Физиологические особенности адаптации сортов и отборных форм смородины красной к засухе и повышенным температурам

Автор: Панфилова О.В., Голяева О.Д.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетические и физиологические основы селекции
Статья в выпуске: 5 т.52, 2017 года.
Бесплатный доступ
Важное требование, которое предъявляется к возделываемым сортам, - это их адаптация к условиям выращивания. Смородина красная обладает высокой урожайностью, скороплодностью, хорошими пищевыми качествами ягод. В Центрально-Черноземном регионе к стрессорам, влияющим на ее урожай, относят засуху на фоне высоких температур. Целью настоящей работы было изучение основных показателей водного режима, фотосинтеза и продуктивности у представителей разных видов подрода Ribesia (Berl.) Jancz., а также выделение устойчивых к засухе и повышенным температурам сортов и отборных форм. Исследования проводили в 2011-2013 годах в Орловской области на 5 сортах и 3 отборных формах смородины красной (в том числе 6 образцов селекции Всероссийского НИИ селекции плодовых культур) разного эколого-географического и генетического происхождения, производных видов Ribes petraeum Wulf., R. vulgare Lam. и R. multiflorum Kit. Лист смородины красной имеет мезоморфную структуру, характерную для подрода Ribesia (Berl.) Jancz. Показано, что морфологические признаки (форма, жилкование) листьев определяются биологическими особенностями сорта, а условия произрастания влияют на анатомическую структуру. Выявлена положительная взаимосвязь между площадью листа и гидротермическим коэффициентом ( r = +0,99) и отрицательная - между площадью листа и температурой окружающей среды ( r = -0,97). Ответной реакцией на стрессовые факторы вегетационного периода становится увеличение толщины листа и разрастание клеток губчатой паренхимы (увеличение объема воздухоносных полостей), снижение содержания хлорофилла а, суммы хлорофиллов а + b и повышение количества каротиноидов. Одним из показателей адаптивности считается коэффициент отношения суммы хлорофиллов к каротиноидам; его высокие значения отмечали у сорта Hollandische Rote (5,14) и отборной формы 1426-21-80 (5,51). Выявлена взаимосвязь между суммой хлорофиллов и потерей воды ( r = -1,00), суммой хлорофиллов и количеством свободной воды ( r = +0,98). Содержание пигментов, фракционный состав воды, водоудерживающая способность листа были взаимосвязаны с развитием водного дефицита. Отмечена положительная корреляционная связь температуры воздуха и водного дефицита растений ( r = +0,84). Общее содержание воды в листьях смородины красной зависело от роста побегов, возраста листа, формирования ягод, сорта, метеоусловий и не было основным показателем устойчивости к повышенным температурам и засухе. Показатели соотношения связанной и свободной воды и водоудерживающая способность листьев варьировали в зависимости от метеорологических условий и водообеспеченности. В неблагоприятный по значениям гидротермического коэффициента 2012 год у всех генотипов смородины красной наблюдалось увеличение коэффициента соотношения связанной и свободной воды и снижение потери воды листом. Высокую адаптивность по показателям водного режима проявили образцы Hollandische Rote и 1426-21-80. Лабораторные методы диагностики согласовывались с полевыми данными устойчивости сортообразцов в условиях высокой температуры и дефицита влаги. Подтверждена перспективность использования физиологических экспресс-методов диагностики в селекции на адаптивность к деструктивным факторам в период вегетации. Комплексная оценка производные разных видов подрода Ribesia (Berl.) Jancz. показала их разную экологическую пластичность. Наибольшую устойчивость к засушливым условиям вегетационного периода Центрально-Черноземного региона проявили производные видов Ribes petraeum Wulf. (Hollandische Rote) и R. multiflorum Kit. (1426-21-80). Низкая засухоустойчивость отмечена у сортов и отборных форм R. vulgare Lam.
