Фотопериодическая зависимость гинецея у разных генотипов сои

Автор: Зеленцов В.С., Цаценко Л.В., Мошненко Е.В.
Статья в выпуске: 2 (144-145), 2010 года.
Бесплатный доступ
Проведѐнные исследования показали зависимость угла наклона столбика пестика цветков сои от складываю-щихся длин дня в период формирования цветков, роста завязей и формирования бобов. Увеличение продолжительности фотопериода вызывает уменьшение угла α°НСП наклона столбика пестика. Формирование цветков и бобов в условиях укороченных фотопериодов относительно генетически детерминированного оптимума для каждого отдельно взятого генотипа сои определяет увеличение угла α°НСП. Выявленная зависимость между степенью адаптированности генотипа сои к фотопериоду и углом наклона столбиков пестиков позволяет рекомендовать еѐ в качестве визуального экспресс-метода предварительной оценки потенциальной реакции растений сои на длину дня и определения их принадлежности к северному или южному экотипу без организации широтных испытаний и использования расчѐтных методов исследования.
Соя, фотопериод, длина дня, завязь, столбик пестика, угол наклона
Короткий адрес: https://sciup.org/142150956
IDR: 142150956 | УДК: 58.035.2:633.853.52
Photoperiodic dependence of gynecium in different soybean genotypes
The carried out researches have shown dependence of inclination angle of pistil style in soybean flower from developing lengths of day in period of flower formation, ovary growth and beans formation. The increase in photoperiod duration causes re-duction of angle α°SPS inclination of pistil style. Flowers and beans formation in the conditions of the truncated photoperiods of the rather genetically determined optimum for each separately taken soybean genotype defines angle α°SPS increase. The revealed dependence between degree of adaptability of soybean genotype to photoperiod and inclination angle of pistil style allows to rec-ommend it as a visual express method of a tentative estimation of soybean plant potential reaction for day length and a determina-tion of its inhering to northern or southern ecotype without organization of latitudinal tests and use of computational methods of research.
Текст научной статьи Фотопериодическая зависимость гинецея у разных генотипов сои
Введение. Соя является фотопериодически высокочувствительной культурой и для перехода в репродуктивную стадию требует определен- ного соотношения освещения и темноты. Отклонения продолжительности светового дня от оптимума способствуют существенному изменению продолжительности вегетативного роста, а также времени наступления и длительности фаз цветения и созревания. Поэтому фотопериодическая чувствительность сои является одним из её главных адаптивных признаков [4, 10, 12].
Широкая изменчивость по фотопериодическим реакциям генофонда сои, интенсивная адаптивная селекция сортов, приспособленных к различным эколого-географическим условиям, а также и их активная интродукция позволили распространить возделывание этой культуры практически на все материки, кроме Антарктиды. В настоящее время зона распространения культурной сои охватывает почти сплошную полосу от 58-60° с. ш. (Канада, Россия, Швеция) до 48-50° ю. ш. (Аргентина), сформировав практически непрерывный диффузный ареал вторичного формообразования, опоясывающий весь земной шар [9].
Многочисленными исследованиями установлено, что изменение длины дня оказывает большое влияние как на вегетационный период в целом, так и отдельно на фазы цветения, формирования бобов и налива семян; количество и ярусную изменчивость междоузлий; высоту растений и их продуктивность [2, 4, 10, 12, 13, 17, 20, 22]. Во всех случаях успешное возделывание отдельно взятого сорта сои возможно только в достаточно узком диапазоне длин дня и географических широт, как правило, не превышающем 1° [1, 3, 19].
Поэтому для увеличения широтного ареала рентабельного возделывания сортов сои необходимы исследования, направленные на изучение реакции генотипов сои на изменяющуюся длину дня и создание сортов, пригодных для возделывания в широком спектре эколого-географических условий.
