Физиологические признаки перспективных штаммов бактерий родов Bacillus и Pseudomonas - продуцентов микробиопрепаратов

Автор: Асатурова А.М.
Статья в выпуске: 2 (141), 2009 года.
Бесплатный доступ
С целью разработки лабораторного регламента массовой наработки микробиопрепаратов были изучены физиологические признаки перспективных штаммов бактерий-антагонистов родов Bacillus и Pseudomonas в условиях периодического культивирования. На основе полученных данных установлены оптимальные температура культивирования и кислотность среды, а также источники углеродного и азотного питания штаммов-продуцентов микробиопрепаратов.
Бактерии-антагонисты, микробиопрепарат, периодическое культивирование, физиологические признаки, температура культивирования, кислотность среды (рн), источники углерода и азота
Короткий адрес: https://sciup.org/142150910
IDR: 142150910
Physiological features of perspective strains of bacterium Bacillus and Pseudomonas genera - producers of microbiopreparations
With the purpose to develop laboratory regulations of mass groundworks of microbiopreparations, the physiological traits of perspective strain of bacteria-antagonists of Bacillus and Pseudomonas genera were studied in conditions of periodical cultivation. The obtained data allowed to determine optimal temperature of cultivation and medium acidity, as well as sources of carbonic and nitrogen nutrition of microbiopreparation strains-producers.
Текст научной статьи Физиологические признаки перспективных штаммов бактерий родов Bacillus и Pseudomonas - продуцентов микробиопрепаратов
Введение. Известно, что основной стадией любого микробиологического производства является производственное культивирование соответствующего микроорганизма, проводимое либо с целью увеличения микробной массы – биомассы, либо для получения продуктов метаболизма растущей популяции микроорганизмов [1]. Для разработки производства биофунгицидов комплексного действия необходимо получение жидкой культуры (ЖК) с оптимальной плотностью микробных клеток и концентрации антифунгальных веществ. Для этого были изучены физиологические признаки перспективных штаммов бактерий родов Bacillus и Pseudomonas : условия культивирования и кислотность среды, а также источники углеродного и азотного питания.
Материалы и методы. Объектом исследований служили штаммы бактерий-антагонистов Fa 4-1 Bacillus subtilis , D 7-1 B. subtilis , Sgrc-1 Pseudomonas fluorescens и Oif 2-1 Pseudomonas sp.
Для определения оптимальной температуры культивирования штаммы выращивали на жидких питательных средах Кинга В [2] (для штаммов Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. ) и Тайлона 3 [3] (для штаммов Fa 4-1 B. subtilis и D 7-1 B. subtilis ) при температурах 20, 25, 30 и 35 °С.
Оптимальную кислотность среды определяли, выращивая штаммы на вышеуказанных жидких питательных средах при оптимальных температурах. Добавлением лимонной кислоты или щелочи (NaOH) pH среды устанавливали в пределах 3, 6, 8, и 10.
Для определения оптимальных источников углеродного и азотного питания штаммы Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. выращивали на жидкой питательной среде Кинга В, а штаммы Fa 4-1 B. subtilis и D 7-1 B. subtilis – Чапека для бактерий [4]. Источниками углерода служили глюкоза, сахароза, меласса и глицерин. При изучении углеродных источников неизменными компонентами азотного питания для бацилл был азотнокислый натрий, а для псевдомонад – пептон. Азотсодержащие вещества (азотнокислый натрий, пептон, дрожжевой и кукурузный экстракты) добавляли в среды в качестве единственного источника азота с неизменными источниками углеродного питания: глюкоза (для бактерий рода Bacillus ) и глицерин (для бактерий рода Pseudomonas ).
Бактерии культивировали в течение двух суток на термостатированных качалках (195 об/мин) в колбах Эрленмейера (750 мл) с объемом питательной среды 150 мл и предварительным внесением посевной (маточной) культуры (2 % от объема питательной среды).
По окончании культивирования определяли численность бактериальных клеток. Для определения численности бактериальных клеток в ЖК использовали метод Коха [5]. Определение числа клеток этим методом включает три этапа: приготовление разведений, посев на питательную среду в чашку Петри (ЧП) и подсчет выросших колоний.