Смородина красная, засухоустойчивость, лист, мезоструктура, пигменты, фракционный состав воды, водоудерживающая способность
Короткий адрес: https://sciup.org/142214083
IDR: 142214083 | УДК: 634.722:581.1:632.112 | DOI: 10.15389/agrobiology.2017.5.1056rus
Physiological features of red currant varieties and selected seedling adaptation to drought and high temperature
Adaptation to weather conditions is an important feature of cultivars. To assess plant adaptability, we applied a physiological test of Ribesia (Berl.) Jancz. leaf resistance to abiotic factors during vegetation. The effect of high temperatures and drought on red currant plant photosynthetic apparatus and water regime was studied in 2011-2013 in the Central-Chernozem region (Orel region). Five varieties and three selected seedling genotypes of different eco-geographic and genetic origin were used including the derivatives of Ribes petraeum Wulf., R. vulgare Lam. and R. multiflorum Kit. (of these, six genotypes were originated in VNIISPK). Leaves of red currant plants are known to have a mesomorphic structure characteristic of Ribesia (Berl.) Jancz. subgenus. In this paper, it was shown that the morphological characteristics of leaves (i.e., shape, venation) are determined by the biological features of varieties while growing conditions alter the anatomical structure. We found a positive correlation between the leaf area and hydrothermal coefficient ( r = +0.99) and negative correlation between the leaf area and environment temperature ( r = -0.97). An increase in leaf thickness, the expansion of spongy parenchyma cells, a decrease in chlorophylls (both Chla and Chla + Chlb), as well as elevated carotenoid level are the response to stress factors during vegetation. The ratio coefficient of chlorophyll sum to carotenoids is considered as one of the adaptation indices. High coefficients were revealed in Hollandische Rote variety (5.14) and 1426-21-80 (5.51). Correlations between the chlorophyll sum and water loss ( r = -1.00) as well as chlorophyll sum and fraction of available water ( r = +0.98) were ascertained. The pigment content, fractional composition of water and water holding capacity of leaves are interconnected with the water shortage. The positive correlation of air temperature and water deficit ( r = +0.84) has been noted. The total water content in red currant leaves depends on the shoot growth, leaf age, variety, meteorological conditions and is not the main indicator of resistance to high temperatures and drought. Ratios of bound and free water and water holding capacity of leaves vary depending on meteorological conditions and water availability. In 2012 the weather conditions were unfavorable, and in all red currant genotypes the increase of the coefficient of bound water to available water as well as drop in water loss were observed. According to the parameters of water regime, Hollandische Rote variety and 1426-21-80 displayed high adaptability. The laboratory diagnostic methods were confirmed by the field data of genotype resistance under high temperature and moisture deficiency. Thus, the physiological express tests are suitable for estimation of red currant plant adaptability to environmental factors in the course of breeding. A comprehensive assessment showed diverse ecological plasticity in the plants with different Ribesia (Berl.) Jancz. subgenus species in the pedigree. The derivatives of Ribes petraeum Wulf. (Hollandische Rote) and R. multiflorum Kit. (1426-21-80) showed the highest tolerance to drought conditions during the vegetation season in the Central Chernozem region. The varieties and seedlings derived from R. vulgare Lam. had low drought resistance.
Текст научной статьи Физиологические особенности адаптации сортов и отборных форм смородины красной к засухе и повышенным температурам
Смородина красная — одна из ценных ягодных культур, обладающая высокой урожайностью, скороплодностью, хорошими пищевыми качествами ягод. От 50 до 80 % потерь ее урожая приходится на негативное действие природно-климатических факторов. Наиболее значимые из них — усиление нестабильности и стрессорности погодных условий (1, 2). В Центрально-Черноземном регионе к таким стрессорам относят засуху на фоне высоких температур. Повышение температуры вызывает морфо-анатомические, физиологические, биохимические изменения, которые влияют на 1056
рост и развитие растений и могут привести к снижению прибыли при производстве (3-5). Изучение морфологических особенностей, структуры фотосинтетического аппарата и водного обмена растений в связи с ареалом произрастания — главное условие для решения фундаментальных и прикладных задач в биологии культуры. В настоящее время приоритетным направлением становится использование физиолого-биохимических экспресс-методов диагностики устойчивости растений к неблагоприятным погодноклиматическим факторам. Это позволяет значительно оптимизировать селекционный процесс, минимизировать потери урожая и получить генотипы, устойчивые к деструктивному воздействию климатических аномалий (6, 7).
Растительные организмы обладают разными механизмами адаптации к стрессорам (8-10). Ксероморфная структура листа, изменения пигментного комплекса и водного баланса служат важными диагностическими признаками засухоустойчивости (11-13). Так, при усилении засухи у смородины черной Ribes americanum Mill. наблюдалось увеличение объема клеток столбчатого мезофилла, повышалось содержание трудноизвлекаемой воды в клетках. У некоторых сортов смородины красной, происходящих от вида R. vulgare Lam. (Голландская розовая, Натали красная), отмечалось снижение содержания суммы хлорофиллов а и b и интенсивности фотосинтеза (14-16). Подобную закономерность при повышении температуры наблюдали у земляники, винограда, каштана, растений чая ( Camellia sinensis ) и оливковых деревьев ( Olea evropaea L.) (17-20). У плодовых, овощных и зернобобовых культур отмечалось снижение водоудерживающей способности в течение светового дня, повышение транспирации и изменения в соотношении форм воды в разных частях растений (21-23). Особенности приспособления ягодных культур, в том числе смородины красной, к действию стрессоров в период вегетации малоизучены и требуют более детальных исследований.
