Феномен инсайта и цифровая визуализация активности головного мозга человека

Автор: Шелепин Константин Юрьевич, Соколов Алексей Викторович, Фокин Владимир Александрович, Васильев Петр Павлович, Пронин Сергей Вадимович
Журнал: Психология. Психофизиология @jpps-susu
Рубрика: Психофизиология
Статья в выпуске: 4 т.10, 2017 года.
Бесплатный доступ
Проблема перехода неосознаваемых проявлений нейрофизиологических механизмов в осознаваемые, сопровождающегося осознанным восприятием происходящих событий, является одной из ключевых в когнитивной науке. Методами цифрового синтеза и обработки изображений проведен анализ активности крупномасштабных нейронных сетей головного мозга человека в пороговых условиях распознавания. Показана активность крупномасштабных нейронных сетей в подпороговых, пороговых и надпороговых условиях в случае наблюдения синтезированных динамических изображений. Особое внимание уделено сравнению изображений, отражающих состояние активности мозга (по измерению колебаний BOLD сигнала) в подпороговых, пороговых и надпороговых условиях между собой и по отношению к изображениям распределения активности нейронных сетей в состоянии покоя. Установлено наличие максимального значения BOLD сигнала при достижении порога в одних зонах мозга и снижение активности нейронных сетей в других зонах мозга.
Инсайт, гештальт, неполные изображения, голлин-тест, пороги распознавания, цифровая обработка изображений, функциональная магнито-резонансная томография (фмрт), крупномасштабные нейронные сети
Короткий адрес: https://sciup.org/147160081
IDR: 147160081 | УДК: 159.9:612.82 | DOI: 10.14529/psy170405
The insight phenomenon and the digital visualization of the human brain activity
The problem of the transition of unconscious of neurophysiological mechanisms into conscious ones, is crucial problem for cognitive science. Methods of digital synthesis and image processing have been used to analyze the activity of large-scale neural networks of the human brain in threshold recognition conditions. Activity of large-scale neural networks in subthreshold, threshold and above-threshold conditions in the case of observing synthesized dynamic images is shown. Particular attention is paid to comparing images reflecting the state of brain activity (by measuring the oscillations of the BOLD signal) in subthreshold, threshold and above-threshold conditions among themselves and with respect to images of the distribution of activity of neural networks at rest. The presence of the maximum value of the BOLD signal, when the threshold was reached, was found in some areas of the brain and the activity of neural networks in other areas of the brain decreased.
Текст научной статьи Феномен инсайта и цифровая визуализация активности головного мозга человека
Цель выполненной работы – на основании цифровой обработки изображений распределения значений BOLD-сигнала (Blood-oxygen-level dependent imaging; от англ. Blood oxygenation level dependent contrast [контрастность, зависящая от степени насыщения крови кислородом] или BOLD-вариант функциональной магнитно-резонансной томографии) изучить перераспределение активности крупномасштабных нейронных сетей до, после и при достижении порогов распознавания неполных динамических изображений.
Существующие методы визуализации активности головного мозга человека (Хараузов и др., 2015) и описанные авторами настоящей публикации разработанные ими цифровые методы синтеза неполных изображений (Шелепин, Пронин, 2015; Шелепин, Шелепин, 2015) позволили определить направление решения поставленной задачи. Порог перехода результата неосознаваемых, подсознательных процессов на осознаваемый уровень принятия решений может быть отождествляем с психологическим моментом возникновения озарения. Озарение, или инсайт, предполагает целостное решение задачи – построение геш- тальта. Противоположный ему механизм – это детальный последовательный аналитический способ решения проблем. В контексте вышеизложенного следует предположить, что выбор метода измерения порога восприятия фрагментированной фигуры может быть определен нейрофизиологическими и модельными исследованиям и основан на общих принципах гештальт-психологии: выделение фигуры из фона и доминирование целого над деталями. Такая точка зрения основана на ставшими классическими исследованиях Роберта Гесса и Дэвида Филда, которые показали возможность моделирование инсайта при распознавании фрагментированного объекта в условиях помехи (Field, Hayes, and Hess, 1993; Hess and Field, 1999, рис. 1).
Ориентационная составляющая коллинеарных элементов, образующих рисунок «змея», очень важна для их эксперимента. Как можно увидеть при рассмотрении рис. 1, она является ключевой. Мы применили другой метод.
В описываемом исследовании для моделирования ситуаций формирования Гештальта, построенного на основе неполных изобра- жений, используемых в качестве тестовых, был использован известный Голлин-тест, во многом напоминающий конструкции тестов, предлагавшихся основателями гештальт-психологии (рис. 2).