Для приготовления разведений стерильную водопроводную воду разливали по 9 мл в пробирки. Затем 1 мл исследуемой суспензии пипеткой переносили в колбу с 99 мл стерильной воды. Затем отбирали 1 мл суспензии и переносили в пробирку с 9 мл воды. Полученное разведение тща- тельно перемешивали, несколько раз вбирая в пипетку и выпуская из нее полученную суспензию. Таким же образом готовили все последующие разведения. Высев исследуемой ЖК осуществляли глубинным способом. Для этого по 1 мл из соответствующего разведения переносили в стерильные ЧП. Затем заливали в чашки по 15-20 мл среды, расплавленной и остуженной до 45-50 °С, и смешивали питательную среду с посевным материалом легкими вращательными движениями, после чего чашки оставляли на горизонтальной поверхности до застывания среды. Колонии бактерий подсчитывали через пять-семь суток инкубации. Количество клеток в 1 мл исследуемой ЖК вычисляли по формуле:
Т = a х 10n ,
V где Т – количество колониеобразующих единиц (КОЕ) в 1 мл;
а – среднее число колоний, выросших после посева из данного разведения;
-
V – объем суспензии, взятый для посева;
-
10n – коэффициент разведения.
Математическую обработку опытных данных проводили с использованием стандартных компьютерных программ.
Определение оптимальной температуры и реакции среды для культивирования штаммов бактерий-антагонистов
Существенное значение для роста микроорганизмов имеет такой фактор внешней среды как температура. Результаты проведенных опытов показали, что отношение к температуре у изучаемых штаммов неодинаково.
Высокая плотность клеток штаммов бацилл-антагонистов обнаруживалась только при температурном режиме 30-35 °С. При этом анализ опытных образцов показал, что оптимальной температурой, при которой происходит максимальное накопление микробной массы и практически полная утилизация питательной среды, для штамма D 7-1 B. subtilis является 30 °С, а для штамма Fa 4-1 B. subtilis – диапазон 30-35 °С (рис. 1).
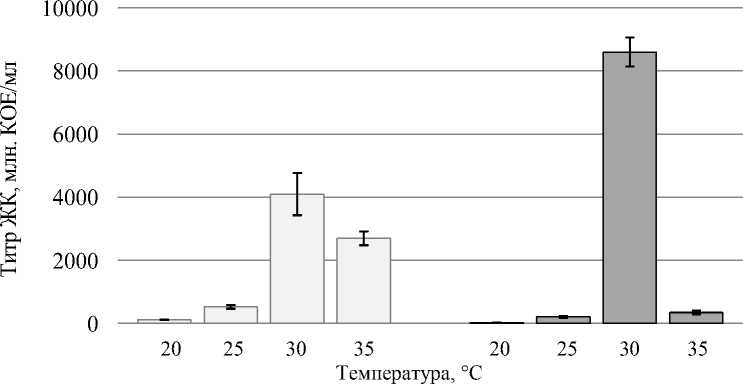
Fa 4-1 B. subtilis D 7-1 B. subtilis
Рисунок 1 – Рост штаммов бактерий-антагонистов рода Bacillus при различных температурах в процессе периодического культивирования
Существенным достоинством штаммов псевдомонад по сравнению с бациллярными бактериями была способность к активному росту при более низких температурах: 20-25 °С (рис. 2).
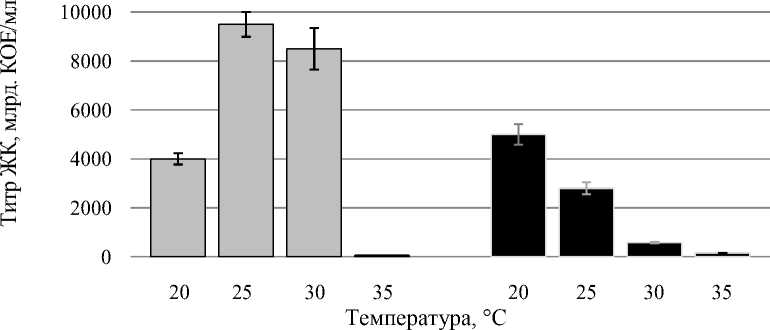
Sgrc-1 P. fluorescens Oif 2-1 Pseudomonas sp.