Полевые испытания сои на различных географических широтах требуют значительных средств и ресурсов на закладку и агротехноло-гическое сопровождение полевых питомников, а также наличия подготовленного научно-исследовате-льского персонала на местах. Как дополнение к полевым исследованиям, сорта сои нередко оценивают в контролируемых условиях искусственного климата. Однако условия искусственного освещения зачастую далеки от естественных из-за разницы в интенсивности и спектральном составе солнечного и искусственных источников света. Кроме этого, существенно ограничивает использование сооружений искусственного климата для оценки реакции сои на длину дня очень высокая энергоемкость тепличного оборудования [15, 16, 18, 21, 23].
В последние годы появились работы отдельных отечественных ученых, указывающие на возможность косвенного определения фотопериодической адаптивности сои по ярусной изменчивости длин междоузлий растений сои, степени фертильности пыльцы, пыльцевой продуктивности, а также по линейным размерам морфологических элементов цветка [6, 7, 14].
Тем не менее имеющихся данных все еще недостаточно для высокоэффективной работы по созданию фотопериодически слабочувствительных или фотонейтральных сортов сои. В связи с этим проведение исследований, направленных на поиск и разработку малозатратных экспресс-методов оценки средовой и генотипической реакции генотипов сои на изменяющуюся длину дня, не требующих дорогостоящих широтных или тепличных испытаний селекционного материала, сохраняет свою актуальность.
Материалы и методы . Изучение фотопериодически зависимого наклона столбика пестиков в цветках сои проводили на 13 сортах с различным вегетационным периодом, включая очень ранние сорта северного экотипа: Светлая, Магева, Касатка, Окская, Ланцетная и Рось, а также ранние, средние и поздние сорта южного экотипа: Лира, Альба, Славия, Белогорская, Вилана, Трембита и Лиана. Кроме этого изучали 3 расщепляющиеся гибридные комбинации F 2 Брянская МИЯ х Вилана, Лира х Магева и Белогорская х Ланцетная с участием в каждой комбинации родительских форм одновременно южного и северного экотипов. Все сорта выращивали в полевых условиях Краснодара (45 ° с. ш., максимальная длина дня 15 ч 37 мин), г. Алексеевка Белгородской области (50 ° с. ш., максимальная длина дня 16 ч 26 мин), а также в камере искусственного климата при фотопериоде 12 ч и интенсивности освещения 8-12 тыс. лк. В условиях искусственного климата все сорта выращивали в почвеннопесчаной смеси в сосудах ёмкостью 10 дм2 по 2 растения на сосуд.
Посев сортов в полевых условиях осуществляли в оптимальные сроки (I декада мая). В период цветения на каждом сорте для морфологического анализа из нижних ярусов растений за сутки до растрескивания пыльников и в день самоопыления рендомизированно отбирали по 10 цветков. Цветки фиксировали и хранили в фиксаторе Кларка. Морфологический анализ и микрофотографирование пестиков цветков исследуемых сортообразцов осуществляли при увеличении х 62,5 после их окрашивания ацетолакмоидом по методике З.П. Паушевой [11]. Кроме этого, в фазы полного налива семян и физиологического созревания в нижнем ярусе растений каждого сорта рен-домизированно отбирали по 10 бобов и фотографировали их апикальную часть.
Угол наклона α°НСП столбиков пестиков в цветке и их засохших остатков у сформированных плодов определяли по пространственному положению верхней части столбика, расположенной непосредственно под рыльцем, к продольной оси завязи или сформировавшегося боба.
Для количественной оценки фотопериодической адаптивности исследуемых сортов использовали коэффициент завершенности вегетативного роста К сс [6]. Этот коэффициент определяли через частное разностей между длиной самого длинного ( Yx max ) и последнего ( Yx n ) междоузлия и между длиной самого длинного и первого ( Yx 1 ) междоузлия растения сои. Значения коэффициента К сс расположены в диапазоне от -1 до +1. Близкие к 0 значения этого коэффициента соответствуют фотопериодической реакции ранней группы генотипов сои, возделываемых на южной границе оптимальных по фотопериодам широт. Положительные значения Ксс соответствуют очень ранним генотипам, адаптированным к более продолжительным фотопериодам по сравнению со складывающимися в месте выращивания. Слабоотрицательные (от -0,1 до -0,3) значения коэффициента К сс имеют среднеспелые сорта сои, наиболее полно адаптированные к складывающимся в данных условиях длинам дня. Средне- и сильноотрицательные значения Ксс свойственны очень поздним в данных условиях среды генотипам, адаптированным к более низким географическим широтам с короткими фотопериодами [6, 8].
