Фитоценологическое изучение и оценка ценопопуляций некоторых видов Euphorbiaceae, распространенных в Нахичевани

Автор: Худавердиева С.Ф.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 12 т.8, 2022 года.
Бесплатный доступ
На территории Нахичевани (Азербайджан) в 2019-2021 годах проводились исследования с целью изучения группировок, образованных видами семейства Euphorbiaceae по типу растительности, поиска редких и исчезающих видов и оценки их на уровне ценопопуляций. В ходе исследований использовались классические геоботанические и фитоценологические методы. В результате исследований установлено, что в местной флоре 35 видов семейства Euphorbiaceae распространены в 8 типах растительности, 29 классах формаций, 87 формациях и 128 ассоциациях. За период исследований выявлено 25 новых фитоценозов в Нахичевани. Дана фитоценологическая классификация группировок, в состав которых входят виды рода Andrachne , находящиеся под угрозой исчезновения, исследован их онтогенетический статус. Установлено, что все ценопопуляции относятся к ювенильным типам (так как ∆ - 0,15-0,34
Молочайные, экология растений, растительные сообщества, продуктивность культур
Короткий адрес: https://sciup.org/14126154
IDR: 14126154 | УДК: 581.552+574.34 | DOI: 10.33619/2414-2948/85/17
Phytocoenological study and assessment of coenopopulations of some species of the Euphorbiaceae distributed in Nakhichevan (Azerbaijan)
On the territory of the Nakhichevan (Azerbaijan) in 2019-2021, studies were carried out to study the groups formed by species of the Euphorbiaceae family according to the type of vegetation, search for rare and endangered species and evaluate them at the level of coenopopulations. The studies used classical geobotanical and phytocoenological methods. As a result of the research, it was found that in the local flora 35 species of the Euphorbiaceae family are distributed in 8 types of vegetation, 29 classes of formations, 87 formations and 128 associations. During the research period, 25 new phytocoenoses of the Nakhichevan were identified. A phytocoenological classification of groups, which include endangered species of the genus Andrachne , is given, and their ontogenetic status is studied. It has been established that all coenopopulations belong to the juvenile type (since ∆ - 0.15-0.34
Текст научной статьи Фитоценологическое изучение и оценка ценопопуляций некоторых видов Euphorbiaceae, распространенных в Нахичевани
Бюллетень науки и практики / Bulletin of Science and Practice
Лекарственные препараты, применяемые при лечении многих заболеваний, синтезируют из биоактивных молекул, выделенных из некоторых видов молочайных в большинстве стран мира [1–3]. В Азербайджане фитохимическое изучение таких лекарственных растений не проводилось до сих пор.
Нахичевань представляет собой типичный горный район с очень разными почвенноклиматическими и ландшафтными характеристиками, что в свою очередь создало возможность для формирования богатого биоразнообразия в этом районе. Ландшафтный покров района включает скалы, каменистые и щебнистые сухие склоны, луга, степи, пустыни и полупустыни, лесной покров, кустарники и пастбища. Хотя на каждой из этих территорий сформировались уникальные флора и растительность, обращает на себя внимание наличие сходных по составу фитоценозов.
В Нахичевани изучение фитоценозов, созданных видами семейства молочайных, выявление полезных свойств и определение путей их использования, охрана и устойчивое использование редких видов является одним из важнейших и хозяйственно важных вопросов. В целом во флоре республики семейство Euphorbiaceae выделено в качестве отдельного объекта исследований, исследования его современного состояния в растительности и проведения исследований в качестве источника лекарственного сырья не проводились. В последние годы некоторые сведения о семействе можно найти в работах отдельных исследователей [4–6]. Отдельные исследования проводились только в направлении флористического изучения семейства [5, 7, 8].
Поэтому оценивали роль важных видов семейства в типах растительности, фитоценологическом статусе и онтогенетическом статусе. В частности, планируется провести фитоценологический анализ видов Andrachne , распространение которых ограничено только территорией Нахичевани, а в пределах ареала являются редкими и исчезающими.
Материалы и методика
Научно-исследовательская работа проводилась в 2019–2021 гг. В качестве объектов были взяты виды рода Andrachne семейства Euphorbiaceae, распространенные во флоре Нахичевани. Фитоценологические исследования проводились следующими методами.
Стадии развития особей растений характеризовались с помощью концепции дискретного представления онтогенеза Т. А. Работнова (1992) и А. А. Уранова (1975) [15, 16]. Описание онтогенеза жизненных форм проводилось на основе диагнозов и ключей онтогенетического состояния растения [2]. Для описания и выделения фаз морфогенеза использовались подходы Т. И. Серебрякова (1954), примененные ранее к другим жизненным формам растения [18].