В настоящей работе мы впервые использовали физиологические методы для диагностики устойчивости фотосинтетического аппарата листа к негативным факторам у растений смородины красной в период вегетации.
Цель работы — изучение основных показателей водного режима, фотосинтеза и продуктивности у представителей разных видов подрода Ribesia (Berl.) Jancz., а также выделение устойчивых к засухе и повышенным температурам сортов и отборных форм.
Методика. Наблюдения проводили в 2011-2013 годах на участке первичного сортоизучения смородины красной Всероссийского НИИ селекции плодовых культур (ВНИИСПК). Засушливость в периоды вегетации оценивали по величине гидротермического коэффициента (ГТК). Объектами исследования служили 8 генотипов смородины красной: 6 сорто-образцов селекции ВНИИСПК (Дана, Нива, Роза, 1518-37-14, 1426-21-80, 1432-29-98) и 2 — зарубежной (Jonkheer Van Tets, Hollandische Rote). Образцы имели разное генетическое и эколого-географическое происхождение и были производными видов Ribes petraeum Wulf. (Hollandische Rote, 1518-37-14), R. vulgare Lam. (Jonkheer Van Tets, Нива, Роза) и R. multiflorum Kit. (Дана, 1426-21-80, 1432-29-98).
Морфо-анатомическую структуру листьев оценивали, изучая их мезоструктуру на временных препаратах поперечных срезов (24) с использованием микроскопа Eclipse 50i («Nicon», Япония; увеличение ½400). Содержание пигментов определяли в 80 % ацетоновой вытяжке спектрофотометрическим методом (25). Для исследований использовался спектрофотометр Smart SpecTMPlus («Bio-Rad», США). Показатели водного режима определяли по разработанным методикам (11, 26, 27).
Для статистической обработки результатов проводили дисперсионный, корреляционный и регрессионный анализ при уровне достоверности 95 % (28) c использованием пакета программ Microsoft Excel 2010.
Результаты . Погодные условия сильно варьировали по годам исследований. С мая по июль 2011 года наблюдалась высокая влагообеспе-ченность на фоне повышенных температур (ГТК = 0,93; 1,12; 2,10; +30,0...+31,5 °С). Наиболее засушливым был 2012 год (ГТК = 0,20; 0,82; 0,40; +31,2...+32,2 °С). Вегетационный период 2013 года по гидротермическим условиям оказался промежуточным (ГТК = 0,72; 0,89; 0,68; +28,6^+31,5 °С).
Для представителей подрода Ribesia (Berl.) Jancz. характерны морфологическое разнообразие и экологическая пластичность. Смородина красная принадлежит к мезофитам. Все ее сорта и формы происходят от диких видов ( Ribes petraeum Wulf., R. rubrum L., R. vulgare Lam., R. multiflorum Kit.), произрастающих в умеренном поясе Северного полушария, в районах с относительно высокой увлажненностью почвы и умеренной температурой воздуха во время вегетации. Местом происхождения R. vulgare Lam. считается Европа, вид Ribes petraeum Wulf. распространен в горных районах Европы и Северной Африки, родина R. multiflorum Kit. — Южная Европа (29, 30). Изучение ассимиляционного аппарата у сортов и отборных форм разных видов смородины красной в условиях Центрального региона России представляет особый интерес, поскольку специфические климатические условия существенно влияют на формирование и функции листового аппарата.
Исследования показали, что некоторые морфологические признаки (форма, жилкование) листьев смородины красной зависели от биологических особенностей сорта, а на анатомическое строение влияли условия произрастания. В периоды с недостаточной водообеспеченностью площадь листовой поверхности снижалась у всех исследуемых образцов по сравнению с оптимальными условиями: в 2011 году она составляла в среднем 17,64 см2, в 2012 году — 13,95 см2; в 2013 году — 14,37 см2. При этом максимальные значения площади листа отмечали в 2011-2013 годах у формы 1426-21-80 (производного вида R. multiflorum Kit.). Была выявлена положительная взаимосвязь между площадью листа и ГТК ( r = +0,99) и отрицательная — между площадью и температурой окружающей среды ( г = -0,97).
Лист смородины красной имел характерную для подрода Ribesia (Berl.) Jancz. мезоморфную структуру. Губчатая паренхима преобладала над столбчатой (палисадной), эпидермальные клетки оказались достаточно крупными, причем на верхней стороне листа они были крупнее, чем на нижней. Высокая температура и засуха по-разному влияли на размеры клеток адаксиального эпидермиса и мезоструктуру листа. В 2012 и 2013 годах с мая по июль основные клетки адаксиального эпидермиса у сорта Holland-ische Rote (представителя R. petraeum Wulf.) и отборной формы 1426-21-80 ( R. multiflorum Kit.) при действии высоких температур (+31,2^+28,6 °С — в мае, +32,2^+31,5 °С — в июле) несколько вытягивались, что было связано со снижением тургора клеток. У остальных образцов мы отмечали сжатие клеток в тангентальном направлении (параллельно поверхности стебля) (рис. 1).
