Физиологические аспекты переваривания полносмешанного рациона в сложном желудке жвачных на примере крупного рогатого скота (Bos taurus taurus)
")
Автор: Василевский Н.В., Елецкая Т.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биологические основы кормления
Статья в выпуске: 4 т.54, 2019 года.
Бесплатный доступ
Современные системы нормированного кормления крупного рогатого скота базируются на положении, что различные технологии скармливания не оказывают влияния на процессы переваривания и всасывания питательных веществ одного и того же рациона. В наших исследованиях впервые получены данные, свидетельствующие об изменениях параметров процесса переваривания рациона в сложном желудке крупного рогатого скота в зависимости от способа скармливания - раздельной выдачи каждого из кормов и их одновременной выдачи в виде полносмешанного рациона. Цель работы состояла в изучении динамики суточного потока химуса, его химического состава и переваримости основных групп питательных веществ у бычков при переходе на полносмешанный рацион. Опыты проводили на физиологическом дворе Института животноводства Украинской академии аграрных наук в 2015-2016 годах. Объектом исследований были кастрированные бычки ( Bos taurus taurus ) украинской красно-пестрой породы (живая масса 300 кг) с дуоденальными канюлями, установленными в начале 12-перстной кишки (ДПК)...
Скот украинской красно-пестрой породы, переваривание, объем химуса, клетчатка, полносмешанный рацион
Короткий адрес: https://sciup.org/142222195
IDR: 142222195 | УДК: 636.2:591.132:636.084.41 | DOI: 10.15389/agrobiology.2019.4.787rus
Physiological aspects of complete mixed diet digestion in complex stomach of ruminants on the example of cattle (Bos taurus taurus)
Modern systems of normalized cattle feeding are based on the position that different feeding technologies do not affect the processes of digestion and absorption of nutrients of the same diet. In our studies, data were obtained for the first time, indicating changes in the parameters of the ration digestion process in the complex stomach of cattle, depending on the method of feeding: separate delivery of separate feeds and simultaneous delivery by the form of the Total Mixed Ration (TMR). To determine mechanisms of increasing feed dry matter intake in cattle when they were switched to Total Mixed Ration feeding, daily flux rates of chyme, its chemical composition and digestibility of main nutrient groups in the compound stomach of young cattle were studied. A Latin square design experiment was carried out using two diets differing in the forage fodder to concentrated fodder ratio and two methods of feeding (separate distribution of feed and in the form of TMR) by the method of periods with two Ukrainian red-motley bulls ( Bos taurus taurus ) of 300 kg live weight, with a sluice T-shaped duodenal canula...
Текст научной статьи Физиологические аспекты переваривания полносмешанного рациона в сложном желудке жвачных на примере крупного рогатого скота (Bos taurus taurus)
В исследованиях многих авторов (6-8) было показано, что переход на кормление крупного рогатого скота полносмешанным рационом (ПСР) увеличивает общее потребление сухого вещества в сравнении с раздельной раздачей компонентов того же рациона. К другим положительным эффектам относится отсутствие возможности выборочного поедания кормов и снижение конкурентных взаимоотношений между животными (9-11). Ранее мы установили влияние технологии скармливания рациона на переваримость питательных веществ (12).
В представленной работе нами впервые получены данные об изменениях процесса пищеварения у молодняка крупного рогатого скота в зависимости способа скармливания одного и того же рациона. Так, при полносмешанном типе кормления переваримость сырого жира и сырого протеина снижалась как на высоко-, так и на низкоконцентратном рационе (соответственно на 39,6 и 27,5 %), сырой клетчатки — повышалась (на 6,9 %), безазотистых экстрактивных веществ — существенно не изменялась для высоконцентратного рациона, но имела тенденцию к снижению для низкоконцентратного.
Цель работы состояла в изучении динамики суточного потока химуса, его химического состава и переваримости основных групп питательных веществ у бычков при переходе на полносмешанный тип кормления.