Выполненное исследование позволяет утверждать, что Голлин-тест как один из наиболее эффективных и изученных методов исследования мозговой деятельности является удобным и надежным способом изучения ме-ханимов возникновения инсайта. Его характеризует легкость измерения порога распознавания задачи, и главное – сопровождение этого порога выраженной эмоциональной реак- цией [Шелепин и др. 2015, Шелепин К., Шелепин Ю., 2015, Harauzov et al., 2016, ]. Ранее нами было показано, что порог распознавания при первом предъявлении стимула в среднем равен 20 %. Но эта характеристика меняется и зависит от сложности рисунка, меры знакомства испытуемого с данным рисунком и с методом исследования, от состояния испытуемого и его возраста. Для применения Голлин-теста как метода, имитирующего инсайт, важно то, что сам момент распознавания сопровождается выраженной осознаваемой эмоциональной реакцией.
Процесс достижения инсайта протекает

Рис. 1. Рисунок «змея» из коллинеарных элементов по ориентации и элементов, ортогональных к оси изображения змеи (по данным Field, Hayes, and Hess, 1993; Hess and Field, 1999)

Рис. 2. Изображения последовательного накопления полноты представления контура фигуры из Голлин-теста при 10, 20 и 100%-ном заполнении контура
неосознаваемо. Наблюдатель видит фрагменты, ожидает увидеть объект, но не может угадать, что он из себя представляет. При этом не происходит построения аналитических схем и не выдвигаются заранее какие-либо гипотезы. Достижение порога распознавания происходит неожиданно для наблюдателя и вызывает у него зарегистрированную нами эмоциональную реакцию. Полученный нами результат позволяет утверждать, что создан новый инструмент исследования нейрофизиологических механизмов инсайта.
Его основу составляют изменения BOLD сигнала (Blood-oxygen-level dependent imaging), полученного на основании данных функциональной магнитно-резонансной томографии (фМРТ) суммарной активности мозга и, в частности, зон, входящих в крупномасштабные нейронные сети, позволяющие определить пороги распознавания динамических изображений фрагментированных объектов.
Методы исследования
В качестве источника динамических изображений выбран компьютеризированный Гол-лин-тест, представляющий собой базу рисунков, которые формируются из постепенно нарастающих по своей полноте заполнения контуров из числа случайно возникающих фрагментов [Foreman. Hemmings, 1987; Шелепин и др., 2015].
Для измерения состояния мозга в момент возникновения инсайта использовали процедуры фМРТ, которые позволяли определять активацию различных областей мозга, установленную на основании оценки изменения локального кровотока в этих областях.
В качестве стимулов был использован динамический сигнал: постепенное нарастание во времени степени заполнения контура различных контурных изображений живых и неживых объектов. Выбор этого теста в качестве модели вызова феномена инсайта был обусловлен возможностью контролировать подпороговые, пороговые и надпороговые состояния мозга в процессе восприятия, стимула, структурированного по мере увеличения удельной доли величины заполнения контура изображения. Контроль состояния в период до возникновения порога, в момент порога и после осуществлялся путем регистрации электроэнцефалографических данных (ЭЭГ), миографии лицевых мышц, измерения размера зрачка и микротремора мышц глазного яблока.
В задаче распознавания стимулов при предъявлении нарастающего числа элементов контура проводил и исследование карты активации распределения мозговой активности (по результатам анализа BOLD-сигнала) в момент предъявления динамического стимула относительно распределенной активности нейронной сети и относительно активности состояний, рассматриваемых применительно характеристик покоя в рамках базового режима мозга (REST). Сравнение осуществлялось визуально и на основании существующих стандартных методов обработки с помощью основанной на высокоуровневом языке интерактивная среда для программирования, численных расчетов и визуализации результатов Matlab и универсального 12канального конвертора сигнала DMX-512 в сигналы напряжения формы DC / ШИМ (драйвер SPM-12) для обработки изображений «виртуальных срезов мозга», полученных по данным фМРТ.
Результаты исследования
Проводилось сравнение изображений, отражающих активность разных зон мозга, по следующим параметрам:
-
а) относительно состояния покоя; б) в момент достижения порога распознавания; в) относительно допорогового состояния головного мозга; г) относительно послепоро-гового состояния мозга.