Повышенная температура и засуха в 2012 и 2013 годах приводили к разрастанию клеток паренхимы и увеличению общей толщины листа (табл. 1). Разрастание клеток мезофилла происходило главным образом за счет увеличения объема воздухоносных полостей губчатой паренхимы, что способствовало улучшению газообмена между листом и окружающей средой. Достоверно большее по сравнению с 2011 годом разрастание клеток губчатой паренхимы и толщины листовой пластинки были 1058
ыявлены практически у всех образцов (кроме сорта Нива), особенно у сорта Hollandische Rote и формы 1426-21-80. Эти изменения следует рассматривать как проявление высокой адаптивности анатомического строе- ния листа к стрессорам в период вегетации.
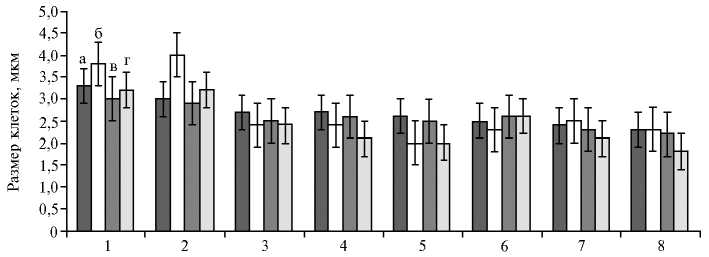
Образец
Рис. 1. Размеры клеток адаксиального эпидермиса листа у сортов и отборных форм смородины красной в мае (а) и июле (б) 2012 года, а также в мае (в) и июле (г) 2013 года: 1 — Hollandische Rote, 2 — 1426-21-80, 3 — Дана, 4 — 1432-29-98, 5 — Нива, 6 — 1518-37-14, 7 — Jonkheer Van Tets, 8 — Роза (Всероссийский НИИ селекции плодовых культур, Орловская обл., д. Жилина).
1. Мезоструктура листа у сортов и отборных форм смородины красной в разные годы (Всероссийский НИИ селекции плодовых культур, Орловская обл., д. Жилина)
|
Год (А) |
||||||||||
|
Образец (В) |
2011 |
2012 |
2013 |
|||||||
|
СП |
ГП 1 |
ОТЛ |
СП |
ГП |
ОТЛ |
СП |
ГП 1 |
ОТЛ |
||
|
1426-21-80 |
10,29 |
13,62 |
26,94 |
12,00 |
20,00 |
37,30 |
9,75 |
15,80 |
||
|
Hollandische Rote |
9,56 |
15,24 |
27,70 |
11,25 |
18,45 |
35,35 |
9,55 |
16,95 |
31,40 |
|
|
Нива |
9,28 |
12,65 |
24,63 |
10,15 |
15,35 |
29,60 |
10,35 |
15,30 |
29,10 |
|
|
Дана |
8,10 |
12,00 |
22,70 |
10,05 |
16,30 |
29,92 |
9,10 |
14,85 |
27,70 |
|
|
1432-29-98 |
7,90 |
12,35 |
23,25 |
11,65 |
17,55 |
33,65 |
10,15 |
15,10 |
29,10 |
|
|
1518-37-14 |
7,70 |
14,68 |
25,38 |
9,15 |
18,15 |
30,80 |
8,55 |
16,90 |
29,15 |
|
|
Jonkheer Van Tets |
7,30 |
10,10 |
20,33 |
10,50 |
15,90 |
30,05 |
9,00 |
12,30 |
24,80 |
|
|
Роза |
6,80 |
11,23 |
20,53 |
7,95 |
13,85 |
25,60 |
8,10 |
13,80 |
25,10 |
|
|
П р и м е ч а н и е. СП — |
столбчатая |
паренхима, мкм; ГП — губчатая паренхима, |
мкм; ОТЛ — общая тол- |
|||||||
|
щина листа, мкм. НСР05 для столбчатой паренхимы: А = 1,57; |
В = 2,56 |
; АВ = |
4,42. НСР05 для губчатой |
|||||||
|
паренхимы: А = 1,92; В |
= 3,14; АВ = |
5,43. НСР 05 для общей толщины листа: А = |
1,39; В |
= 2,27; АВ |
= 3,93. |
|||||
|
Пропуск означает отсутствие данных. |
||||||||||
Погодные условия вегетационных периодов оказали существенное влияние на работу ассимиляционного аппарата. В засушливый 2012 год наблюдалось снижение содержания хлорофилла а и хлорофиллов а + b (соответственно 1,23-2,21 и 2,15-3,36 мг/г) в листьях у всех сортообразцов смородины красной по сравнению с 2011 годом (1,53-2,46 и 2,40-3,92 мг/г) (рис. 2). Следует отметить, что более резкое снижение количества пигментов в 2012 году отмечалось у всех производных вида R. vulgare Lam., минимальное — у представителей видов R. petraeum Wulf. и R. multiflorum Kit. Незначительное ослабление засухи в 2013 году способствовало небольшому повышению содержания пигментов.