Изучение корневой системы растения проводили методом сплошной выкопки с использованием бинокулярной лупы [11, 13]. При морфологическом анализе подземных органов основное внимание обращали на способность корня образовывать боковые побеги, а также на наличие главного корня, его длину; длина боковых корней и порядок ветвления; количество и длина дополнительных корней; отмечается направление роста корня (расположение корней в растении). Кроме того, проведен сравнительный и морфологический анализ заново собранного гербарного материала, позволивший выявить взаимосвязь между подземными органами у растения и образованием дополнительных очагов в меристеме растения. Анализ проводился следующим образом. Наличие или отсутствие главного корня растения отмечено у имматурных (im), виргинильных (v), молодых генеративных (g1), средневозрастных (g2) и старых генеративных (g3), субсенильных (ss) и сенильных (s) периодах ценопопуляции.
Полученные результаты были проанализированы с использованием критерия сравнения χ 2.
Предпринята попытка изучить скорость роста побегов и корневых ветвей. Морфологически описаны основные родительские особи. Собранные раметы высаживали рядами размером 1,5×2,0 м с расстоянием между растениями 10 см. Онтогенетическое состояние отмеченных ветвей растения регистрировали в период орошения.
В каждом ряду высаживали по 50 образцов растений. Перед посадкой фиксировали их корневую систему — количество дополнительных побегов и их расположение на разных типах корней, а также количество разрушенных розеток, затем фотографировали подземные органы и только после этого растения высаживали. В течение вегетационного периода наблюдали скорость роста корневых побегов (раметов) растений, новообразованных корневых побег и их онтогенетическое состояние.
В ходе исследований изучались ценопопуляции изучаемых видов в разных фитоценозах. Материалы собирались по общепринятой методике популяционных исследований. Сбор материалов осуществляли на площадках или трансектах, расположенных последовательно и разрозненно. Трансекты закладывались следующим образом; они должны пересекать поле и характеризовать относительно высокую и низкую численность любого выбранного вида. Размер опытных участков, таких как проекция надземной и подземной частей, был больше трех фитогенных участков, определяющих, в частности, другие элементы ценопопуляции [15].
Опытные площади для растений имели размеры 50×50 см, количество площадок 10– 156. Исследуемое растение удаляли с полей и определяли его онтогенетический статус. На основании полученных результатов был составлен спектр онтогенетического статуса (возрастной спектр по А. А. Уранову).
Оценку питательной среды растения проводили по экологической шкале Л. Г. Раменского [17].
В качестве интегральных характеристик демографической структуры растения использовались следующие показатели популяций:
Возрастной индекс ( Δ ) [15]:
∑К и ×Н и
∆= Н и — онтогенетическое положение; Ки — «оценка»; Ни — количество особей; I — состояние популяций; Н — общее количество особей в популяции.
Индекс восстановления [9] (И в ) выражается как отношение прегенеративных особей к сумме прегенеративов и генеративов:
И в
∑ 5 и=3 Н и
∑ 8 и=3Ни
Индекс старения [9]:
И с
∑ 1 и 1 =9 Н и
= 11
∑ и=3 Н и
Индекс эффективности [11, 12]:
∑ни × еи
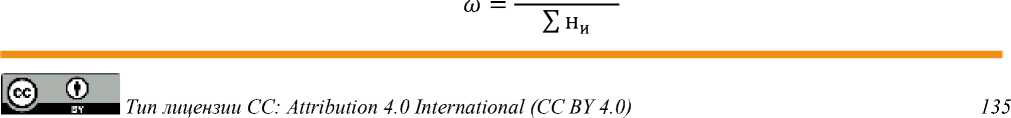
н и — количество растения; и — положение; е и — эффективность растения. Эффективная экологическая плотность популяции (M э ) по [11, 12]:
М э
^Н и X ^
ни — количество растения; и — положение в единичном поле; ωi — энергоэффективность популяции.
Нахождение интервального значения параметров осуществлялось бутстрап-методом.
При этом для оценки плотности ценопопуляций использовали 1n (n+1), ∆, и в , arcsin √ p, p [9].
Сравнение параметров популяций растений в разные годы в пределах одной популяции проводили с помощью t-критерия Стьюдента, представляющего собой поправку Шидака на множественные сравнения.