В качестве контрольной оценки фотопериодической реакции гинецея исследуемых сортообразцов использовали морфологический метод предварительной оценки генотипов сои, основанный на сравнении сумм длин трех нижних и трех верхних междоузлий L1:L2 [6, 8]. Согласно описанию этого метода, если сумма длин верхних междоузлий (L2) примерно равна или до двух раз превышает сумму нижних междоузлий (L1), такое растение пред- варительно следует отнести к адаптированным к данному фотопериоду. Если визуальная сумма длин верхних междоузлий меньше визуальной суммы длин нижних междоузлий – растение относится к длиннодневным и адаптировано к более высоким широтам. Если отношение сумм длин верхних междоузлий явно больше двойной суммы нижних междоузлий – растение относится к короткодневным и адаптировано к более низким широтам.
Результаты и обсуждение. Визуальные наблюдения за пространственной конфигурацией элементов в цветках сортов сои, выращенных при различных фотопериодах, позволили обнаружить реакцию геницея на укороченный фотопериод в виде изменения угла наклона столбика (α° НСП ) (рис. 1).
В оптимальных по длине дня для отдельно взятого генотипа сои условиях в развивающемся бутоне столбик сначала согнут так, что рыльце примыкает к завязи и прижато к ней. Оно практически недоступно в этот период для чужеродной пыльцы, попавшей вовнутрь цветка, например, при помощи насекомых – трипсов или пчел. При полном созревании яйцеклетки и готовности завязи к опылению (примерно за сутки до растрескивания пыльников) столбик постепенно начинает выпрямляться, отдаляясь от боковой поверхности завязи (рис. 2).
К моменту созревания пыльцы столбик в оптимальных по длине дня условиях выпрямляется под острым углом к продольной оси пестика. При этом рыльце оказывается обращенным наружу и располагается на одном уровне с тесно окружающими его пыльниками тычиночной трубки. При растрескивании пыльников пыльца сразу же попадает на рыльце и прорастает.
В условиях излишне короткого фотопериода столбик пестика сохраняет большой угол наклона, или процесс его выпрямления остается незавершенным. Поэтому на момент растрескивания созревших пыльников рыльце оказывается обращенным к стенке завязи, оставаясь малодоступным для собственной пыльцы, уменьшая, таким образом, вероятность самоопыления или искусственного переопыления посредством насекомых пыльцой другого сорта.
