Genetic Portraits of the Yakut Regional Populations (Based on Y-Chromosome Polymorphism Data)
")
Автор: Adamov D.S., Potanina A.Y., Bogunov Y.V., Zhabagin M.K., Balanovska E.V.
Журнал: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Рубрика: Anthropology and paleogenetics
Статья в выпуске: 2 т.53, 2025 года.
Бесплатный доступ
We present data on SNP haplogroups and STR haplotypes of the Y-chromosome in main Yakut ethno-local populations. Clusters of STR haplotypes and median networks of principal haplogroup N3a2 (N-M1982) indicate three lines: N-M1983, N-M1933, and N-Y25011. The first one, widely distributed throughout Yakutia and present in one half of Yakut males, marks the putative descendants of Elley, the legendary progenitor of the Yakuts. Line N-M1933 is found where descendants of another legendary progenitor, Omogoy, live. The N-Y25011 line is frequent in the Vilyuy basin, where the Yakuts had migrated from the Middle Lena in the 1600s. The predominance of lines N-M1983 and N-Y25011 is less marked in the northeastern part of this region than in the southwestern one. The frequency of haplogroup N3a2 drops to 70 % in the Yakuts of the northeastern Vilyuy basin, northeastern Yakutia, and northeastern Khabarovsk Territory. The estimated time to the most recent common ancestor of Yakuts, marked by the N3a2 haplogroup, 1200 ± 480 years, shows a fair agreement with aDNA data. Samples of the Yakuts from Yakutia and the Okhotsk coast and of Sakha-speaking Evenki reveal a compact “Yakut” core area in the multidimensional genetic space of the Y-gene pools of Siberia and the Far East. This core is marked by an elevated frequency (68–95 %) of the N3a2 haplogroup, which is rare (0–21 %) in other populations of Siberia (Dolgans, Nanais, Negidals, Nivkhs, Tuvans, Ulchi, Evenki, and Evens). Regional Y-chromosome profiles are consistent with historical data regarding Yakut origins and legends.
Gene pool, Yakuts, Y-chromosome, haplogroups, STR haplotypes, median networks
Короткий адрес: https://sciup.org/145147501
IDR: 145147501 | DOI: 10.17746/1563-0110.2025.53.2.144-154
Текст научной статьи Genetic Portraits of the Yakut Regional Populations (Based on Y-Chromosome Polymorphism Data)
The problem of the origin of the Yakuts has been studied for over 300 years by historians, archaeologists, ethnographers, and linguists. Numerous studies have formulated various hypotheses and made significant progress in understanding this issue; however, a definitive solution has yet to be found. Therefore, data from population genetics can serve as an additional source when discussing the questions of the origin and settlement of the Yakuts.
Russian pioneers who established Fort Lensky (Yakutsk) in 1632, encountered approximately 28,000 Yakuts (self-designation – Sakha) (Dolgikh, 1960: 542–544). The economic lifestyle of the Sakha (northern herders and horse breeders) was markedly different from that of their closest neighbors, whose way of life was based on reindeer herding, hunting, fishing, and gathering. Their language also differed, belonging to the Turkic language group. No written records about the Yakuts have been found till the early 17th century, but oral folk tradition
points to the names Elley Bootur and Omogoy Baai as the ancestral founders of the Sakha (Ksenofontov, 1977: 16–28). The ancestors of the few Khorinsky clans are considered to be the elder Uluu Khoro and his relatives (Istoriya Yakutii, 2020: Vol. I, p. 198).
By the beginning of annexation of the territory of modern Yakutia to the Russian State (17th century), more than 90 % of the Yakuts were concentrated in its central part, exactly in the interfluve of the Lena and Aldan rivers (Istoriya Yakutii, 2021: Vol. II, p. 9). Over the next two centuries, they settled across a vast territory: the area of the Republic of Sakha (Yakutia) is about 3 million km², by 2021 the population of the Yakuts in this region reached 470,000 people, traditionally divided into central, Vilyuy, and northern groups (Istoriya Yakutii, 2020: Vol. I, p. 251).