Тип ценопопуляции определяли с помощью классификации А. А. Уранова по критерию абсолютного максимума и классификации дельта-омега нормальных ценопопуляций Л. А. Живатовского [15, 17].
Результаты и обсуждение
В результате исследований установлено, что во флоре региона распространено 35 видов семейства, входящих в 5 родов. При изучении фитоценозов, образованных видами, принадлежащими к семейству Euphorbiaceae, распространенными на территории Нахичевани, впервые для местной флоры были обнаружены 4 новых вида, принадлежащих к 2 родам: Acalypha australis L., A. hispida Burm. f., Euphorbia marginata Pursh и E. milii Des Moul. Фитоценологическая классификация территории составлена с учетом доминирующих, субдоминантных или составных видов в типах растительности: 8 типов растительности, 29 классов формаций, 87 формаций и 128 ассоциаций. За период исследований выявлено 25 новых фитоценозов растительности Нахичевани.
Определены фитоценозы видов исследуемого семейства, подверженных природным и антропогенным воздействиям, даны соответствующие предложения и рекомендации по их улучшению, повышению продуктивности и охране.
Главную роль в сложности и физиономии растительности играет не обилие отдельных семейств и родов, а обилие и назидательность отдельных видов растений. На основании сказанного были определены доминирующие, эдификаторные и субдоминантные виды местной флоры семейства Euphorbiaceae (Таблица 1).
Таблица 1
ДОМИНИРУЮЩИЕ ВИДЫ СЕМЕЙСТВА Euphorbiaceae ПО РОДАМ
|
Роды |
Количество видов |
% |
Доминирующие виды |
В % от общего числа |
|
Andrachne L. |
4 |
11,43 |
2 |
5,71 |
|
Chrozophora Neck. |
ex 2 |
5,71 |
1 |
2,86 |
|
A. Juss. |
||||
|
Euphorbia L. |
26 |
74,29 |
8 |
22,86 |
|
Acalypha L. |
2 |
5,71 |
— |
— |
|
Ricinus L. |
1 |
2,86 |
— |
— |
|
Всего: |
35 |
100 |
11 |
31,43 |
Из них Euphorbia seguieriana , E. marschalliana, E. iberica, E. humifusa и др. хотя в растительности они представлены одним видом, но превосходят другие виды в образовании групп, таких как виды, отличающиеся своим обилием. Как видно из Таблицы 1, в 5 родах
Бюллетень науки и практики / Bulletin of Science and Practice Т. 8. №12. 2022 насчитывается 11 ценозных почвоукрепляющих видов растений, которые в основном являются доминантными видами, но могут также участвовать в группах в качестве эдификаторов и субдоминантов.
В ходе исследования в растительных фитоценозах определяли численность и стратификацию видов семейства Euphorbiaceae, распространенных на территории Нахичевани (Таблица 2).
Таблица 2
|
ФИТОЦЕНОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ ВИДОВ, ВХОДЯЩИХ В СЕМЕЙСТВО Euphorbiaceae |
||||
|
Название вида |
Обилие |
Высота |
Фенофаза |
Ярус |
|
Andrachne buschiana |
Sos |
15–40 |
V–VI |
II |
|
A. filiformis |
Sp |
30–80 |
V–IX |
I |
|
A. rotundifolia |
Sol |
10–15 |
IV–V |
III |
|
A. telephioides |
Sp |
18–25 |
III |
III |
|
Chrozophora hierosolymitana |
Sol |
7–18 |
IV–V |
III |
|
Ch. tinctoria |
Cop2 |
15–65 |
IV–VII |
II |
|
Ricinus communis |
Sp |
5–10 |
IV–VI |
IV |
|
*Acalypha hispida |
Sp |
180–250 |
V–IX |
I |
|
*A. australis |
Cop2 |
20–34 |
V–IX |
III |
|
Euphorbia arvalis |
Cop2 |
25–70 |
V–VII |
II |
|
E. azerbajdzhanica |
Cop2 |
18–45 |