Обменную энергию рационов рассчитывали по формуле:
ОЭк = 14,46 - 0,0007 ½ СП + 0,0168 ½ СЖ - 0,0192 ½ СК - 0,00028 ½ БЭВ, где СП, СЖ, СК, БЭВ — соответственно содержание сырого протеина, сырого жира, сырой клетчатки и безазотистых экстрактивных веществ в рационе, г/кг сухого вещества (СВ) рациона, ОЭк — концентрация обменной энергии, Мдж/кг СВ (14).
В первый период скармливали рацион, выдавая в кормушку сено и силос, а поверх силоса — сухие концентрированные корма. Адаптация пищеварения бычков к изучаемому рациону происходила за 14 сут. Затем в течение 14 сут измеряли поступление химуса в ДПК за 9-часовый интервал после утреннего кормления (n = 6). Между измерениями делали перерывы на 1-2 сут для отдыха животных. Во второй период (28 сут) предварительно измельченное сено, силос и комбикорм взвешивали и смешивали (для каждого животного индивидуально), используя смеситель барабанного типа (изготовлен в лаборатории механизации животноводческих процессов Института животноводства УААН). Такой прием обеспечивал выдачу всех компонентов рациона в виде ПСР каждому животному с той же точностью, что и при раздельном кормлении.
Поступления химуса в ДПК измеряли по описанной выше методике. Химический анализ основных питательных веществ рационов, химуса и кала проводили по стандартным методикам (15). Перед опытом и после него животных взвешивали.
Статистические расчеты, построение графиков и диаграмм проводили с помощью пакета лицензионных программ Office Standard 2010 32-bit Russian (лицензия GGWA-A) методами дисперсионного анализа. Вычисляли средние арифметические значения (M), ошибки среднего (±SEM), среднее квадратическое отклонение (±σ). Достоверность различий оценивали с помощью парного t-критерия Стьюдента (для сопряженных рядов), что позволило устранить системную ошибку, возникающую вследствие индивидуальных особенностей пищеварения животных.
Результаты . Рационы для подопытных животных составляли таким образом, чтобы их характеристики различались как можно значительнее (табл. 1). Это было сделано для того, чтобы оценить изменения физиологических параметров пищеварения под влиянием скармливания ПСР на фоне диаметрально противоположных условий, возникающих в рубце при низко- и высококонцентратных рационах. Различия в обеспеченности энергией и протеином между рационами составляли более 20 %.
1. Состав и характеристика высоко- и низкоконцентратных рационов в опытах на бычках ( Bos taurus taurus ) украинской красно-пестрой породы (Институт животноводства Украинской академии аграрных наук, 2015-2016 годы)
|
Состав и характеристика |
Рацион |
Сопоставление рационов по |
|
|
высококонцен- |
низкоконцен- |
характеристикам (рацион ¹ 2 |
|
|
рациона |
тратный (¹ 1) |
тратный (¹ 2) |
к рациону ¹ 1, %) |
|
Силос кукурузный, кг |
11,00 |
16,80 |
|
|
Сено люцерновое, кг |
1,00 |
2,08 |
|
|
Дерть пшеничная, кг |
0,30 |
||
|
Дерть кукурузная, кг |
0,30 |
||
|
Жмых подсолнечниковый, кг |
3,30 |
||
|
Дерть ячменная, кг |
0,72 |
||
|
Минеральная добавка, кг |
0,03 |
0,03 |
|
|
Сухое вещество, г |
7008 |
6355 |
90,68 |
|
Органическое вещество, г |
6615 |
5996 |
90,65 |
|
Зола, г |
393 |
359 |
91,28 |
|
Сырой протеин, г |
1413 |
501 |
35,48 |
|
Сырой жир, г |
563 |
112 |
19,80 |
|
Сырая клетчатка, г |
1955 |
2170 |
111,01 |
|
Безазотистые экстрактивные |
|||
|
вещества, г |
2684 |
3213 |
119,74 |
|
Кальций, г |
42,55 |
30,74 |
72,24 |
|
Фосфор, г |
25,62 |
21,18 |
82,67 |
|
Обменная энергия, МДж |
71,52 |
50,86 |
71,10 |
|
Содержание обменной |
|||
|
энергии, МДж/кг |
10,21 |
8,00 |
78,41 |
|
Энергопротеиновое |
|||
|
соотношение, г/МДж |
19,76 |
9,86 |
49,89 |
Продолжение таблицы 1
Содержание сырого протеина, % 20,17 7,89 39,12
Обеспеченность основного обмена, МДж/кг 1,03 0,63 61,56
Прим еч ани е. Обеспеченность основного обмена приводится в расчете на 1 кг метаболической массы тела (живая масса в степени 0,75). Прочерк означает отсутствие компонента в рационе.