Рисунок 2 – Рост штаммов бактерий-антагонистов рода Pseudomonas при различных температурах в процессе периодического культивирования
Несмотря на то, что титр клеток оставался высоким вне зависимости от температурного режима и составлял 2,8-9,5 × 1012 КОЕ/мл при 20-25 °С и от 6,0 × 1010 до 8,5 × 1012 КОЕ/мл при 3035 °С, повышение температуры культивирования до 30-35 °С приводило к появлению клеток с отсутствием желто-зеленого внеклеточного пигмента, что связано с потерей клетками антифунгаль-ной активности (рис. 3) [6].
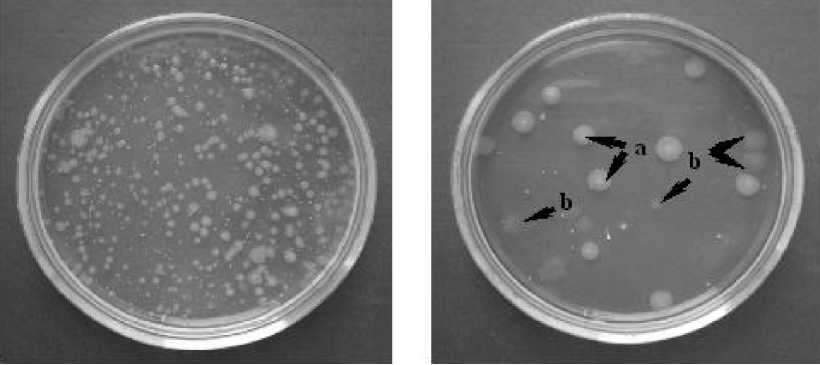
1 2
Рисунок 3 – Влияние температуры на пигментообразующую способность штаммов бактерий-антагонистов рода Pseudomonas (ориг.)
1 – типичные клетки с желто-зеленым внеклеточным пигментом (культивирование при температуре 20-25 °С);
2 – типичные (а) и нетипичные (b) клетки с отсутствием желто-зеленого внеклеточного пигмента (культивирование при температуре 35 °С).
Следовательно, оптимальной для роста штаммов флюоресцирующих бактерий Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. является температурный диапазон 20-25 °С.
Одним из важных факторов, определяющих нормальный рост бактерий, является реакция среды. При изменении ее в неблагоприятную сторону микроорганизм перестает расти даже в тех случаях, если все остальные условия окружающей среды будут оптимальными [7].
При определении оптимальной кислотности среды у всех исследованных штаммов отмечался достаточно высокий титр ЖК при рН среды 6 и 8. Лимитирующими рост бактерий-антагонистов оказались значения реакции среды 3 и 10 (рис. 4, 5).
У штаммов Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. при указанных уровнях рН практически не обнаруживалось роста клеток, а в случае с бациллярными штаммами были получены ЖК с низкой плотностью бактериальных клеток: 1,4-4,0 х 106 КОЕ/мл при рН 3 и 4,75,6 ×106 КОЕ/мл при рН 10. Однако, принимая во внимание полученный титр (для штаммов Fa 4-1 B. subtilis , D 7-1 B. subtilis 8,5×108 и 2,3×109 КОЕ/мл соответственно) и объем посевной культуры (3 мл на 150 мл питательной среды), можно сделать вывод о том, что при рН среды 3 и 10 бактерии рода Bacillus не размножались, а находились в состоянии покоя, сохраняя жизнеспособность благодаря наличию споровой стадии (рис. 4).
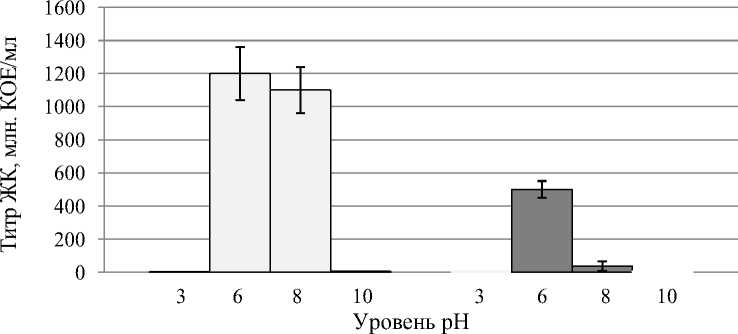
Fa 4-1 B. subtilis D 7-1 B. subtilis
Рисунок 4 – Рост штаммов бактерий-антагонистов рода Bacillus при различных уровнях рН в процессе периодического культивирования
В отличие от бацилл штаммы бактерий рода Pseudomonas не формируют спор и других форм покоя, поэтому при рН среды 3 полностью погибали, а при рН 10 выживали лишь единичные клетки (рис. 5).