Засуха на фоне высоких температур стимулировала образование каротиноидов в клетке у всех исследуемых образцов подрода Ribesia (Berl.) Jancz., что согласуется с данными литературы о защитной функции этой группы пигментов при стрессе (21, 31, 32). Мы выявили положительную корреляцию между содержанием каротиноидов и температурой (r = +0,77). Некоторые исследователи для оценки устойчивости к стрессорам предлагают использовать коэффициент отношения суммы хлорофиллов к каротиноидам (31, 33, 34). В стрессовых условиях 2012 года по этому показате- лю выделялись сорт Hollandische Rote (величина коэффициента 5,14) и отборная форма 1426-21-80 (величина коэффициента 5,51), что может свидетельствовать об их устойчивости к засухе и высоким температурам летнего периода. Гидротермический режим влияет на продуктивность посредством воздействия на функциональное состояние растений, что подтверждалось высокими корреляциями между количеством хлорофилла а и урожайностью (r = +0,85), суммой хлорофиллов и урожайностью (r = +0,78).
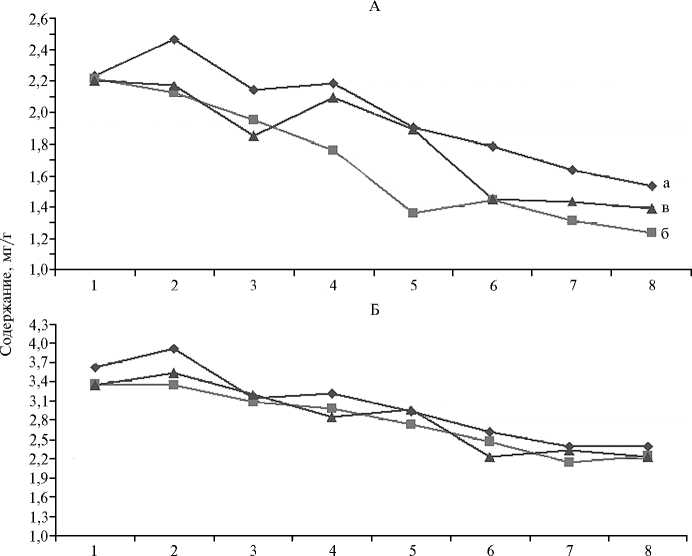
Образец
Рис. 2. Содержание хлорофилла а (А) и суммы хлорофиллов а + b (Б) в период вегетации 2011 (а), 2012 (б) и 2013 годов (в) в листьях у сортов и отборных форм смородины красной: 1 — Hollandische Rote, 2 — 1426-21-80, 3 — Дана, 4 — 1432-29-98, 5 — Нива, 6 — 1518-37-14, 7 — Jonkheer Van Tets, 8 — Роза (Всероссийский НИИ селекции плодовых культур, Орловская обл., д. Жилина). А: а — НСР 05 = 0,31; б — НСР 05 = 0,26; в — НСР 05 = 0,29. Б: а — НСР 05 = 0,61; б — НСР 05 = 0,53; в — НСР 05 = 0,58.
Зависимость содержания пигментов от показателей водного режима подтверждалась коэффициентом парной корреляции между суммой хлорофиллов и потерей воды ( r = -1,00), суммой хлорофиллов и количеством свободной воды ( r = +0,98). Эти показатели (содержание пигментов, фракционный состав воды, водоудерживающая способность листа) были связаны с развитием водного дефицита, который зависел от температуры воздуха ( r = +0,84). Взаимосвязь показателей водного режима и метеорологических особенностей вегетационного периода была описана с помощью коэффициентов множественной регрессии (табл. 2).
2. Корреляция между физиологическими показателями и метеорологическими условиями у исследуемых образцов смородины красной (Всероссийский НИИ селекции плодовых культур, Орловская обл., д. Жилина)
Показатель Коэффициент регрессии
Оводненность 0,90•F; r = 0,22
Свободная фракция воды 0,84•F; r = 0,33
Потеря воды - 0,88•F; r = 0,30
Прим еч ани е. F — значения гидротермического коэффициента, r — стандартная ошибка опыта.
Следует отметить, что общее содержание воды в листьях смородины красной не служит главным показателем устойчивости растений к повышенным температурам и засухе, поскольку зависит от целого ряда факторов (рост побегов, возраст листа, формирование ягод, метеоусловий и сорта) (35) (рис. 3).