Были получены статистически значимые и видимые «на глаз» различия максимальной активности головного мозга в момент достижения порога различения (достижения «инсайта»). На графиках такая картина проявлялась в виде пика на картах визуализации активности мозга. Появление такого пика на графиках соответствовало моменту распознавания. Характерно, что последующее увеличение четкости и структурированности представленного контура сопровождается не увеличением, а уменьшением активности головного мозга (рис. 3–6).
Важно отметить, что в момент достижения порога различения наблюдается максимум общего числа проактивированных воксе-лей фМРТ-срезов. Кроме того, наблюдаются локальные особенности уровня активации, которые значительно ниже «покоя» до стимула представлены в одних зонах (определяемых по Бродману) и существенно выше – в других цитоархитектонических зонах.
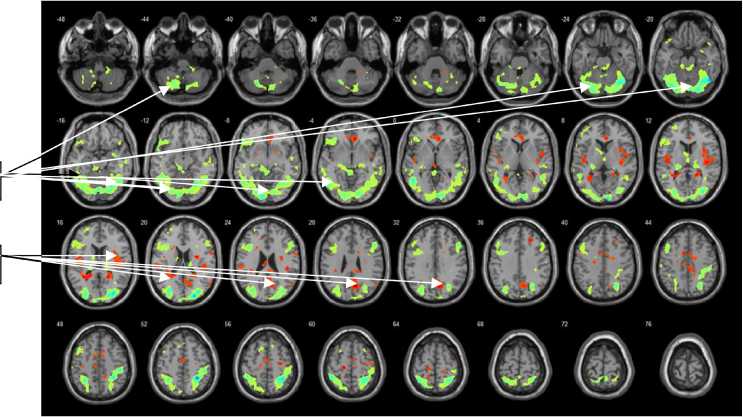
Рис. 3. Усредненный рисунок уровня активации для группы испытуемых численностью 22 человек (FDR-corrected, p = 0,01), обследованных в задачах на распознавание неполно представленного (10 %) изображения контура объекта
Примечание.
Зоны мозга, окрашенные в зеленый цвет (1), – результат распределения активности в головном мозге при вычитании показателей состояния « до инсайта» минус показатели состояния «покой» при 10%-ном заполнении контура в предъявляемом стимуле.
Зоны мозга, окрашенные в красный цвет (2), – результат распределения активности в головном мозге при вычитании показателей состояния «покой» минус показатели состояния « до инсайта» при 10%-ном заполнении контура в предъявляемом стимуле.
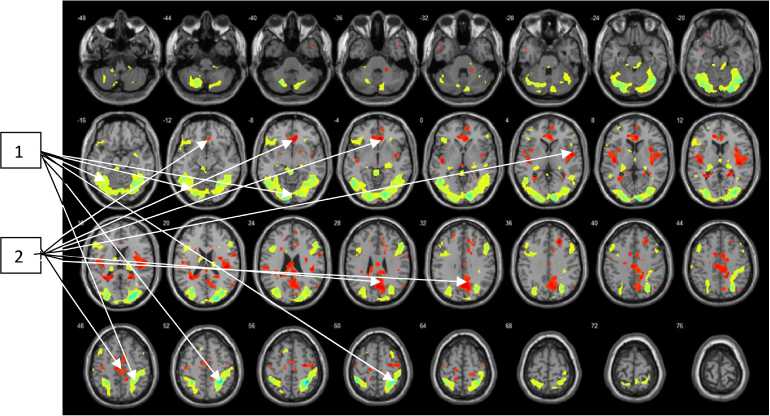
Рис. 4. Усредненный рисунок уровня активации для группы испытуемых численностью 22 человек (FDR-corrected, p = 0,01), исследованных в задачах на распознавание неполно представленного (20 %) изображения контура объекта
Примечание.
Зоны мозга, окрашенные в зеленый цвет (1), – результат распределения активности в головном мозге при вычитании показателей состояния «инсайта » минус показатели состояния «покой» при 20%-ном заполнении контура в предъявляемом стимуле.
Зоны мозга, окрашенные в красный цвет (2), – результат распределения активности в головном мозге при вычитании показателей состояния «покой» минус показатели состояния « инсайта » при 20%-ном заполнении контура в предъявляемом стимуле.

Рис. 5. Усредненный рисунок уровня активации для группы испытуемых численностью 22 человек (FDR-corrected, p = 0,01) в задачах на распознавание неполно представленного (60 %) изображения контура объекта
Примечание.
Зоны мозга, окрашенные в зеленый цвет (1), – результат распределения активности в головном мозге при вычитании показателей состояния « после инсайта» минус показатели состояния «покой» при 60%-ном заполнении контура.