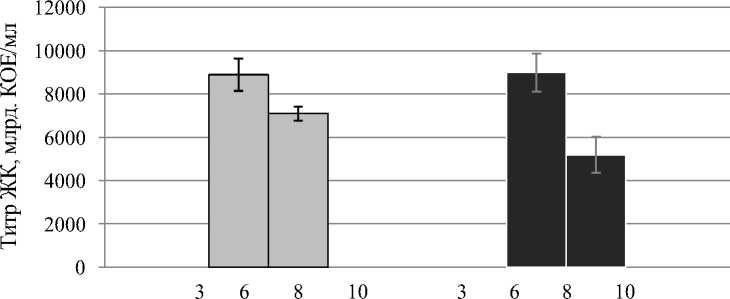
Уровень рН
Sgrc-1 P. fluorescens Oif 2-1 Pseudomonas sp.
Рисунок 5 – Рост штаммов бактерий-антагонистов рода Pseudomonas при различных уровнях рН в процессе периодического культивирования
Следовательно, для перспективных биоагентов Fa 4-1 B. subtilis , Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. оптимальным диапазоном реакции среды является 6-8, а для штамма D 7-1 B. subtilis – рН 6. Именно при данных значениях реакции среды отмечена высокая плотность популяций бактерий и максимальная утилизация питательной среды.
Определение оптимальных источников углеродного и азотного питания для культивирования штаммов бактерий-антагонистов
Эффективность использования штаммами-антагонистами источников углерода и азота определялась также по количеству колониеобразующих единиц.
Максимальная плотность клеток штамма Fa 4-1 B. subtilis отмечалась на среде, где в качестве источников углерода были меласса и глюкоза (1,3-1,6 × 109 КОЕ/мл), а для штамма D 7-1 B. subtilis – меласса и сахароза (1,9-2,8 × 109 КОЕ/мл) (рис. 6).

Fa 4-1 B. subtilis
D 7-1 B. subtilis
Рисунок 6 – Рост штаммов бактерий-антагонистов рода Bacillus в зависимости от источников углерода в процессе периодического культивирования
Минимальный титр штаммов Fa 4-1 B. subtilis и D 7-1 B. subtilis отмечен на питательной среде с добавлением в качестве источника углеродного питания глицерина (1,1 × 107 и 8,7 × 106 КОЕ/мл соответственно). Это указывает на неспособность бациллярных бактерий использовать данное соединение. Несмотря на присутствие всех остальных компонентов среды, а также оптимальные температуру и реакцию среды, данные количественного учета свидетельствуют о том, что размножения бактерий с данным источником углерода не происходило.
Штаммы пигментирующих бактерий оказались более пластичными и активно использовали весь спектр испытываемых источников углерода, что наглядно показал анализ образцов ЖК штаммов. Максимальный титр для штамма Sgrc-1 P. fluorescens отмечен в вариантах с добавлением сахарозы, глюкозы и глицерина (2,9-4,8 × 1012 КОЕ/мл), а для штамма Oif 2-1 Pseudomonas sp. – глицерина и глюкозы (2,6-4,9 × 1012 КОЕ/мл) (рис. 7).
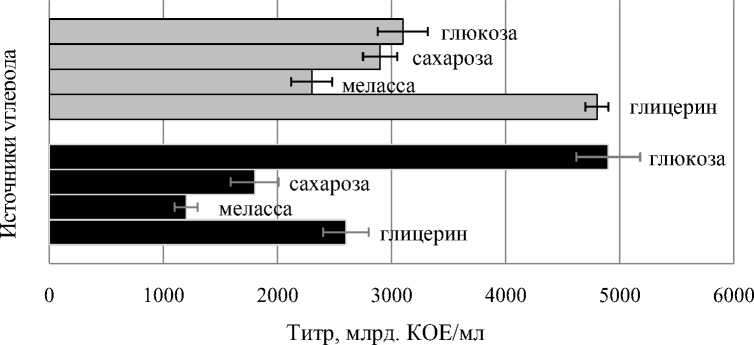
Sgrc-1 P. fluorescens
Oif 2-1 Pseudomonas sp.