Модельный эксперимент с участием адаптированных к разным фотопериодам сортов сои в камере искусственного климата, а также в полевых условиях г. Краснодара и г. Алексеевки Белгородской области подтвердил выявленную зависимость угла наклона столбика пестика
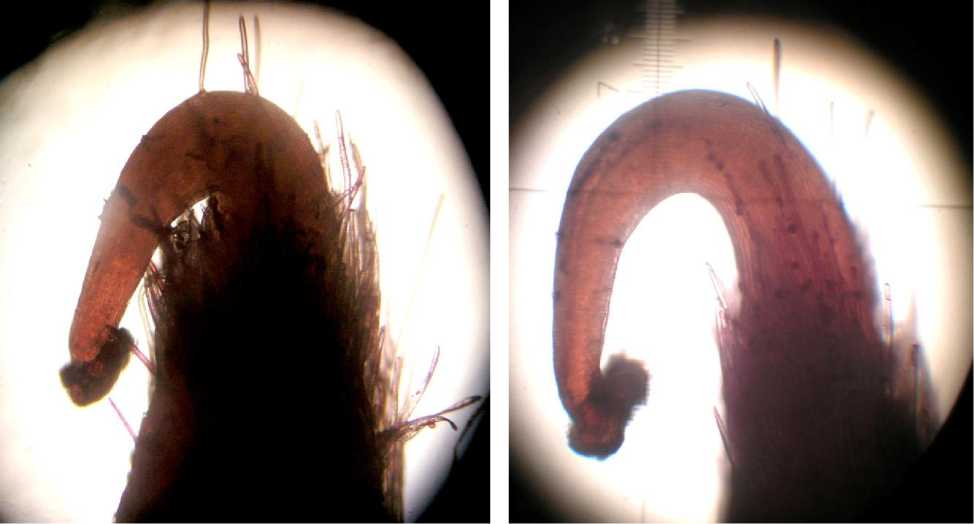
Фотопериод 12 ч
Фотопериод 16 ч
Рисунок 1– Форма столбика пестика за сутки до самоопыления в зависимости от продолжительности фотопериода, сорт Трембита, камера искусственного климата, увеличение х 62,5
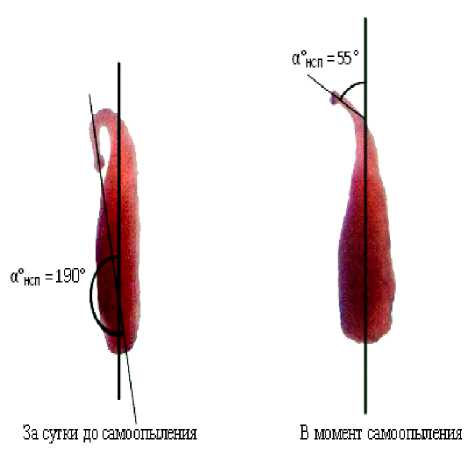
Рисунок 2 – Угол наклона столбика пестика (α° НСП ) в зависимости от возраста цветка, сорт Трембита
(угол α° НСП ) от продолжительности фотопериода в период формирования цветков (рис. 3, табл. 1).
Удлинение продолжительности дня в период формирования цветков с 12 до 16,5 ч приводило к уменьшению угла α°НСП. При этом были выявлены генотипические различия по реакции признака «угол α° НСП » на продолжительность фотопериода. Так, если у всех сортов сои при длине дня 12 ч угол α° НСП был примерно равным, то при длине дня 15,5 ч у выведенных на широте 45° сортов южного экотипа этот угол уменьшался, при этом поверхность рыльца разворачивалась к окружающим пестик пыльникам. У длиннодневных сортов северного экотипа при длине дня 15,5 ч угол α° НСП изменялся незначительно по сравнению с 12-часовым фотопериодом. Увеличение продолжительности фотопериода до 16,5 ч у сортов сои краснодарской селекции вызывало дальнейшее уменьшение угла α° НСП . У длиннодневных сортов изгиб столбика также уменьшался. Угол наклона становился близким к таковым у южных сортов при фотопериоде 15,5 ч.
Сорта сои, адаптированные к макс. длине дня 15,5 часов
(на примере сорта Вилана)
Сорта сои, адаптированные к макс. длине дня 16,5 и более часов
(на примере сорта Магева)



г. Алексеевка, максимальная длина дня – 16,5 ч
Камера искусственного климата, длина дня – 12 ч
г. Краснодар, максимальная длина дня – 15,5 ч
Рисунок 3 – Влияние длины дня на угол наклона столбика пестика (α° НСП ) на момент самоопыления у различающихся по фотопериодической чувствительности генотипов сои, 2008 г.