IV–VI |
II |
|
E. chamaesyce |
Sol |
35-80 |
V–VII |
I |
|
E. condylocarpa |
Cop2 |
40–85 |
V–VII |
I |
|
E. eriophora |
Sol |
60–80 |
VI–IX |
I |
|
E. falcata |
Sp |
20–70 |
VII–IX |
II |
|
E. grossheimii |
Sol |
30–100 |
VI–IX |
I |
|
E. helioscopia |
Cop2 |
75–90 |
VI–VIII |
II |
|
E. heteradena |
Cop2 |
30–35 |
V–VII |
III |
|
E. humifusa |
Cop2 |
8–40 |
V–IX |
II |
|
E. iberica |
Cop3 |
8–40 |
V–IX |
II |
|
E. marschalliana |
Cop3 |
10–40 |
V–VIII |
III |
|
E. orientalis |
Cop2 |
14–22 |
IV–V |
III |
|
E. seguieriana |
Sos |
15–25 |
IV–V |
III |
|
E. szovitsii |
Sp |
6–30 |
IV–V |
III |
|
E. glareosa |
Cop2 |
9–60 |
IV–VI |
II |
|
E. nutans |
Cop2 |
5–30 |
IV–VI |
IV |
|
E. coniosperma |
Sp |
23–30 |
V–VII |
II |
|
E. rhabdotosperma |
Sp |
12–18 |
VI–VIII |
III |
|
E. armena |
Cop2 |
34–40 |
VII–IX |
IV |
|
E. virgata |
Cop3 |
25–40 |
V–VI |
IV |
|
E. granulata |
Cop3 |
30–45 |
VII–VIII |
II |
|
*E. milii var. splendens |
Sp |
120–200 |
V–XII |
I |
|
*E. marginata |
Sp |
65–70 |
V–XII |
II |
|
E. stricta |
Cop3 |
20–70 |
V–VIII |
II |
|
E. woronowii |
Sp |
10–30 |
V–VIII |
III |
|
* — обнаружены впервые для флоры Нахичевани |
||||
В ходе фитоценологических исследований растительные ценозы, образованные видами Andrachne buschiana и A. filiformis , встречающимися в равнинной, горной и высокогорной зонах и распространение которых ограничивается только территорией автономной республики в составе Азербайджана, а также видами A. rotundifolia и A. telephioides , описан род Andrachne , находящийся под угрозой исчезновения (Таблица 2).
Andrachne rotundifolia — редкое растение, самостоятельных растительных сообществ не образует. Однако на равнинах Нахичевани, от предгорий до нижнего горного пояса, в пойме Седарака, на равнинах Бойукдуза, Гюлюстана, Яйчи, Дасты, Ордубада, на высотах 600–1300 (1450 м), встречается индивидуально в составе некоторых фитоценозов горностепной и горной ксерофитной растительности [5].
Растения здесь состоят из ксерофитов, недолговечных эфемеров и эфемероидов из-за неблагоприятных почвенно-климатических условий, особенно малого количества осадков (250–300 мм). В фитоценозах, в которых участвует андрахна, развито около 65–78 растений разных жизненных форм (деревья, кустарники, полукустарнички, многолетние травы и др.). Часто встречаются Artemisietum, Anabasietum, Suaedetum, Stipetum, Salsoletum, Salsoletum-Stiposum, Thymetum, Thymetum-Astragaleto-Salsolosum, Kalidietum, Halostachietum, Halocnemetum, Petrosimonetum-Ephemerosum и другие типичные виды, играющие субдоминантную роль в других фитоценозах: Chenopodium vulvaria L., Astragalus caraganae C. A. Mey., A. szovitsii Fisch. et C. A. Mey., Scutellaria karjaginii Grossh., Stachys fruticulosa M. Bieb., S. inflata Benth., Verbascum saccatum K. Koch, Marrubium nanum Knorring, Salvia limbata C. A. Mey., S. hydrangea DC. ex Benth., Rhinopetalum gibbosum (Boiss.) Losinsk. et Vved., Dichodon cerastoides (L.) Rchb., Oxyria digyna (L.) Hill, Thymus collinus Bieb. и др. встречается. Установлено, что состав фитоценозов формировался при участии 23–45 видов. Поскольку видовой состав указанных фитоценозов разрежен, общее проективное покрытие колеблется в пределах 42–55% (Таблица 3).