Среднесуточный прирост живой массы за время исследований составил 1,12 и 0,26 кг при размахе колебаний 0,95-1,29 и 0,21-0,32 кг соответственно на рационах ¹ 1 и ¹ 2, что согласуется с данными литературы (16, 17).
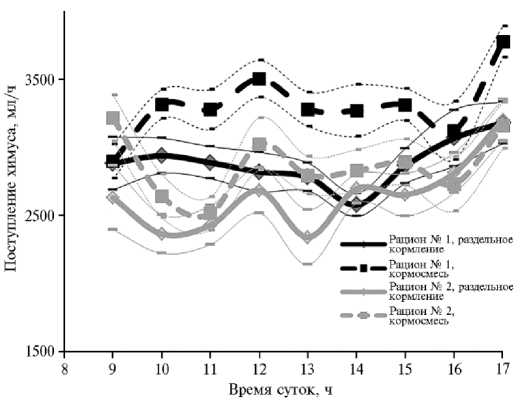
Динамика поступления химуса в 12-перстную кишку у бычков ( Bos taurus taurus ) украинской красно-пестрой породы при раздельном кормлении и содержании на полносмешанном рационе. Описание рационов и условия проведения опытов см. в разделе «Методика» (Институт животноводства Украинской академии аграрных наук, 2015-2016 годы).
(p < 0,001). С ПСР во всех случаях наблюдались
Скармливание ПСР усиливало поступление в ДПК кальция, фосфора и общего минерального остатка (табл. 2). При этом было установлено недостоверное, на уровне тенденции (0,1 > p > 0,05), увеличение количества Ca для рациона ¹ 1 и общей золы — для рациона ¹ 2, по всем остальным показателям повышение было достоверным (р < 0,05). Концентрация минеральных веществ в поступающем химусе, за исключением Ca, при переходе на ПСР ¹ 2 снижалась. Это происходило на фоне заметного повышения общей минерализации дуоденального химуса. Отсюда можно сделать вывод, что скармливание корма в виде ПСР в большей степени стимулирует поступление воды в сложный желудок.
2. Количество минеральных веществ в химусе в начальном участке 12-перстной кишки у бычков ( Bos taurus taurus ) украинской красно-пестрой породы при раздельном и полносмешанном типе кормления (Институт животноводства Украинской академии аграрных наук, 2015-2016 годы)
|
Рацион |
Раздельное кормление |
Полносмешанный рацион |
|||
|
Ca |
1 P 1 |
зола |
Ca 1 P 1 |
зола |
|
|
Поступило в |
12-перстн |
у ю к и ш к у, г/сут |
|||
|
¹ 1 |
60,71 |
48,99 |
602,5 |
64,27* 53,94** |
653,5** |
|
¹ 2 |
60,35 |
38,70 |
615,3 |
71,75*** 42,16**** |
669,5 |
|
Концентрация |
м и н е р ал о в в х и м у с е, мг% |
||||
|
¹ 1 |
87,68 |
70,51 |
868,0 |
81,75** 68,57 |
832,4**** |
|
¹ 2 |
96,88 |
61,94 |
982,1 |
104,51*** 61,03* |
971,0 |
П р и м е ч а н и е. Описание рационов и условия проведения опытов см. в разделе «Методика».
* Различия для полносмешанного рациона с показателями при раздельном кормлении статистически недостоверны (0,1 > p > 0,05).
**, ***, **** Различия для полносмешанного рациона с показателями при раздельном кормлении статистически значимы соответственно при р < 0,05; р < 0,01 и p < 0,001.