Таблица 1 – Зависимость углов наклона столбиков пестиков южных и северных экотипов сои от продолжительности фотопериода
Краснодар-Алексеевка, 2008 г.
|
Сорт сои |
Период вегетации на широте Краснодара при оптимальном сроке сева, сутки |
Угол наклона столбиков пестиков (α°НСП) в фазе цветения при максимальном фотопериоде: |
Угол α° НСП у зрелых бобов при фотопериоде 15,5 ч** |
||
|
12 ч* |
15,5 ч** |
16,5 ч*** |
|||
|
Сорта сои южного экотипа, адаптированные к широте 45° |
|||||
|
Вилана |
117 |
151 |
136 |
86 |
48 |
|
Лира |
90 |
171 |
122 |
115 |
69 |
|
Альба |
113 |
161 |
150 |
108 |
78 |
|
Славия |
98 |
195 |
159 |
125 |
65 |
|
Белогорская |
98 |
184 |
173 |
148 |
69 |
|
Среднее по группе |
103 |
172,4 |
148,0 |
116,4 |
65,8 |
|
Сорта сои северного экотипа, адаптированные к широтам > 50° |
|||||
|
Светлая |
82 |
176 |
164 |
136 |
113 |
|
Магева |
87 |
199 |
174 |
140 |
118 |
|
Касатка |
82 |
213 |
198 |
156 |
113 |
|
Окская |
87 |
230 |
204 |
165 |
135 |
|
Ланцетная |
82 |
221 |
186 |
176 |
140 |
|
Среднее по группе |
84 |
207,8 |
185,2 |
154,6 |
123,8 |
|
Среднее по опыту |
94 |
190,1 |
166,6 |
135,5 |
94,8 |
* – в камере искусственного климата; ** – в полевых условиях, г. Краснодар;
*** – в полевых условиях, г. Алексеевка Белгородской обл.
Как следует из данных, представленных в таблице 1, среднее по опыту значение угла а° НСП относительно продольной оси пестиков в условиях 12-часового фотопериода составило 190°. В условиях 15,5-часового фотопериода этот показатель сократился до 167°, а при 16,5часовом фотопериоде ещё уменьшился до 64,8°. Однако при этом выявлены групповые различия по углу а° НСП между сортами южного и северного экотипов.
Так, среднегрупповой угол а° НСП у сортов сои южного экотипа в условиях 12-часового фотопериода составил 172° с варьированием этого показателя у отдельных сортов от 151 до 195°. У группы сортов северного экотипа угол до а° НСП в среднем составлял 207,8° с варьированием значений α° от 176 до 230°. Увеличение продолжительности дня до 15,5 ч привело к уменьшению угла а° НСП до 148°, а у раннего в условиях Краснодара сорта Лира - до 122°. У сортов северного экотипа этот показатель в среднем составлял 185,2°.
В условиях наиболее продолжительного в опыте фотопериода (16,5 ч) угол а° НСП еще уменьшился и составил 116,4°, а у среднеспелого сорта Вилана даже 86°. У сортов северного экотипа угол наклона столбика также уменьшился и составил 154,6°, что оказалось близким (а° НСП = 148°) к среднегрупповым значениям этого показателя у сортов южного экотипа при 15,5часовом фотопериоде.
Полевые наблюдения показали, что у некоторых бобов столбик сохраняется вплоть до их полного созревания. При этом у сортов южного экотипа среднегрупповое значение угла а° НСП составляло всего 65,8° с варьированием этого показателя между сортами от 48 у сорта Вилана до 78° у сорта Альба. У длиннодневных сортов северного экотипа значения угла а° НСП также уменьшились и, в среднем, составили 123,8° с варьированием от 113 у сорта Светлая до 140° у сорта Ланцетная (см. табл. 1). Уменьшение угла а° НСП у зрелых бобов обеих групп сортов по сравнению с этим же показателем в фазе растрескивания пыльников позволяет сделать предположение о продолжении процесса распрямления столбика пестика еще некоторое время после самоопыления цветка.