Зоны мозга, окрашенные в красный цвет (2), – результат распределения активности в головном мозге при вычитании показателей состояния «покой» минус показатели состояни-ия « после инсайта» при 60%-ном заполнении контура в предъявляемом стимуле.
А Количество активированных Б Количество активированных векселей векселей
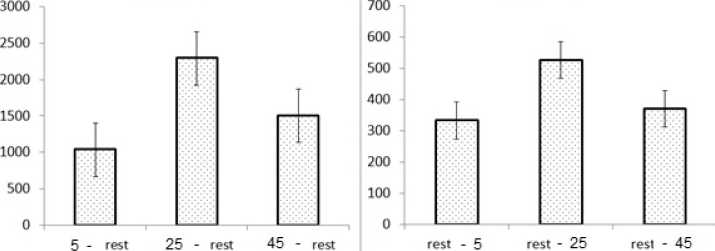
Рис. 6. Значения среднего арифметического количества активированных вокселей всего головного мозга при различных типах вычитания (с указанием стандартной ошибки среднего значения, SE) в зависимости от времени от предъявления стимула до появления инсайта, во время инсайта и после достижения инсайта:
А и Б – различные варианты вычитания характеристик («предъявление стимула» минус «покой» и «покой» минус «предъявление стимула»)
Изучение активности крупномасштабной префронтальной коры (зоны по Бродману BA9, BA10, BA11), височно-теменной коры (BA39, BA40, BA22), лимбической коры (BA23, BA24, BA29, BA30, BA31) задневисочной (BA 37) и затылочной коры (BA 17, BA 18, BA19) и подкорковых ядер (инсулы, амигдалы и др.) показало наличие относительно устойчивых взаимодействий.
Оппонентные взаимодействия, например, BA17 и BA37 или BA9 и BA10, в затылочных, височных и, главное, во фронтальных областях головного мозга, отражают многоуровневый процесс принятия решений по конфликтующим оценкам, предсказанным в теоретических исследования (Saaty, 2008) и реализованных в оппонентных структурах головного мозга человека. Установлен максимум актив- ности одних зон в момент достижения порогового уровня предъявления контура и его снижение после достижения порогового уровня. В оппонентных зонах максимум активности в одной и минимумом в другой зоне совпадает с порогом распознавания (рис. 7).
Обеспечение баланса критериев при принятии решения осуществляется путем перераспределения активности между областями головного мозга. Структуры мозга, обеспечивающие важнейшие когнитивные и непременно сопровождающие их эмоциональные процессы, взаимодействуют по принципу синхронности и оппонентности, т. е. осуществляют решения по множественным критериям и по конфликтующим оценкам (Saaty, 2008). На полученных при фМРТ «срезах мозга» в момент достижения порога, показатели активации существенно больше по сравнению с аналогичными данными в состоянии покоя и со значением активации при подпороговой и надпороговой стимуляции.
Обсуждение
На период подпорогового предъявления стимула приходится первый известный подъем увеличения BOLD-сигнала (на 4–10 секунде от начала предъявления). Этот подъем активности не связан с порогом распознавания и развивается всегда в одно и то же время при любой стимуляции. Для большинства зон (кроме зоны BA17) в этот момент характерна активация. Для того чтобы дифференцировать вызывающий интерес процесс и наступающий сразу после начала предъявления любого стимула известный подъем активности, были проведены отдельные серии исследования, в которых варьировалось время предъявления фрагментов и, тем самым, наступление времени порога распознавания. Вызывающая интерес динамика активности отслеживала динамику изменения тестового сигнала во времени. Так, в случае 20%-ного уровня заполнения контура соответствующий порогу распознавания эффект мог наступать приблизи-

Рис. 7. Изменение во времени BOLD-сигнала в оппонентных зонах мозга префронтальной коры (BA9, BA10) и в зонах BA37 и прекуниус
Примечание. Максимум расхождения значим и совпадает с моментом возникновения «инсайта» или с порогом распознавания и максимумом проявлений эмоциональной реакции испытуемого.
тельно через 20–30 секунд после начала предъявления.
Фактически впервые были рассмотрены основные нейрофизиологические механизмы порога воприятия стимула по данным изменения BOLD-сигнала, установленным на основании разницы значений этого сигнала в покое, при подпороговой стимуляции, при стимуляции в момент достижения порога и при стимуляции после достижения порога в ответ на изменяющейся во времени сигнал. В частности, в момент надпорогового предъявления стимула был установлен спад активности. При распознавании Голлин-теста активность зоны ВА37 правого полушария значимо превышает активность ВА37 левого полушария.