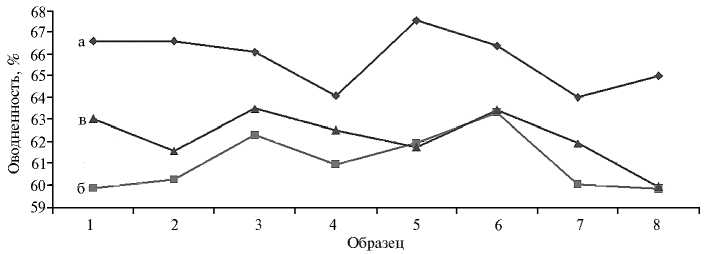
Рис. 3. Оводненность листьев в период вегетации 2011 (а), 2012 (б) и 2013 годов (в) у сортов и отборных форм смородины красной : 1 — Hollandische Rote, 2 — 1426-21-80, 3 — Дана, 4 — 143229-98, 5 — Нива, 6 — 1518-37-14, 7 — Jonkheer Van Tets, 8 — Роза (Всероссийский НИИ селекции плодовых культур, Орловская обл., д. Жилина). НСР 05 : а — 4,38, б — 3,74, в — 3,98.
Наиболее лабильны (в зависимости от метеорологических условий и водообеспеченности) показатели соотношения связанной и свободной воды и водоудерживающая способность листьев (11, 36, 37). Мы оценивали водоудерживающую способность по количеству потерянной воды. По изучаемым признакам в стрессовый период значительные различия были связаны с сортовыми и видовыми особенностями. В 2012 году, неблагоприятном по значениям ГТК, по сравнению с 2011 годом у всех образцов смородины красной наблюдалось увеличение коэффициента количественного соотношения связанной и свободной воды и снижение потери воды листом. В 2013 году это соотношение оказалось выше, чем в 2011 году, но ниже по сравнению с показателем 2012 года (табл. 3). Наиболее высокие значения коэффициента и минимальную потерю воды отмечали у сорта Hollandische Rote и отборной формы 1426-21-80, это дает основание считать, что генотипы этих образцов видов R. petraeum Wulf. и R. multiflorum Kit. проявляют более высокую адаптивность к воздействию высоких температур на фоне дефицита почвенной влаги.
3. Основные показатели водного режима у сортов и отборных форм смородины красной в разные годы (Всероссийский НИИ селекции плодовых культур, Орловская обл., д. Жилина)
|
Образец (В) |
Год (А) |
|||||||
|
2011 |
2012 |
2013 |
среднее |
|||||
|
СВЗ/СВ |
ПВ, % |
СВЗ/СВ |
ПВ, % |
СВЗ/СВ |
ПВ, % |
СВЗ/ СВ |
ПВ, % |
|
|
Hollandische Rote |
1,33 |
35,82 |
2,09 |
20,36 |
2,03 |
21,11 |
1,82 |
25,76 |
|
1426-21-80 |
1,31 |
36,16 |
2,41 |
20,62 |
2,09 |
19,37 |
1,94 |
25,38 |
|
1432-29-98 |
1,12 |
30,17 |
1,42 |
25,01 |
1,20 |
26,98 |
1,25 |
27,39 |
|
Jonkheer Van Tets |
1,01 |
38,10 |
1,31 |
21,85 |
1,04 |
27,40 |
1,12 |
29,12 |
|
1518-37-14 |
0,96 |
40,91 |
1,06 |
27,62 |
1,08 |
26,66 |
1,03 |
31,73 |
|
Нива |
0,82 |
42,56 |
1,71 |
24,84 |
1,50 |
25,58 |
1,34 |
30,99 |
|
Дана |
0,70 |
40,97 |
1,76 |
25,51 |
1,66 |
25,44 |
1,37 |
30,64 |
|
Роза |
0,63 |
39,75 |
0,98 |
26,82 |
0,87 |
27,85 |
0,83 |
31,47 |
|
НСР 05 |
0,08 |
2,06 |
0,11 |
3,03 |
0,14 |
1,86 |
||
|
П р и м е ч а н и е. |
СВЗ/СВ — |
коэффициент соотношения количества связанной и свободной воды, ПВ — |
||||||
|
потеря воды за 24 |
ч, % от оводненности. НСР05 для СВЗ/СВ: |
А — 2,18; В |
— 4,59; АВ — 7,95. |
НСР 05 для |
||||
|
ПВ: А — 0,13; В — |
0,21; АВ — |
0,37. |
||||||
Таким образом, показана перспективность использования физиологических методов при оценке устойчивости образцов подрода Ribesia (Berl.)