Дисперсионный анализ (табл. 3) выявил заметное влияние (более 50 % по шкале Чеддока, Cheddock scale) способа скармливания на поступление P (оба рациона) и Ca (рацион ¹ 2) в ДПК. При этом изученные факторы (состав рациона и форма скармливания) влияли на общее содержание минерального остатка в химусе ДПК.
3. Дисперсионный анализ влияния способа скармливания и состава рационов на поступление минеральных веществ в химус 12-перстной кишки у бычков ( Bos taurus taurus ) украинской красно-пестрой породы (Институт животноводства Украинской академии аграрных наук, 2015-2016 годы)
|
Степень влияния, % |
|||
|
Ca |
1 P 1 |
зола |
|
Состав рациона
|
Рацион ¹ 1 |
10,5 |
57,8 |
12,3 |
|
Рацион ¹ 2 |
59,2 Способ скармливания |
52,8 |
13,7 |
|
Раздельное кормление |
23,3 |
48,8 |
12,8 |
|
Полносмешанный рацион |
46,4 |
61,8 |
13,2 |
П р и м е ч а н и е. Описание рационов и условия опытов см. в разделе «Методика».
Исходя из достоверного увеличения поступления P в ДПК для обоих рационах, Ca — для рациона ¹ 2 и общей золы — для рациона ¹ 1 (см. табл. 2), можно предположить, что увеличение оттока химуса из рубца в наших опытах связано с усилением слюноотделения. В то же время полученные данные о некотором снижении концентрации изученных минера- лов свидетельствуют и о повышении поступления воды в рубец помимо ее поступления в составе слюны. Ввиду отсутствия данных о динамике потреблении воды животными сделать вывод о происхождении ее дополнительного количества не представляется возможным. Наиболее вероятно увеличение потребления и поступления воды непосредственно через стенку рубца. Сам факт разбавления рубцовой жидкости свидетельствует скорее об увеличении потребления воды, поскольку ее поступление из межклеточной жидкости, вероятнее всего, сопровождалось бы повышением количества электролитов в рубцовом содержимом без эффекта разбавления.
Некоторые авторы наблюдали эффект повышения скорости оттока, связанного с увеличением потребления воды при использовании гранулированных кормов (26). Если принять во внимание тот факт, что значительное количество жидкости всасывается при прохождении рубцового содержимого через книжку, а поступление электролитов из желудочного сока в сычуг в некоторой степени должно сглаживать эффект разбавления выпитой водой рубцового содержимого, то увеличение поступления воды в рубец при кормлении ПСР может быть гораздо больше обнаруженного нами. Известно также о функции книжки в регулировании водно-солевого баланса жидкости, поступающей из рубца. Следовательно, факт разбавления рубцовой жидкости в ДПК свидетельствует о значительном влиянии ПСР на водно-солевой баланс в рубце. Вероятной причиной наблюдаемого в наших опытах эффекта разбавления Сa и P в дуоденальном химусе могло быть то, что регуляция водного баланса организма осуществляется Na-зависимыми рецепторами (27, 28), а поддержание водно-солевого равновесия содержимого пищеварительного тракта — главным образом за счет ионов Na. Косвенным подтверждением этому может служить отсутствие сколь-нибудь заметного влияния как рациона, так и способа скармливания на поступление общей золы в ДПК (см. табл. 3).
* Различия для полносмешанного рациона с показателями при раздельном кормлении статистически недостоверны (0,1 > p > 0,05).
