main part of the archaeological material of upper layer VI is confined to the concentration on the coastal portion of the excavation area. Soil at this place is ashy. The accumulation also included faunal remains, potsherds from different vessels, stone tools (prismatic blades, inserts, a burin, an adzelike tool), a harpoon, and a bone part of a composite tool (Fig. 6, 11 , 12 ). The set and typology of ceramic and stone tools found in lower and upper layers VI are similar, which suggests the formation of complexes within the common cultural tradition, in the same chronological interval (Ibid).
The number of hearths discovered in layers VII and VI, and their physical proximity, indicate that the system of stone structures reflects several subsequent occupations in various seasons, while the relatively minor thickness of ash pits shows that the occupations were short-term.
Judging by the occurrence of faunal remains in hearths and fireplaces as well as near them, carcasses
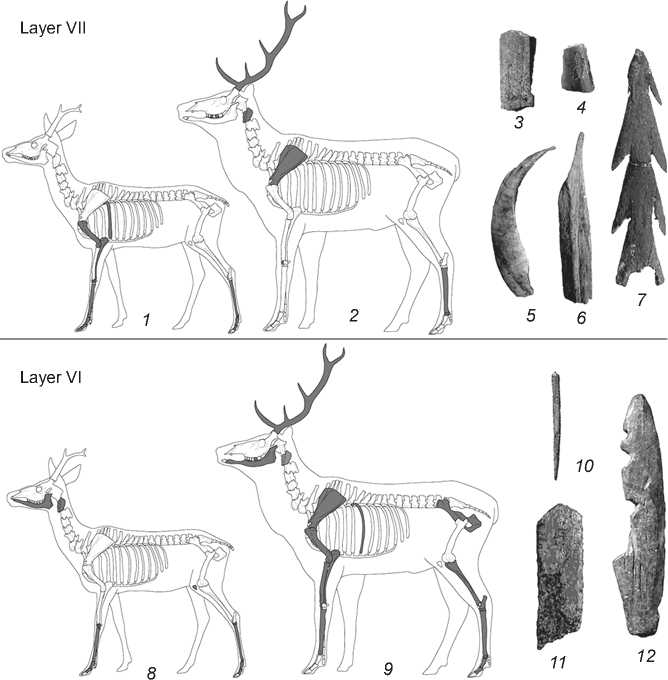
Fig. 6. Skeletal elements of ungulates (marked by black color) and bone tools.
might have been processed and partially consumed directly at the site. This is confirmed by traces on some bones. In materials of layer VII, bones with butchering and gnawing marks, burned bones, and bone tools total 299 specimens, or 28 % of the total quantity of faunal remains from this layer. They belong mainly to mammals. Burned bones, including 15 belonging to Baikal seals, several to ungulates, and one to a bird, prevail (293 total specimens were burnt), which can be immediately associated with hearths and fireplaces. Bones with traces of being worked and bones in the form of tools are represented by 10 specimens. Gnawing marks left by carnivores have been discovered on one long bone.
In the assemblages of layer VI, bones with modification traces are represented by 64 specimens, which comes to 5 % of the total quantity of faunal remains from this layer. These are also dominated by burned bones (53 specimens, including 15 from Baikal seals). Cut marks have been found on a Baikal seal tibia, a red deer humerus, an ungulate rib, and on an indeterminate fragment of mammal bone. In addition, five bones, retouched and in the form of tools, and two bones with carnivore gnawing marks have been found. The presence of the latter may indicate that animal bones at the site were also eaten by carnivores; but, since these finds are single, such actions can be considered as minimal.
Species composition of animals
Faunal materials found as a result of excavations of layers VII and VI are represented by 2351 specimens (Table 2). Most of these (94 % of the total amount of faunal remains) belong to mammals. Species identification in this category is possible according to the following taxonomic levels (Table 2): species (roe deer, red deer, elk, boar, Baikal seal), family (cervids with subdivision to large and smaller sized animals), order (artiodactyls and carnivores), and class (large and small mammals). Other fauna are represented by 17 bird bones and 112 fish remains, as well as by several fragments of shells (Table 2).