Jancz. к неблагоприятным погодным факторам. Полученные результаты полностью согласовывались с полевыми данными по устойчивости сортов и отборных форм смородины красной в условиях повышенной температуры и дефицита влаги. Это позволяет использовать рассмотренные методы как экспресс-диагностику и существенно интенсифицировать селекцию на адаптивность. Как показала комплексная физиологическая оценка, производные разных видов подрода Ribesia (Berl.) Jancz. обладают неодинаковой экологической пластичностью, что объясняется различием их генетического и эколого-географического происхождения. Наибольшую устойчивость к засушливым условиям вегетационного периода Центрально-Черноземного региона проявили производные видов Ribes petraeum Wulf. (Hollandi-sche Rote) и R. multiflorum Kit. (1426-21-80). Низкая засухоустойчивость отмечена у сортов и отборных форм R. vulgare Lam.
Список литературы Физиологические особенности адаптации сортов и отборных форм смородины красной к засухе и повышенным температурам
- Vollenweider P., Günthardt-Goerg M.S. Diagnosis of abiotic and biotic stress factors using the visible symptoms in foliage. Environ. Pollut., 2005, 137: 455-465 ( ) DOI: 10.1016/j.envpol.2005.01.032
- Christensen J.H., Christensen O.B. A summary of the PRUDENCE model projections of changes in European climate by the end of this century. Climatic Change, 2007, 81: 7-30 ( ) DOI: 10.1007/s10584-006-9210-7
- Kim K., Portis A.R. Temperature dependence of photosynthesis in Arabidopsis plants with modifications in Rubisco actives and membrane fluidity. Plant Cell Physiol., 2005, 46: 522-530 ( ) DOI: 10.1093/pcp/pci052
- Beck E.H., Fettig S., Knake C., Hartig K., Bhattarai T. Specific and unspecific responses of plants to cold and drought stress. J. Biosciences, 2007, 32: 501-510 ( ) DOI: 10.1007/s12038-007-0049-5
- Hasanuzzaman M., Nahar K., Fujita M. Extreme temperature responses, oxidative stress and antioxidant defense in plants. In: Abiotic stress -plant response and applications in agriculture/K. Vahadati, C. Leslie (eds.). INTECH, 2013 ( ) DOI: 10.5772/54833
- Neeru K., Kalpna B., Kadambot H.M.S., Harsh N. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food & Agriculture, 2016, 2(1): 271-313 ( ) DOI: 10.1080/23311932.2015.1134380
- Wise R.R., Olson A.J., Schrader S.M., Sharkey T.D. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ., 2004, 27(6): 717-724 ( ) DOI: 10.1111/j.1365-3040.2004.01171.x
- Wahid A., Gelani S., Ashraf M., Foolad M. Heat tolerance in plants: An overview. Environ. Exp. Bot., 2007, 61: 199-223 ( ) DOI: 10.1016/j.envexpbot.2007.05.011
- Bacelar E.A., Santos D.L., Moutinho-Pereira J.M., Gonçalves B.C., Ferreira H.F., Correia C.M. Immediate responses and adaptative strategies of three olive cultivars under contrasting water availability regimes: Changes on structure and chemical composition of foliage and oxidative damage. Plant Science, 2005, 170(3): 596-605 ( ) DOI: 10.1016/j.plantsci.2005.10.014
- Koini M.A., Alvey L., Allen T., Tilley C.A., Harberd N.P., Whitelam G.C., Franklin K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol., 2009, 19: 408-413 ( ) DOI: 10.1016/j.cub.2009.01.046
- Кушниренко М.Д. Методы изучения водного обмена и засухоустойчивости плодовых растений. Кишинев, 1970.
- Давлатов С.Х., Ашуров А.А., Байкова Е.В. Морфо-анатомическое строение листа дикорастущих видов рода Berberis из Таджикистана. Вестник Томского государственного университета, 2009, 323: 348-350.
- Схаляхо Т.В., Ненько Н.И, Киселева Г.К. Оценка степени засухоустойчивости сортов винограда в условиях Анапо-Таманской зоны по физиолого-биохимическим и анатомо-морфологическим показателям. Плодоводство и виноградарство Юга России, 2012, 17: 69-78.
- Резанова Т.А., Сорокопудов В.Н. Некоторые особенности анатомо-морфологического строения Ribes americanum Mill. Мат. Межд. науч.-практ. конф. «Биологически активные соединения природного происхождения: фитотерапия, фармацевтический маркетинг, фармацевтическая технология, ботаника». Белгород, 2008: 133-135.
- Резанова Т.А. Морфо-анатомические и экологические особенности Ribes americanum Mill. при интродукции на юге Среднерусской возвышенности. Автореф. канд. дис. Саратов, 2010.
- Филатова Л.А., Пачина Т.Д. Некоторые физиологические особенности двухгодичных саженцев разных сортов красной смородины. Вестник Пермского университета, 2007, 5(10): 28-30.