У части зрелых бобов независимо от генотипа столбик отсутствовал, не позволяя определить угол α°НСП. По всей вероятности причиной этому были неблагоприятные условия внешней среды, в частности, пониженная влажность воздуха, вызывающая сверхкрити- ческую потерю влаги из тканей столбиков и их последующее засыхание и опадение. Не исключена также вероятность механического обламывания столбиков при прорастании верхней части завязи сквозь засохший после цветения венчик цветка или при взаимном трении созревших растений сои относительно друг друга в ветреную погоду. Визуальный анализ бобов в полевых условиях показал, что в засушливых условиях процесс засыхания столбиков может развиваться уже с фазы налива семян. Тем не менее наблюдения за углами а°НСП отдельно взятых сортообразцов в фазы бобообразования, налива семян, а в отдельных случаях даже в фазе физиологического созревания, являются весьма информативными, позволяя сделать предварительные выводы о фотопериодической чувствительности сортов (рис. 4).
В связи с этим нами было предположено, что выявленная закономерность может быть использована в качестве визуального экспресс-метода предварительного полевого определения адаптированности к фотопериоду генотипов сои по углу а° НСП у бобов.
Подавляющая часть известных методов оценки фотопериодической чувствительности генотипов сои и их адаптированности к определенным длинам дня, как правило, связана с широтными посевами или со сложными математическими исчислениями на основе особенностей динамики ярусной изменчивости длин междоузлий растений с различной реакцией на длину дня. В связи с этим селекционеры испытывают необходимость в простых методах полевой оценки фотопериодической чувствительности генотипов сои, в том числе, при индивидуальном отборе растений в гибридных питомниках. Для предварительной оценки адаптированности генотипов сои к фотопериоду в расщепляющихся гибридных популяциях было предложено использовать обнаруженное нами явление различных углов наклона столбиков пестиков у длиннодневных сортов сои северного экотипа и сортов южного экотипа, адаптированных к широте Краснодара.
С этой целью в 2009 г. в гибридном питомнике отдела сои ВНИИМК были подобраны и исследованы три гибридные комбинации F2 с участием родительских форм северного и южного экотипов: Брянская МИЯ х Вилана, Лара х Магева и Белогорская х Ланцетная. Одновременно признак «угол α° НСП » был оценен у исходных родительских форм: Вилана, Белогорская, Лира, Брянская МИЯ, Ланцетная и
Магева – в условиях искусственного климата при фотопериодах 12 и 16 ч (табл. 2).
Таблица 2 – Некоторые статистические параметры углов наклона столбиков (α° НСП ) у бобов длиннодневных и короткодневных родительских генотипов сои, формирующихся при разных фотопериодах
Камера искусственного климата, 2009 г.
|
Сорт |
Фотопериод 12 ч |
||
|
n * |
X ** |
σ *** |
|
|
Вилана |
66 |
165,0 ± 10,9 |
45,1 |
|
Белогорская |
51 |
169,5 ± 11,8 |
42,9 |
|
Лира |
57 |
175,9 ± 4,9 |
18,7 |
|
Брянская МИЯ |
54 |
180,2 ± 5,6 |
20,8 |
|
Ланцетная |
47 |
154,3 ± 11,5 |
41,4 |
|
Магева |
43 |
173,5 ± 5,7 |
19,0 |
|
Фотопериод 16 ч |
|||
|
Вилана |
n |
X |
σ |
|
Белогорская |
44 |
85,0 ± 6,5 |
22,1 |
|
Лира |
31 |
84,3 ± 10,2 |
23,2 |
|
Брянская МИЯ |
63 |
123,2 ± 5,4 |
21,7 |
|
Ланцетная |
36 |
141,7 ± 13,6 |
41,6 |
|
Магева |
55 |
118,2 ± 9,3 |
35,2 |
|
80 |
164,4 ± 4,1 |
14,5 |
|
* – выборка бобов, шт.;
** – средний угол α°НСП по выборке;
*** – стандартное отклонение
Анализ данных таблицы 2 показывает, что все родительские формы в условиях короткого 12-ча-сового фотопериода характеризовались большими углами α° НСП . Их выращивание при 16часовом фотопериоде закономерно сопровождалось некоторым уменьшением угла наклона столбика.