In the remains of birds, postcranial elements of skeletons of at least two cormorants, a coracoid bone of raven, a humerus and a coracoid bone of bird belonging to the genus of sea eagles, a phalanx and a furcula fragment of representative of the duck family, and remains of vertebrae indeterminate above the class level have been found. Several elements of fish skulls were attributed to
Table 2 . Faunal remains
|
Taxon |
Name |
Layer VII |
Layer VI |
Total |
||
|
Number of bones |
Number of animals, min |
Number of bones |
Number of animals, min |
|||
|
Mammalia |
Mammals |
711 |
886 |
1597 |
||
|
Mammalia large |
31 |
28 |
59 |
|||
|
Mammalia medium |
30 |
15 |
45 |
|||
|
Artiodactyla |
Ungulates |
7 |
1 |
1 |
8 |
|
|
Artiodactyla large |
– |
– |
4 |
4 |
||
|
Artiodactyla medium |
5 |
5 |
10 |
|||
|
Cervidae |
Cervids |
– |
– |
4 |
2 |
4 |
|
Cervidae large |
5 |
1 |
11 |
2 |
16 |
|
|
Cervus elaphus |
Red deer |
19 |
1 |
13 |
2 |
32 |
|
Alces alces |
Elk |
– |
– |
1 |
1 |
1 |
|
Cervidae medium |
7 |
1 |
7 |
2 |
14 |
|
|
Capreolus pygargus |
Roe deer |
9 |
2 |
10 |
2 |
19 |
|
Sus scrofa |
Boar |
2 |
1 |
2 |
1 |
4 |
|
Carnivora medium |
Carnivores |
1 |
1 |
1 |
1 |
2 |
|
Phoca sibirica |
Baikal seal |
101 |
5 |
296 |
9 |
397 |
|
Aves |
Birds |
3 |
– |
– |
3 |
|
|
Anatidae |
Duck family |
2 |
1 |
– |
– |
2 |
|
Phalacrocax carbo |
Cormorant |
9 |
2 |
– |
– |
9 |
|
Haliaeetus sp. |
Genus of sea eagles |
1 |
1 |
1 |
1 |
2 |
|
Corvus c.f. corax |
Raven |
– |
– |
1 |
1 |
1 |
|
Pisces unidentified |
Fish |
107 |
1 |
1 |
108 |
|
|
Thymallus articus |
Grailing |
3 |
2 |
– |
– |
3 |
|
Acipenser baeri baic. |
Sturgeon |
– |
– |
1 |
1 |
1 |
|
Mollusca |
Shells |
– |
– |
6 |
6 |
|
|
Unidentified |
4 |
– |
– |
4 |
||
|
Total |
1057 |
1294 |
2351 |
|||
at least two grayling and a sturgeon. Other fish remains, represented by scale, ribs, and skull fragments, are unidentifiable.
Most of the faunal remains (17 % of the total amount of bones, or 75 %, if we consider only fauna that can be determined to the order level) belong to Baikal seals. Remains of this species belong to at least 14 animals. Elements of all skeleton parts are available, without any predominance of bones from particular parts of animal carcasses. The second most significant group consists of ungulates: 4.8 % of the total amount of faunal remains, or 21 %, if we consider only fauna that can be identified to the order level. Here, the bones of roe deer and red deer prevail; also, a fragment of an elk radius and remains of boar (a skull fragment, the second phalanx, and a split canine) are noted.
Large and small ungulates are represented by elements of various skeleton sections, though mainly by bones of limbs and head with fragments of antlers (Fig. 6, 1 , 2 , 8 , 9 ). It remains unclear to what extent this situation reflects the selection of certain parts of the carcasses of these animals; since, among mammal remains, elements of other parts if the skeleton are also encountered—for example, fragments of ribs and vertebrae, whose fragmentary nature makes identification of species difficult.
Determination of age and sex of animals
Owing to the fact that bones and teeth of Baikal seals and ungulates prevail among the faunal remains at the
Sagan-Zaba II site, additional studies were carried out to determine the sex and age of some of these animals.
The age of Baikal seals was determined by two methods. The first method is based on assignment of individual skeletal elements to generalized age categories according to the degree of fusion of proximal and distal epiphyses on the bones (mainly, on limbs) (Storå, 2000), while the second relies on calculation of annual structures in dentine from thin-sections of teeth. The latter was developed and applied earlier to analyzing teeth of Baikal seals from a number of multilayered sites in the Cis-Baikal region (Weber, Goriunova, Konopatskii, 1993; Weber et al., 1998). In contrast to the previous studies, final calculations were performed taking into account a minimal number of Baikal seals to preclude the repeated use of the same animals in interpretation of results.
Judging by the degree of epiphyses fusion, seals are represented by remains of at least one animal under one year of age and two mature animals (young and middleaged) in layer VII; and by two animals under one year of age, one immature, and three middle-aged animals in layer VI. Analysis of dentine-layers from thin-sections of 10 teeth has demonstrated that they belonged to two Baikal seals under one year of age in layer VII; two animals under one year of age, five animals of 1–2 years of age, and one seal of five years of age in layer VI. Generalized data on the age-indicators of Baikal seals are given in Fig. 7.
The relative age of ungulates was determined by the state of their teeth, and the degree of the epiphyses fusion of some long bones. These data have been obtained for four roe deer (Tomé, Vigne, 2003). Remains of two animals older and younger than 1.3 years are represented in layer VII, and those of animals before 12 months and older than 1.3 years in layer VI. Bones of red deer between 2 and 4 years of age have been found in layer VI. Judging by the presence of cervid antlers, it can be assumed that some bone remains belonged to male deer (bucks). A canine of an adult male boar was found in layer VII (determination of sex after: (Hillson, 2005: 131)).
Site seasonality
The seasons of use of the Sagan-Zaba II site were reconstructed on the basis of early complexes, using the above analysis of dentine layers from thin-sections of Baikal seal teeth, and by the presence or absence of those animal species that are only available in the CisBaikal region during the warm months of the year, such as migrating birds.