** Различия для полносмешанного рациона с показателями при раздельном кормлении статистически значимы при р < 0,05.
В результате дезаминирования аминокислот в процессе основного обмена в тканях организма образуется аммиак, токсичный для клетки. Для его обезвреживания выработан энергозатратный механизм синтеза мочевины из трех свободных аминогрупп (29). Следует отметить, что синтез мочевины происходит не только в печени животных, но и непосредственно в стенке рубца (30-32). Синтезированная таким образом мочевина поступает в кровь и составляет так называемый пул мочевины, из которого она частично направляется в рубец, а частично выводится с мочой. Основным механизмом, направляющим мочевину в рубец, служит ее экскреция со слюной. При попадании в рубец мочевина разрушается уреазами микрофлоры до аммиака, который используется в процессах микробиального синтеза для образования аминокислот и последующего синтеза микробного и протозойного белка. Наиболее эффективно проходит экскреция мочевины в околоушной железе (20-30 мг%), содержание этого соединения в крови составляет 10-15 мг%, в секрете подчелюстной и подъязычной желез — соответственно 10-15 и 8-10 мг% (33). В зависимости от интенсивности и равномерности образования мочевины в течение суток соотношение ее рециркуляции и выведения может существенно изменяться. Значительную роль в этом играет наличие в рационе животного доступной для использования микрофлорой энергии, особенно легкопереваримых углеводов (34).
Исходя из результатов нашего опыта, на синтез микробиального
СП влияет как кормовой фактор, так и интенсивность поступления в рубец мочевины в составе слюны. Так, в случае рациона ¹ 1 после перехода на ПСР переваримость СП в рубце достоверно повысилась, что свидетельствует о снижении вовлечения мочевины в процесс рециркуляции азота и ее более активном выведении из организма. Другими словами, количество доступного азота в рубце не обеспечивалось соответствующим количеством энергии для синтеза микробиального сырого протеина. При использовании рациона ¹ 2 потенциальная обеспеченность микрофлоры доступной энергией была выше, как следствие, эндогенный азот тоже вовлекался в микробиальный синтез. Усиление слюноотделения, вызванное ПСР, и связанное с этим повышенное поступление мочевины в рубец способствовало дополнительному синтезу микробиального СП, что отразилось на увеличении поступления СП в ДПК.
Следует учитывать, что с усиленным оттоком жидкости из рубца ускоряется удаление тонкоизмельченной нерастворимой фракции частиц кормов. Можно предположить, что усиление этого фракционного оттока уменьшало бы время микробиального ферментирования частиц корма, что неизбежно должно было приводить к снижению видимой переваримости питательных веществ. В наших опытах наблюдалось повышение переваримости БЭВ и снижение переваримости СЖ на рационе ¹ 1 при снижении переваримости БЭВ и повышении переваримости СЖ на рационе ¹ 2. По-видимому, рост интенсивности микробиальной ферментации в одном случае был достаточно высок и преобладал над снижением переваримости вследствие повышенной скорости оттока, а в другом, наоборот, снижение переваримости из-за увеличения фракционной скорости оттока не компенсировалось микробиальным синтезом, что и отразилось на повышении общей видимой переваримости.
Адекватная интерпретация полученных данных невозможна без учета того, что питательные вещества в рубце не только разрушаются, но и, вновь синтезируясь, накапливаются в составе микробиальной биомассы. Для оценки взаимодействия ускоренного фракционного оттока мелких частиц корма и улучшения условий ферментации вследствие ускоренного удаления продуктов микробиального брожения в качестве маркера следует рассматривать сырую клетчатку, поскольку она только переваривается в рубце и микрофлорой не синтезируется. В связи с этим установленное нами увеличение потока химуса при изменении способа скармливания на одном и том же типе рациона может снижать переваримость клетчатки вследствие ускоренного оттока в составе мелких частиц рубцового содержимого, тогда как при другом типе рациона, наоборот, увеличивать степень ее переваривания из-за улучшения условий ферментации (сдвиг рН в щелочную сторону).