Таблица 3
Andrachne rotundifolia (SP I) В БУЮКДУЗСКОМ РЕГИОНЕ КЕНГЕРЛИНСКОГО РАЙОНА
|
Название растений |
Обилие |
Высота (см) |
Фенофаза |
Ярус |
|
Andrachne rotundifolia C. A. Mey. |
1–2 |
15–40 |
цветок-плод |
III |
|
Calligonum aphyllum (Pall.) Gürke |
2–3 |
180–200 |
плод |
I |
|
Chenopodium botrys L. |
3–4 |
25–40 |
плод |
II |
|
Chenopodium vulvaria L. |
2–3 |
15–40 |
цветок-плод |
III |
|
Atriplex tatarica L. |
1–2 |
25–70 |
цветок |
II |
|
Poa bulbosa L. |
2 |
10–20 |
плод |
III |
|
Halocnemum strobilaceum (Pall.) Bieb. |
2–3 |
10–50 |
плод |
II |
|
Koelpinia linearis Pall. |
2 |
15–30 |
цветок |
III |
|
Ceratocarpus arenarius L. |
3 |
5–25 |
плод |
III |
|
Salsola dendroides Pall. |
3 |
80–100 |
цветок |
II |
|
Halostachys caspica (Pall.) C. A. Mey. |
4 |
150–200 |
цветок |
I |
|
Hordeum leporinum Link |
2 |
10–40 |
цветок |
III |
|
Camphorosma lessingii Litv. |
2–3 |
10–35 |
плод |
III |
|
Adonis flammea Jacq. |
2–3 |
10–50 |
плод |
II |
|
Petrosimonia glauca (Pall.) Bunge |
3 |
5–50 |
плод |
III |
|
Eremopyrum triticeum (Gaertn.) Nevski |
2–3 |
10–30 |
плод |
III |
|
Spinacia tetrandra M. Bieb. |
1–2 |
10–40 |
плод |
III |
|
Suaeda microphylla Pall. |
1–2 |
25–75 |
цветок |
II |
|
Tribulus terrestris L. |
2 |
10–60 |
цветок-плод |
II |
Бюллетень науки и практики / Bulletin of Science and Practice Т. 8. №12. 2022
|
Название растений |
Обилие |
Высота (см) |
Фенофаза |
Ярус |
|
Aegilops cylindrica Host |
2–3 |
25–40 |
плод |
III |
|
Kochia prostrata (L.) Schrad. |
4 |
30–60 |
цветок-плод |
III |
|
Kalidium caspicum (L.) Ung.-Sternb. |
4–5 |
10–70 |
цветок-плод |
III |
|
Thesium szowitsii A. DC. |
2 |
20–50 |
цветок |
III |
Andrachne buschiana в отдельных фитоценозах участвует индивидуально. Такие фитоценозы встречаются в горной зоне Нахичевани, на сухих каменистых участках нижнего горного пояса, в растительных группах редкокустарниковых склонов. Основными группировками видов Andrachne buschiana , наблюдаемыми в качестве компонента, являются молочай козий, полынь горная и полынь обыкновенная. Надземные части видов растений здесь не соприкасаются друг с другом. Видовой состав беден и рассеян. В целом в этих ценозах зарегистрировано 19–35 видов. Для этих группировок Centaurea iberica Trevis. ex Spreng., Echinops sphaerocephalus L., Rhamnus pallasii Fisch. et. C. A. Mey., Atraphaxis spinosa L., Cotoneaster melanocarpus Fisch. ex Blytt, Amygdalus fenzliana (Fritsch) Lipsky, Juniperus pygmaea K. Koch, J. polycarpos K. Koch, Ephedra procera Fisch. et. C. A. Mey., Tulipa biflora Pall., Iris lycotis Woronow, Ferula oopoda (Boiss. et Buhse) Boiss., Senecio vernalis Waldst. et Kit., Scabiosa rotata M. Bieb., Ceratocarpus arenarius , Salsola crassa Bieb., Achillea millefolium L., Allochrusa versicolor (Fisch. et C. A. Mey.) Boiss., Capparis herbacea Willd., Artemisia vulgaris L. и др. являются характерными.
Проведена оценка фитоценологического состава Andrachne buschiana в формации Eremurieta persicariae вокруг Дарыдага и сел Диза-Яйджи. Проективное покрытие вида составляет 35%, а общее проективное покрытие 65–70% (Таблица 4).