Рисунок 7 – Рост штаммов бактерий-антагонистов рода Pseudomonas в зависимости от источников углерода в процессе периодического культивирования
Высокий титр штамма Fa 4-1 B. subtilis наблюдался на питательных средах, где в качестве источников азота использовались кукурузный экстракт, азотнокислый натрий и пептон (2,3-5,4×108 КОЕ/мл). Тогда как штамм D 7-1 B. subtilis оказался более требовательным к источникам азота, хорошая плотность популяции данной бактерии отмечалась только с добавлением в качестве источника азота кукурузного экстракта (1,5×108 КОЕ/мл) (рис. 8).

Рисунок 8 – Рост штаммов бактерий-антагонистов рода Bacillus в зависимости от источников азота в процессе периодического культивирования

Fa 4-1 B. subtilis

D 7-1 B. subtilis
Практически непригодными для утилизации в качестве источников азотного питания для штамма Fa 4-1 B. subtilis оказался дрожжевой экстракт (4,3 × 106 КОЕ/мл), а для штамма D 7-1 B. subtilis – пептон (2,0 × 106 КОЕ/мл).
Для обоих штаммов флюоресцирующих бактерий был отмечен хороший рост при добавлении следующих азотсодержащих веществ: азотнокислый натрий, дрожжевой экстракт и пептон. Титр ЖК с указанными азотсодержащими веществами составлял от 7,7 × 1011 до 8,6 × 1012 КОЕ/мл (рис. 9) . Минимальный титр штаммов Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. отмечен на питательной среде с добавлением кукурузного экстракта (4,4-6,1 × 105 КОЕ/мл). Указанные биоагенты оказались неспособными использовать данный источник азота.
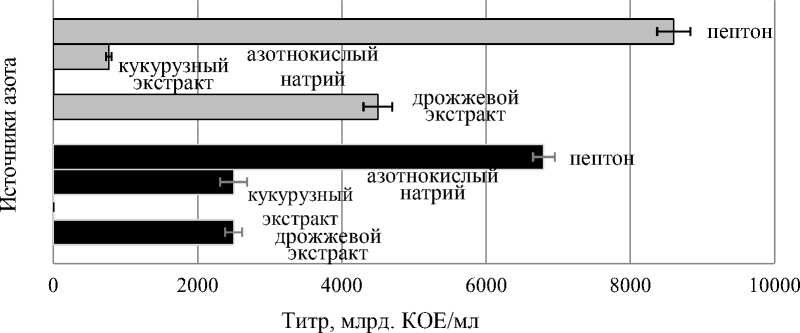
Sgrc-1 P. fluorescens Oif 2-1 Pseudomonas sp.
Рисунок 9 – Рост штаммов бактерий-антагонистов рода Pseudomonas в зависимости от источников азота в процессе периодического культивирования
Выводы. 1. Выявлен температурный оптимум для культивирования перспективных штаммов бактерий: Fa 4-1 B. subtilis – 30-35 °С, D 7-1 B. subtilis – 30 °С, Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. – 20-25 °С.
-
2. Определен оптимум рН для культивирования перспективных штаммов бактерий: D 7-1 B. subtilis – 6, Fa 4-1 B. subtilis , Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. – 6-8.
-
3. Установлены оптимальные источники углерода: для штамма Fa 4-1 B. subtilis меласса и глюкоза, для штамма D 7-1 B. subtilis – меласса и сахароза, для штаммов Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. – глюкоза, сахароза, меласса и глицерин.
-
4. Оптимальным источником азота для всех биоагентов был азотнокислый натрий, для штамма Fa 4-1 B. subtilis также пептон и кукурузный экстракт, для D 7-1 B. subtilis – дрожжевой и кукурузный экстракты, а для штаммов Sgrc-1 P. fluorescens и Oif 2-1 Pseudomonas sp. – пептон и дрожжевой экстракт.
-
5. Полученные экспериментальные данные впоследствии могут быть использованы для разработки элементов технологии производства микробиопрепаратов на основе перспективных штаммов бактерий родов Bacillus и Pseudomonas .
Работа выполнена при финансовой поддержке гранта РФФИ № 08-04-99010-р_офи.