Список литературы Физиологические аспекты переваривания полносмешанного рациона в сложном желудке жвачных на примере крупного рогатого скота (Bos taurus taurus)
- Golder H.M., Denman S.E., McSweeney C., Wales W.J., Auldist M.J., Wright M.M., Marett L.C., Greenwood J.S., Hannah M.C., Celi P., Bramley E., Lean I.J. Effects of partial mixed rations and supplement amounts on milk production and composition, ruminal fermentation, bacterial communities, and ruminal acidosis. J. Dairy Sci., 2014, 97(9): 5763-5785 ( ). DOI: 10.3168/jds.2014-8049
- Auldist M.J., Marett L.C., Greenwood J.S., Hannah M., Jacobs J.L., Wales W.J. Effects of different strategies for feeding supplements on milk production responses in cows grazing a restricted pasture allowance. J. Dairy Sci., 2013, 96(2): 1218-1231 ( ). DOI: 10.3168/jds.2012-6079
- Mertens D.R. Creating a system for meeting the fiber requirements of dairy cows. J. Dairy Sci., 1997, 80(7): 1463-1481 ( ). DOI: 10.3168/jds.S0022-0302(97)76075-2
- Teimouri Y.A., Valizadeh R., Naserian A., Christensen D.A., Yu P., Eftekhari Shahroodi F. Effects of alfalfa particle size and specific gravity on chewing activity, digestibility, and performance of Holstein dairy cows. J. Dairy Sci., 2004, 87(11): 3912-3924 ( ). DOI: 10.3168/jds.S0022-0302(04)73530-4
- Mirzaei-Aghsaghali A., Maheri-Sis N. Importance of "physically effective fibre" in ruminant nutrition: a review. Annals of Biological Research, 2011, 2(3): 262-270.
- Keunen J.E., Plaizier J.C., Kyriazakis I., Duffield T.F., Widowski T.M., Lindinger M.I., McBride B.W. Effects of a subacute ruminal acidosis model on the diet selection of dairy cows. J. Dairy Sci., 2002, 85(12): 3304-3313 ( ).
- DOI: 10.3168/jds.S0022-0302(02)74419-6
- Spiekers H., Potthast V. Erfolgreiche Milchviehfütterung. DLG-Verl., Frankfurt am Main, 2004.
- Khan M.A., Bach A., Castells L., Weary D.M., von Keyserlingk M.A.G. Effects of particle size and moisture levels in mixed rations on the feeding behavior of dairy heifers. Animal, 2014, 8(10): 1722-1727 ( ).
- DOI: 10.1017/S1751731114001487
- Kononoff P.J., Heinrichs A.J., Lehman H.A. The effect of corn silage particle size on eating behaviour, chewing activities and rumen fermentation in lactating dairy cows. J. Dairy Sci., 2003, 86(10): 3343-3353 ( ).
- DOI: 10.3168/jds.S0022-0302(03)73937-X
- Rottman L.W., Ying Y., Zhou K, Bartell P.A., Harvatine K.J. The effects of feeding rations that differ in neutral detergent fiber and starch concentration within a day on production, feeding behavior, total-tract digestibility, and plasma metabolites and hormones in dairy cows. J. Dairy Sci., 2015, 98(7): 4673-4684 ( ).
- DOI: 10.3168/jds.2014-8859
- Kmicikewycz A.D., Harvatine K.J., Heinrichs A.J. Effects of corn silage particle size, supplemental hay, and forage-to-concentrate ratio on rumen pH, feed preference, and milk fat profile of dairy cattle. J. Dairy Sci., 2015, 98(7): 4850-4868 ( ).
- DOI: 10.3168/jds.2014-9249
- Василевский Н.В., Елецкая Т.А. Цюпко В.В, Берестовая Л.Е. Влияние технологии скармливания и уровня сырого протеина в рационе на переваримость питательных веществ. Проблемы биологии продуктивных животных, 2013, 1: 67-74.
- Алиев А.А. Новейшие оперативные методы исследования жвачных животных. М., 1985.
- Валигура В.И. Закономерности переваривания и использования питательных веществ и энергии разноструктурных рационов овцами. Докт. дис. М., 1990.
- Лабораторнi методи дослiджень у бiологiї, тваринництвi та ветеринарнiй медицинi. Львiв, 2012.
- Цюпко В.В., Пронина В.В., Берус М.В. и др. Методические рекомендации по нормированию энергии в кормлении крупного рогатого скота. Харьков, 1989.
- AFRC Technical Committee on Responses to Nutrients, Report number 5, Nutritive requirements of ruminant animals: energy. Nutrition Abstracts and Reviews. Series B, Livestock Feeds and Feeding, 1990, 60(10): 729-804.