У сортов северного экотипа этот угол уменьшился на 28°, или на 16 %. При этом минимальная чувствительность признака «угол α° НСП » на увеличение продолжительности фотопериода отмечена у сорта Магева, при фотопериоде 16 ч уменьшившаяся всего на 5 % от этого показателя при длине дня 12 ч. Максимальная реакция на увеличение фотопериода отмечена у сорта Ланцетная. Углы наклона столбиков у бобов этого сорта уменьшились относительно 12-часового фотопериода на 23 %. Сорт Брянская МИЯ по этому показателю занимал промежуточное между этими сортами положение.
Реакция признака «угол наклона столбика» у сортов, выведенных на широте 45° и адаптированных к длинам дня юга России, отличалась от таковой у длиннодневных сортов северного экотипа.
Так, если угол α° НСП при фотопериоде 12 ч в среднем по группе сортов южного экотипа составлял практически те же 170°, что и в группе сортов северного экотипа, то при фотопериоде
16 ч среднее значение этого показателя уменьшилось более чем на 70°, или на 43 %.
Так, у самого раннего в группе сортов южного экотипа сорта Лира угол наклона столбика уменьшался на 30 %. А у раннего сорта Белогорская и среднего сорта Вилана этот же показатель уменьшился практически вдвое. При этом средние значения углов α° НСП у этих сортов составили 84 и 85°, что составляет 50 и 52 % соответственно от исходных углов наклона столбиков этих сортов при 12-часовом фотопериоде.
Фенологические наблюдения и морфологические измерения гибридных растений в расщепляющихся популяциях F2, полученных с участием сортов северного и южного экотипов, показали, что основные показатели фотопериодической адаптации, такие как вегетационный период и высота растений, имеют непрерывный вариационный ряд изменчивости между ранним и поздним, а также между низким и высоким родительскими сортами. В связи с этим дополнительные исследования фотопериодической чувствительности проводили на искусственно сформированных группах растений, занимающих крайние полярные позиции в пределах каждой гибридной популяции, по признакам вегетационный период и высота растений.
В отобранных группах растений оценивали вегетационный период, средний угол наклона столбика α° НСП , а также вычисляли такие параметры фотопериодической чувствительности, как коэффициент завершенности вегетативного роста К сс и отношение сумм длин 3-х нижних и 3-х верхних междоузлий центральных побегов индивидуальных растений. Полученные данные представлены в таблицах 3 и 4.
Анализ данных, представленных в таблице 3, показывает, что средняя продолжительность вегетационного периода у ранних гибридных растений в разных гибридных популяциях была близка и составляла от 88-90 до 91-95 суток. Однако этот же показатель у наиболее поздних растений варьировал более существенно: от 100105 суток в гибридной популяции, полученной от скрещивания очень раннего сорта южного экотипа Лира и очень раннего сорта северного экотипа Магева до 106-110 суток при использовании в качестве родительской формы сорта Белогорская с более продолжительным периодом вегетации, что привело к закономерному увеличению вегетационного периода поздней группы гибридных растений. Соответственно, включение в скрещивание среднего сорта Вилана еще больше удлинило вегетационный период этой группы растений, в условиях 2009 г. составивший 115-120 суток.
Таблица 3 – Некоторые параметры фотопериодической адаптивности гибридных популяций сои при оптимальных (майских) сроках сева на широте 45°
Краснодар, 2009 г.