Таблица 4
ВИДОВОЙ СОСТАВ И СТРУКТУРА ФИТОЦЕНОЗА С УЧАСТИЕМ ВИДОВ Andrachne buschiana (SP II) ВОКРУГ ДАРЫДАГА ДЖУЛЬФИНСКОГО РАЙОНА
|
Название растений |
Обилие |
Высота (см) |
Фенофаза |
Ярус |
|
Andrachne buschiana Pojark. |
1–2 |
15–45 |
цветок-плод |
II |
|
Helichrysum pallasii (Spreng.) Ledeb. |
2–3 |
18–35 |
плод |
II |
|
Koelpinia linearis Pall. |
2 |
15–30 |
плод |
III |
|
Ceratocarpus arenarius L. |
3 |
19–25 |
плод |
III |
|
Hypericum linarioides Bosse |
3 |
80–95 |
цветок- плод |
I |
|
Salsola crassa M. Bieb. |
3–4 |
5–50 |
цветок |
III |
|
Herniaria glabra L. |
3 |
5–15 |
цветок |
III |
|
Spinacia tetrandra M. Bieb. |
1–2 |
10–40 |
плод |
III |
|
Buglossoides arvensis (L.) I. M. Johnst. |
1–2 |
25–75 |
цветок |
II |
|
Aegilops cylindrica Host |
2–3 |
25–40 |
плод |
III |
|
Atriplex tatarica L. |
1–2 |
25–80 |
цветок |
II |
|
Poa bulbosa L. |
2 |
10–20 |
плод |
III |
|
Achillea millefolium L. |
4 |
35–50 |
цветок |
II |
|
Hordeum leporinum Link |
2 |
10–25 |
цветок |
III |
|
Camphorosma lessingii Litv. |
2–3 |
10–35 |
плод |
III |
|
Adonis flammea Jacq. |
2–3 |
10–45 |
плод |
II |
|
Eremopyrum triticeum (Gaertn.) Nevski |
2–3 |
10–30 |
плод |
III |
|
Velezia rigida L. |
2 |
10–50 |
плод |
III |
|
Allochrusa versicolor (Fisch.et C. A. Mey.) Boiss. |
3–4 |
18–30 |
цветок- плод |
III |
Andrachne filiformis широко распространена в горной зоне Автономной Республики, в фитоценозах на сухих каменистых склонах нижнего горного пояса. Отмечены ассоциации и микрогруппировки в развитых фитоценозах Валидагского участка Садаракского района и близлежащей горы Дахна. Андрахнея шаровидная обнаружена и в фитоценозах этих территорий, где широко распространены ксерофитные травы, некоторые деревья и кустарники. Фитоценоз немногочисленный, представлен 34–40 видами растений. Известные виды включают: Peganum harmala L., Rubia rigidifolia Pojark., Dorema glabrum Fisch. et C. A. Mey., Hypericum scabrum L., Scutellaria orientalis L., Teucrium polium L., Geranium tuberosum L., Helichrysum pallasii (Spreng.) Ledeb., Salsola crassa , S. nodulosa (Moq.) Iljin, Crambe orientalis L., Rheum ribes L. Общее проективное покрытие фитоценозов составляет 60–65% (Таблица 5).
Таблица 5
ВИДОВОЙ СОСТАВ И СТРУКТУРА ФИТОЦЕНОЗА, В КОТОРОМ ВИД Andrachne filiformis (SP III) ЯВЛЯЕТСЯ КОМПОНЕНТОМ ВОКРУГ ВАЛИДАГА ШАРУРСКОГО РАЙОНА
|
Название растений |
Обилие |
Высота (см) |
Фенофаза |
Ярус |
|
Andrachne filiformis Pojark. |
1–2 |
7–20 |
цветок, плод |
IV |
|
Daphne mucronata Royle |
2 |
35–50 |
плод |
III |
|
Astragalus tribuloides Delile |
3 |
10–15 |
плод |
IV |
|
Adonis flammea Jacq. |
2–3 |
10–45 |
плод |
II |
|
Eremopyrum triticeum (Gaertn.) Nevski |
2–3 |
10–30 |
плод |
III |
|
Spinacia tetrandra M. Bieb. |
1–2 |
10–40 |
плод |
III |
|
Aegilops cylindrica Host |
2–3 |
25–40 |
плод |
III |
|
Atriplex tatarica L. |
1–2 |
25–80 |
цветок |
II |
|
Euphorbia marschalliana Boiss. |
3–4 |
15–40 |
цветок, плод |
II |
|
Euphorbia szowitsii Fisch. et C. A. Mey. |
1–2 |
2–10 |
цветок, плод |
IV |
|
Koelpinia linearis Pall. |
2 |
15–30 |
плод |
III |
|
Ceratocarpus arenarius L. |
3 |
19–25 |
плод |
III |
|
Hypericum linarioides Bosse |
3 |
80–95 |
цветок |
II |
|
Achillea millefolium L. |
3–4 |
35–50 |
цветок |
I |
|
Hordeum leporinum Link |
2 |
10–25 |
цветок |
III |
|
Rheum ribes L. |
1–2 |
30–40 |
цветок, плод |
III |
|
Camphorosma lessingii Litv. |
2–3 |
10–35 |
плод |
III |
|
Poa bulbosa L. |
2 |
10–20 |
плод |
III |
|
Hedypnois cretica (L.) Dum. Cours. |
3 |
15–20 |
плод |
III |
Фитоценозы, сформированные с участием вида Andrachne telephioides , обнаружены на сухих склонах с каменистыми и редкокустарничковыми участками вокруг Нехрам-Дарешам Бабекского района. Большинство видов растений в ассоциациях и микрогруппировках здесь относятся к ксерофитам. Однако мезоксерофитные и ксеромезофитные растения встречаются и в затененных северных частях и понижениях.