In general, the results of the analysis of dentine-layers from thin-sections of 10 Baikal seal teeth point to the death of these animals during spring or summer time (from March to September): layer VII—two animals under one year of age were killed in the period from March to June; layer VI—the five-year Baikal seal was hunted during March–June, three animals (1–2 years of age) during May–June, and the remainder (under one year of age and 1 year of age) from July to September (Fig. 7). Thus, these animals were hunted during two seasons: in spring, when they go out onto ice and are found near breathing holes, thus forming gatherings on the disappearing ice; and in summer during the opening period of Lake Baikal, when seals are distributed across the whole lake, and groups of them come ashore (Pastukhov, 1993).
The presence of bones of migrating birds also suggests the use of the site during the warm season. For example, ducks and cormorants, which remains have been found in layer VII, and sea eagles, whose bones have been discovered in both layers under study, might have been hunted near Sagan-Zaba bay from the end of March to the beginning of October during their arrival in the CisBaikal region (Fig. 7). However, it must be borne in mind that some sea eagles stay for the winter in this region
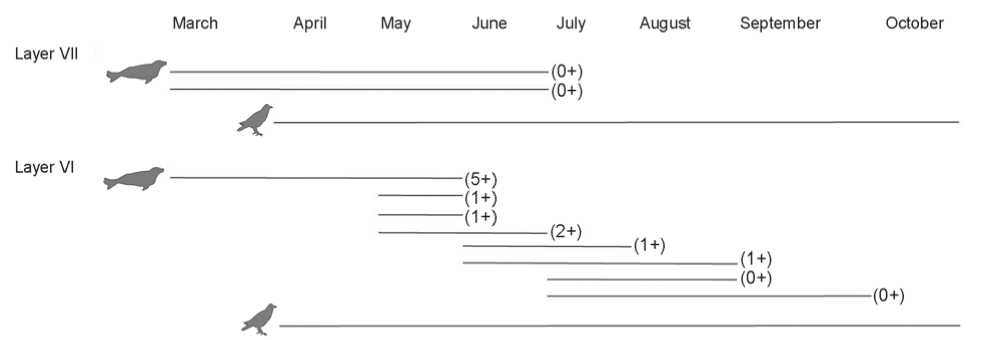
Fig. 7. Seasonality of death of Baikal seals (the age of animal is specified in parentheses) and birds at the Sagan-Zaba II site.
sometimes (Ptitsy Baikala, 2005). Thus, the available data on the studied complexes suggest that the Sagan-Zaba II site was mainly used in the spring and summer months, but other seasons are not excluded.
Conclusions
On the basis of the analysis of a series of radiocarbon dates and calculation of their combined averaging indicators, it was possible to determine the offset between uncalibrated dates obtained from remains of ungulates and Baikal seals (682 years for layer VII and 509 years for layer VI). As a result, the dating of early complexes of the Sagan-Zaba II habitation site was better defined. Taking the correction of dates obtained from Baikal seal bones into account, layer VII is now dated to the period between 9120 and 8610 cal BP, and layer VI to the period between 8340 and 7880 cal BP.
Planigraphic analysis has demonstrated that faunal remains in layers VII and VI were mainly located in concentrations along with stone and bone tools, and also with pottery in the area of the hearths and fireplaces of layer VI. These are probably undifferentiated domestic waste. No independent accumulations of faunal remains have been recorded in the layers. Concentration of the archaeological materials in the area of hearths and fireplaces is typical of all complexes of the Early–Middle Holocene; not only on the shore of Lake Baikal, but in the Cis-Baikal region as a whole.
Most likely, the site featured short-term, seasonal occupations, as indicated by the distribution of hearths and other materials, minor thickness of fireplaces, the absence of specialized production areas, and by relatively small total amount of faunal remains. The results of analysis of dentine layers from thin-sections of Baikal seal teeth, as well as the availability of animal bones from species which are only present during the warm months of the year in the Cis-Baikal region (for example, those of migrating birds), are consistent with the model of seasonal site-use. Thus, it can be assumed that Sagan-Zaba bay was inhabited in the period between 9120 and 7880 cal BP, predominantly during the spring and summer seasons.
To judge by the species composition of the animals, the subsistence activity in this period was based upon the hunting of Baikal seals and ungulates. Bird hunting and fishing also had a definite significance. The large quantities of broken bones (“kitchen waste” and production waste from bone tools) available in the complexes and their localization mainly near hearths and fireplaces, together with the presence of bones with traces of butchering and burned bones, suggest that hunting, butchering, and consumption of some animals took place nearby, or directly in the bay. Notably, antlers and bones of ungulates were used to make tools. The results of the analysis of faunal materials from layers VII and VI of the multilayered habitation site Sagan-Zaba II are indicative of a wide use of natural resources. A noteworthy detail is that no changes in the hunting of animals of one or another species can be observed in the studied complexes throughout the duration of the time-interval under consideration (from 9120 to 7880 cal BP).