|
Гибридная комбинация |
Отношение к макс. длине дня на широте г. Краснодара |
Период вегетации, сутки |
Средний угол α°НСП |
Коэффициент завершённости вегетативного роста, K cc |
Оценка сои по реакции на фотопериод по соотношению сумм длин 3-х нижних и 3-х верхних междоузлий растений |
||
|
L 1, мм |
L 2, мм |
L 2 : L 1 |
|||||
|
Брянская МИЯ х Вилана |
Излишне короткая |
90-95 |
196 |
0,09 |
81 |
74 |
0,92 |
|
Оптимальная |
115-120 |
77 |
-0,29 |
86 |
163 |
1,89 |
|
|
Лира х Магева |
Излишне короткая |
85-90 |
165 |
0,11 |
79 |
69 |
0,87 |
|
Оптимальная |
100-105 |
82 |
-0,26 |
83 |
123 |
1,48 |
|
|
Белогорская х Ланцетная |
Излишне короткая |
85-90 |
182 |
0,16 |
69 |
59 |
0,86 |
|
Оптимальная |
105-110 |
86 |
-0,18 |
92 |
143 |
1,55 |
|
Таблица 4 – Углы наклона столбиков у бобов гибридных растений F2 с разной фотопериодической адаптивностью
Краснодар, 2009 г.
|
Гибридная комбинация |
Отношение к макс. длине дня на широте г. Краснодара |
Угол наклона столбиков у бобов растений F 2 в фазы налив семян–созревание |
|
Брянская МИЯ х Вилана |
излишне короткая (северный экотип) |
жжм |
|
оптимальная (южный экотип) |
||
|
Лира х Магева |
излишне короткая (северный экотип) |
|
|
оптимальная (южный экотип) |
^^^ |
|
|
Белогорская х Ланцетная |
излишне короткая (северный экотип) |
^^^|^^^^^ ^^Ц^^^^^^ ^|^^^^^ |
|
оптимальная (южный экотип) |
Вычисления коэффициентов завершенности вегетативного роста (см. табл. 3) у анализируемых групп гибридных растений в пределах гибридных популяций показали, что у ранних форм они приобретали слабоположитель-ные( К сс = 0,09-0,16) значения, а у самых поздних растений в пределах каждой из гибридных комбинаций – слабоотрицательные ( Ксс = от -0,18 до -0,29) величины.
Применение эмпирического морфологического метода предварительной оценки генотипов сои по реакции на фотопериод, основанного на методике сравнения сумм длин трех нижних и трех верхних междоузлий [8], также показало различия между группами гибридных растений в исследуемых гибридных популяциях.
В наших исследованиях группы ранних растений во всех трех гибридных комбинациях имели искомое отношение длин L 1 и L 2 менее единицы, и следовательно, они должны быть отнесены к категории длиннодневных. Все растения поздней группы спелости имели соотношение длин L 1 и L 2 более единицы (от 1,48 до 1,89) и являлись наиболее адаптированными к фотопериодам, складывающимся в данных условиях выращивания.
Оценка углов наклона столбиков α° НСП у исследуемых групп гибридных растений показала высокую положительную взаимосвязь между этим показателем и расчетными параметрами фотопериодической чувствительности. Так, ранние формы, на основе значений коэффициентов К сс и соотношения длин верхних и нижних междоузлий отнесенные к длиннодневным, характеризуются большими значениями угла α° НСП , варьирующими от 165 до 196°. Тогда как у растений с фотопериодической чувствительностью, наиболее адаптированной к выращиванию на широте Краснодара, углы α° НСП были более чем в 2 раза меньше и варьировали от 77 до 86°. Эта разница легко визуально различима в полевых условиях на бобах, нередко вплоть до их созревания, даже без дополнительных измерений углов наклона столбиков (см. табл. 4).
Выводы. Таким образом, проведенные исследования показали зависимость угла наклона столбика пестика в цветках сои от складывающихся длин дня в период формирования цветков, роста завязей и формирования бобов. Увеличение продолжительности фотопериода вызывает уменьшение угла α°НСП наклона столбика пестика. Формирование цветков и бобов в условиях укороченных фотопериодов относительно генетически детерминировано-го оптимума для каждого отдельно взятого генотипа определяет увеличение угла α°НСП. Выявленная зависимость между степенью адаптированности генотипа сои к фотопериоду и углом наклона столбиков пестиков позволяет рекомендовать ее в качестве информативного визуального экспресс-метода предварительной оценки потенциальной реакции растений сои на длину дня и установления их принадлежности к северному или южному экотипу без организации широтных посевов и использования расчетных методов исследования.