Из кустарников и кустарничков Rhamnus pallasii, Amygdalus fenzliana, Daphne mezereum L., Rubia rigidifolia, Atraphaxis spinosa, Ephedra equisetina Bunge; травные растения Achillea tenuifolia Lam., Hypericum scabrum, Dorema glabrum и др. часто можно увидеть. Видовой состав небогат, но немногочислен, отмечено 15–45 видов растений.
Прогнозируемое покрытие фитоценоза видом Andrachne telephioides составляет 45–55% (Таблица 6).
Таблица 6
ВИДОВОЙ СОСТАВ И СТРУКТУРА ФИТОЦЕНОЗА, В КОТОРОМ КОМПОНЕНТОМ ЯВЛЯЕТСЯ Andrachne telephioides (SP IV) ВОКРУГ ДУЗДАГА БАБЕКСКОГО РАЙОНА
|
Название растений |
Обилие |
Высота(см) |
Фенофаза |
Ярус |
|
Andrachne telephioides L. |
2–3 |
17–25 |
цветок-плод |
III |
|
Euphorbia szowitsii Fisch. et C. A. Mey. |
1–2 |
2–10 |
цветок-плод |
IV |
|
Daphne mucronata Royle |
1–2 |
35–50 |
плод |
III |
|
Lepidium vesicarium L. |
3–4 |
80–100 |
цветок |
I |
|
Salvia hydrangea DC. ex Benth. |
3–4 |
10–40 |
цветок-плод |
III |
|
Ferula oopoda (Boiss. et Buhse) Boiss. |
1–2 |
90–100 |
цветок-плод |
I |
|
Atriplex tatarica L. |
1–2 |
25–80 |
цветок |
II |
|
Amberboa moschata (L.) DC. |
3–4 |
24–38 |
цветок |
III |
|
Ceratocarpus arenarius L. |
3 |
19–25 |
плод |
III |
|
Zygophyllum atriplicoides Fisch. et C. A. Mey. |
1–2 |
78–95 |
цветок-плод |
II |
|
Achillea millefolium L. |
3–4 |
35–50 |
цветок-плод |
I |
|
Erodium cicutarium (L.) L’Her. |
2–3 |
12–17 |
цветок |
IV |
|
Camphorosma lessingii Litv. |
2–3 |
10–35 |
плод |
III |
|
Bifora radians Bieb. |
2 |
15–20 |
цветок-плод |
IV |
|
Moltkia coerulea (Willd.) Lehm. |
2–3 |
23–34 |
цветок |
III |
В ценопопуляциях видов Andrachne изучали состав фитоценозов, отслеживали периоды онтогенеза особей в течение года, оценивали возрастные и продуктивные показатели (Таблица 7).
Сбор растений и онтогенетические наблюдения проводились два раза в месяц в течение вегетационного периода. На начальном этапе уточнялись онтогенетические особенности видов и разделялись особи на категории по возрасту. На каждой пробной площади были заложены прямоугольные разрезы для изучения возрастной структуры. На трансекте через каждые 10–20 м в зависимости от рельефа отводили пробные площади по 4 м2. На каждом участке выборки были определены критерии для всех видов по возрастному статусу. Плотность ценопопуляции оценивали по количеству особей на пробной площади.
ОЦЕНКА ЦЕНОПОПУЛЯЦИЙ ВИДОВ РОДА Andrachne
Таблица 7
|
Периоды онтогенеза |
ЦП |
||||
|
I ЦП |
II ЦП |
III ЦП |
IV ЦП |
||
|
Фазы роста |
j |
27,8 |
33,3 |
33,3 |
25 |
|
онтогенеза, всего в % |
im |
22,2 |
26,7 |
33,3 |
37,5 |
|
v |
11,1 |
6,7 |
11,2 |
12,5 |
|
|
g 1 - g 3 |
16,7 |
20 |
22,2 |
25 |
|
|
ss-s |
22,2 |
13,3 |
— |
— |
|
|
Индексы |
I b |
0,37 |
0,33 |
0,35 |
0,30 |
|
I q |
0,51 |
0,44 |
0,47 |
0,57 |
|
|
Δ |
0,34 |
0,29 |
0,15 |
0,21 |
|
|
ω |
0,23 |
0,28 |
0,31 |
0,33 |
|
Установлено, что все ценопопуляции относятся к ювенильному типу (так как Δ < 0,35 и ω < 0,60). Более высокий индекс эффективности вида A. filiformis с Δ =0,15 по сравнению с остальными можно объяснить отсутствием сенильных и субсенильных особей и преобладанием прегенеративных особей. Отмечено, что развитие вполне зрелых и генеративных особей ослаблено из-за влияния негативных факторов во всех ЦП, и особи не могут завершить онтогенетические циклы.