Следует отметить, что по данным Роберта Гесса и Дэвида Филда, полученным на основе анализа собственных результатов моделирования горизонтальных взаимодействий в первичной зрительной коре, известные ранее представления основателей гештальт-психологии о свойствах целостного объекта, построенного из фрагментов, подтвердили возможность восстановления восприятия всего контура за счет внутрикорковых горизонтальных связей [Field, Hayes and Hess,1993; Hess and Field, 1999].
Однако предложенная ими схема вызывает возражения в отношении зоны ВА17. Горизонтальные связи могут развиваться в пределах стриарной коры ВА17 лишь на короткие дистанции. Стимулы, примененные в виде фигуры «змеи», представляются более оптимальными, так как при их предъявлении один фрагмент следует за другим. В то же время, если элементы фигуры змеи не коллинеарные, то зрительной системе требуется использование другого механизма. Таким другим механизмом может служить опция, связанная с зоной, обеспечивающей целостное восприятие (ВА37), что и получило уже подтверждение в дальнейших наших исследованиях.
Заключение и выводы
В описываемом исследовании впервые показана динамика изменения сигнала от подпорогового до надпорогового уровня. Измерение порогов распознавания методами цифровой обработки откликов головного мозга позволит перейти к целенаправленному моделированию искусственных нейронных сетей. При этом порог распознавания стимула сопровождается максимумом активности все- го мозга, превышающем не только уровень «покоя» при отсутствии стимулов, но и так же подпороговый и надпороговый уровень, когда объект был виден ясно и четко. Перестройка активности естественной нейронной сети обусловлена перераспределением активности оп-понентных корковых областей мозга и подкорковых ядер, что было установлено на основании изменения BOLD-сигнала. Установлено, что изменение BOLD-сигнала во времени коррелирует с изменением спектра ЭЭГ: снижением альфа- и повышением гамма-ритма. Этот результат подтверждается данными, представленными другими исследователями (Kounios, Beeman, 2014). Максимум нейронной активности в момент порогового распознавания формы фигуры сопровождается выраженной эмоциональной реакцией, которую регистрировали на основании оценки устных ответов испытуемых и результатов объективных измерений колебаний размеров диаметра зрачков, проявлений микротремора глазных яблок и напряжением мимических мышц, сопровождающих любую эмоциональную реакцию (Шелепин и др., 2015).
Список литературы Феномен инсайта и цифровая визуализация активности головного мозга человека
- Хараузов, А.К. Восприятие изображений в задачах зрительного поиска в условиях динамической помехи/А.К. Хараузов, П.П. Васильев, Ю.Е. Шелепин и др.//Оптический журнал. -2015. -Т. 82, № 5. -С. 42-55.
- Шелепин, К.Ю. Распознавание фрагментированных изображений и возникновение «инсайта»/К.Ю. Шелепин, С.В. Пронин, Ю.Е. Шелепин//Оптический журнал. -2015. -Т. 82, № 10. -P. 70-78.
- Шелепин, К.Ю. Нейрофизиология «инсайта»/К.Ю. Шелепин, Ю.Е. Шелепин//Петербургский психологический журнал. -2015. -№ 11. -С. 19-38.
- Field, D.J. Contour integration by the human visual system: evidence for a local «association field»/D.J. Field, A. Hayes, R.F. Hess//Vision Res. -1993. -Jan; 33(2). -P. 173-193. PMID: 8447091
- Foreman, N. The Gollin incomplete figures test: a flexible, computerised version/N. Foreman, R. Hemmings//Perception. -1987. -February 16(4). -P. 543-548. •
- Harauzov, A.K. The time course of pattern discrimination in the human brain/A.K. Harauzov, Y.E. Shelepin, Y.A. Noskov et al.//Vision Research. -2016. -Vol. 125. -P. 55-63.
- Hess, R. Integration of contours: new Insights/R. Hess, D. Field//Trends in Cognitive Sciences. -1999. -Vol. 3, No. 12. -P. 480-486.
- Saaty, T.L. The analytic hierarchy and analytic network measurement processes: Applications to decisions under Risk/T.L. Saaty//European Journal of Pure and Applied Mathematics. -2008. -Vol. 1 (1). -P. 122-196.
- Kounios, J. The Cognitive Neuroscience of Insight»/J. Kounios, M. Beeman//Annu. Rev. Psychol. -2014. -Vol. 65. -P. 71-93.