- Du Y.Y., Shin S., Wang K.R., Lu J.L., Liang Y.R. Effect of temperature on the expression of genes related to the accumulation of chlorophylls and carotenoids in albino tea. The Journal of Horticultural Science and Biotechnology, 2009, 84(3): 365-369 ( ) DOI: 10.1080/14620316.2009.11512533
- Bacelar E.A. Ecophysiological responses of olive (Olea europaea L.) to restricted water availability: Limitations, damages and drought resistance mechanisms. Universidade De Trás-Os-Montes E Alto Douro, Vila Real, 2006.
- Zhang B.C., Laranjo J.G., Correia C.M., Moutinho-Pereira J.M., Carvalho Goncalves B.M., Bacelar E.A., Peixoto F.P., Response V.G. Response, tolerance and adaptation to abiotic stress of olive, grapevine and chestnut in the Mediterranean region: role of abscisic acid, nitric oxide and microRNAs. In: Plants and environment/K.N. Hemanth (ed.). INTECH, 2011: 179-206 ( ) DOI: 10.5772/24719
- Хвостов Д.С. Механизмы адаптации и засухоустойчивость сортов и гибридов земляники. Автореф. канд. дис. Краснодар, 2000.
- Кисилева Н.С. Влияние стресс-факторов внешней среды на морфо-анатомическое строение листовой пластинки груши. Мат. Межд. науч.-практ. конф. «Адаптивный потенциал и качество продукции сортов и сорто-подвойных комбинаций плодовых культур». Орел, 2012: 115-122.
- Mazorra L.M., Nunez M., Echerarria E., Coll F., Sánchez-Blanco M.J. Influence of brassinosteriods and antioxidant enzymes activity in tomato under different temperatures. Biologia Plantarum, 2002, 45: 593-596 ( ) DOI: 10.1023/A:1022390917656
- Omae H., Kumar A., Shono M. Adaptation to high temperature and water deficit in the common bean (Phaseolus vulgaris L.) during the reproductive period. Journal of Botany, 2012, (2012): Article ID 803413 ( ) DOI: 10.1155/2012/803413
- Мокроносов А.Т., Борзенкова Р.А. Методика количественной оценки структуры и функциональной активности фотосинтезирующих тканей и органов. Труды по прикладной ботанике, генетике и селекции, 1978, 61(3): 119-128.
- Гавриленко В.Ф., Ладыгина М.Е., Хандобина Л.М. Большой практикум по физиологии растений. Фотосинтез. Дыхание. М., 1975.
- Баславская С.С., Трубецкова О.М. Практикум по физиологии растений. М., 1964.
- Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур/Под ред. Е.Н. Седова, Т.П. Огольцовой. Орел, 1999.
- Доспехов Б.А. Методика полевого опыта с основами статистической обработки результатов исследования. М., 1985.
- Федоровский В.Д. Ribes spicatum Robson. -смородина колосистая (систематика, география, изменчивость, интродукция). Киев, 2001.
- Сорокопудов В.Н., Соловьева А.Е., Смирнов А.С. Красная смородина в лесостепи Приобья. Новосибирск, 2005.
- Криворучко В.П. Эколого-биологические основы повышения продуктивности яблоневых садов северного Кыргызстана. Док. дис. Бишкек, 1998.
- Хвостов Д.С., Киртбая Е.К. Адаптационный потенциал земляники в условиях юга России. Садоводство и виноградарство, 1999, 4: 16-17.
- Киселева Н.С. Оценка адаптационной способности различных генотипов груши по морфоанатомическому и физиологическому состоянию листьев. Сельскохозяйственная биология, 2009, 3: 34-38.
- Ненько Н.И., Ильина И.А., Схаляхо Т.В. Адаптационная устойчивость винограда в летний период. Мат. Межд. науч. конф. «Высокоточные технологии производства, хранения и переработки винограда». Краснодар, 2010: 50-59.
- Панфилова О.В., Ожерельева З.Е., Голяева О.Д. Сравнительная оценка адаптивного потенциала генотипов смородины красной (Ribes rubrum L.) к абиотическим факторам вегетационного периода. Сортовивчення та охорона прав на сорти рослин, 2014, 1(22): 24-27.
- Machado S., Paulsen G.M. Combined effects of drought and high temperature on water relations of wheat and sorghum. Plant Soil, 2001, 233(2): 179-187 ( ) DOI: 10.1023/A:1010346601643
- Миронова Л.Н., Денисова С.Г., Зейнетдинова Г.С., Реут А.А. К оценке адаптационного потенциала декоративных многолетников. Вестник Воронежского государственного университета, Серия: География. Геоэкология, 2011, 1: 157-159.