- Physiology of Digestion and Metabolism in the Ruminant. Proc. of the Third International Symposium, Cambridge, England, August 1969 /A.T. Phillipson (ed.). Oriel Press Limited., 1970.
- Чичилов А.В. Выделение макроэлементов околоушными слюнными железами лактирующих коров в связи с рубцовым метаболизмом. Автореф. канд. дис. М., 1984.
- Ishler V.A., Heinrichs A.J., Varga G.B. From feed to milk: understanding rumen function. Pennsylvania State University, 1996.
- Beal A.M. Salivary electrolyte concentrations and electrical potential difference across the parotid salivary duct of anaesthetized sodium-replete sheep. Australian Journal of Biological Sciences, 1980, 33(2): 197-204.
- Рузанов В.Е. Обмен калия, натрия и хлора у коров черно-пестрой, голштино-фризской пород и их помесей. Автореф. канд. дис. М., 2002.
- Алиев А.А. Достижения физиологии пищеварения сельскохозяйственных животных в ХХ веке (основные концепции). Сельскохозяйственная биология, 2007, 2: 12-23.
- Tirloni L., Reck J., Terra R.M., Martins J.R., Mulenga A., Sherman N.E., Fox J.W., Yates III J.R., Termignoni C., Pinto A.F.M., Vaz I.daS. Jr. Proteomic analysis of cattle tick Rhipicephalus (Boophilus) microplus saliva: a comparison between partially and fully engorged females. PLoS ONE, 2014, 9(4): e94831 ( ).
- DOI: 10.1371/journal.pone.0094831
- Боровский Е.В., Леонтьев В.С. Биология полости рта. М., 1991.
- Olsson K., McKinley M.J. Central control of water and salt intake in goats and sheep. In: Digestive physiology and metabolism in ruminants /Y. Ruckebusch, P. Thivend (eds.). Springer, Dordrecht, 1980: 161-175 ( ).
- DOI: 10.1007/978-94-011-8067-2_8
- Schröder B., Vössing S., Breves G. In vitro studies on active calcium absorption from ovine rumen. Journal of Comparative Physiology B, 1999, 169(7): 487-494 ( ).
- DOI: 10.1007/s003600050246
- Grabherr H., Spolders M., Lebzien P., Hüther L., Flachowsky G., Fürll M., Grün M. Effect of zeolite A on rumen fermentation and phosphorus metabolism in dairy cows. Archives of Animal Nutrition, 2009, 63(4): 321-336 ( ).
- DOI: 10.1080/17450390903020430
- Ёрсков Э.Р., Рил М. Энергетическое питание жвачных животных. Боровск, 2003.
- Mutsvangwa T., Davies K.L., McKinnon J.J., Christensen D.A. Effects of dietary crude protein and rumen-degradable protein concentrations on urea recycling, nitrogen balance, omasal nutrient flow, and milk production in dairy cows. J. Dairy Sci., 2016, 99(8): 6298-6310 ( ).
- DOI: 10.3168/jds.2016-10917
- Sun F., Aguerre M.J., Wattiaux M.A. Starch and dextrose at 2 levels of rumen-degradable protein in iso-nitrogenous diets: effects on lactation performance, ruminal measurements, methane emission, digestibility, and nitrogen balance of dairy cows. J. Dairy Sci., 2019, 102(2): 1281-1293 ( ).
- DOI: 10.3168/jds.2018-15041
- Fessenden S.W., Foskolos A., Hackmann T.J., Ross D.A., Block E., Van Amburgh M.E. Effects of a commercial fermentation byproduct or urea on milk production, rumen metabolism, and omasal flow of nutrients in lactating dairy cattle. J. Dairy Sci., 2019, 102(4): 3023-3035 ( ).
- DOI: 10.3168/jds.2018-15447
- Скопичев В.Г., Яковлев В.И. Частная физиология. Ч. 2. Физиология продуктивных животных. М., 2008.
- Василевский Н.В. Доступность сырого протеина для переваривания в тонком кишечнике и поступление эндогенного азота в сложный желудок бычков. Канд. дис. Харьков, 1993.