Таким образом, показатели индексов старения и восстановления были неудовлетворительными, а тот факт, что процесс старения превышал восстановительный процесс, свидетельствовал о низком темпе развития ЦП. Это приводит к тому, что числовая тенденция в ЦП со временем уменьшается и находится под угрозой исчезновения.
Список литературы Фитоценологическое изучение и оценка ценопопуляций некоторых видов Euphorbiaceae, распространенных в Нахичевани
- Bezerra F. W., do N Bezerra P., de Oliveira M. S., da Costa W. A., Ferreira G. C., de Carvalho R. N. Bioactive Compounds and Biological Activity of Croton Species (Euphorbiaceae): An Overview // Current Bioactive Compounds. 2020. V. 16. №4. P. 383-393. https://doi.org/10.2174/1573407215666181122103511
- Gatsuk L. E., Smirnova O. V., Vorontzova L. I., Zaugolnova L. B., Zhukova L. A. Age states of plants of various growth forms: a review // The Journal of Ecology. 1980. P. 675-696. https://doi.org/10.2307/2259429
- Maryam J., Bushra M., Abida Y., Mir A. K. Pharmacological activities of selected plant species and their phytochemical analysis // Journal of Medicinal Plants Research. 2012. V. 6. №37. P. 5013-5022. https://doi.org/10.5897/JMPR09.259
- Ibadullayeva S. J., Mamedova S. E., Sultanova Z. R., Movsumova N. V., Jafarli I. A. Medicinal plants of Azerbaijan flora used in the treatment of certain diseases // Afr. J. Pharm. Pharmacol. 2010. V. 4. №8. P. 545-548.
- Талыбов Т. Г., Ибрагимов А. Ш. Таксономический спектр флоры Нахичеванской автономной Республики. Нахчыван: Аджами. 2008. Т. 364. С. 202-210.
- Талыбов Т. Г., Сафарова Ф. А. Ядовитые растения Нахичеванской Автономной Республики. Нахчыван: Аджеми, 2017. 232 с.
- Марданлы С. Г. Лекарственные растения Нахичеванской Автономной Республики. Орехово-Зуево, 2018. 452 с.
- Гельтман Д. В. Род Euphorbia L.(Euphorbiaceae) во флоре Азербайджана // Новости систематики высших растений. 2016. Т. 47. С. 85-94.
- Глотов Н. В., Жукова Л. А. Демографо-генетические подходы к изучению популяций растений // Экология популяций: структура и динамика. 1995. Т. 1. С. 203-215.
- Злобин Ю. А., Скляр В. Г., Клименко А. А. Популяции редких видов растений: теоретические основы и методика изучения. Сумы: Университетская книга, 2013. Т. 439.
- Животовский Л. А. Популяционная биометрия. М.: Наука, 1991. 270 с.
- Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. 2001. №1. С. 3-7.
- Зубкова Е. В., Ханина Л. Г., Грохлина Т. И., Дорогова Ю. А. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы Ecoscalewin. Йошкар-Ола, 96 с.
- Мехтиева Н. П., Гельтман Д. В. Acalypha australis (Euphorbiaceae)-новый чужеродный вид для флоры Азербайджана // Ботанический журнал. 2015. Т. 100. №4. С. 403-406.
- Уранов А. А. Возрастной спектр ценопопуляций как функция времени и энергетических волновых процессов // Научные доклады высшей школы. Биологические науки. 1975. №2. С. 7-34.
- Работнов Т. А. Фитоценология. М.: Изд-во МГУ, 1992. 349 с.
- Раменский Л. Г. Проблемы и методы изучения растительного покрова. Л.: Наука, 1971. 334 с.
- Серебряков И. Г. Экологическая морфология растений: Жизненные формы покрытосеменных и хвойных. М.: Высш. школа, 1962. 378 с.