One of the genetic tools that allows us to look back through the centuries is the comprehensive analysis of Y-chromosome polymorphism. Since the Y-chromosome is present in the cell nucleus only in males, there is virtually no shuffling of genetic material between parental genomes, and a significant portion of the Y-chromosome is passed down unchanged from father to son across generations. Rare mutations, gradually accumulating, give rise to a global genealogical tree of the Y-chromosome, where the genome of each man occupies its own place. This tree arises from single nucleotide polymorphisms (SNPs)—nucleotides substitutions. Its branches, of varying thickness and antiquity, are reflected in a strict hierarchical system of haplogroups at different levels. For historical reconstructions, mutations of another type, known as short tandem repeats (STRs), are also widely used. STR haplotypes differ from one another by the varying number of repeats of the same “motifs” (combinations of two to six nucleotides). The probability of STR mutations (increases or decreases in the number of repeated “motifs”) is much higher than that of single-nucleotide mutations. The approach to the mutational process as “molecular clocks” is based on mathematical models of probability theory, and allows for probabilistic estimates of the age of genetic events. If the rate of SNP mutations in “molecular clocks” is likened to the movement of the hour hand, then the rate of STR mutations is akin to the movement of the second hand. The use of both allows for reconstructing the timing of genetic events with varying degrees of accuracy.
All studies of the Y-gene pool of the Yakuts indicate a striking feature: an extremely low diversity of Y-chromosome haplogroups, with the dominance of the N-Tat haplogroup (Zerjal et al., 1997; Puzyrev et al., 2003), whose frequency reaches 80–90 % among the Yakuts (Tambets et al., 2004; Pakendorf et al., 2006;
Kharkov et al., 2008; Fedorova, 2008: 139), exhibiting a pronounced founder effect. Time to the most recent common ancestor (TMRCA) of modern carriers of this Y-chromosome variant is ca 1300 BP (Adamov, 2022).
In a number of studies (Pakendorf et al., 2006; Fedorova, 2008: 139–154; Fedorova et al., 2014), the main Yakut ethno-local populations were investigated— central, Vilyuy, and northern. The proximity of the first two was identified, as well as a decrease in the frequency of the main haplogroup N-Tat among northern Yakuts (about 70 %) as compared to central and Vilyuy Yakuts (80–90 %).
The study of ancient DNA from anthropological materials of Yakut burials has confirmed the continuity and relative stability of Y-chromosome lineages over the past 400 years (Fedorova, Zvenigorosky, Alekseev, 2023). Analysis of 17-marker Y-STR haplotypes revealed three dominant haplotypes in ancient and modern Yakut populations: the most frequent Ht1 is present in all groups; Ht2 is prevalent in the central part of Yakutia; and Ht3 predominates in the Vilyuy region.
Following the human genome sequencing and the rapid development of whole-genome sequencing technologies, new opportunities have arisen for studying the genetic history of the Yakuts (Poznik et al., 2013; Lippold et al., 2014; Karmin et al., 2015; Ilumäe et al., 2016). It has been established that the N-M2019/M2118 lineage emerged on the phylogenetic tree of the parent N-Tat haplogroup ca 6200 BP (YFull..., 2023). The Yakut lineage N-M1982 separated from other branches of N-M2019/ M2118 haplogroup about 2300 BP (Adamov, 2022). It is divided into two branches of unequal frequency: the major N-M1991 and the minor N-M1933 (Karmin et al., 2015). The former, in turn, consists of subbranches N-M1983 and N-Y25011 (Adamov, 2022).
Owing to clear correlations between branches N-M1983, N-Y25011, N-M1933 and combinations of haplotypes at STR loci, there is an opportunity to investigate various territorial populations of the Yakuts in more detail (Ibid.). The integration of Y-chromosome data obtained by research teams with new findings allows for a more precise reconstruction of the genetic history of the Yakuts.
Material and methods
Studied samples. A total of 758 Y-STR haplotypes from Yakut samples from Yakutia were selected for analysis: 135 from our own data (hereinafter referred to as o.d.), and 623 from publications by other authors. For 561 samples (121 o.d., 440 from literature data), the geographical origin of the donors is known (see Fig. 1). The grouping of samples by regions depended
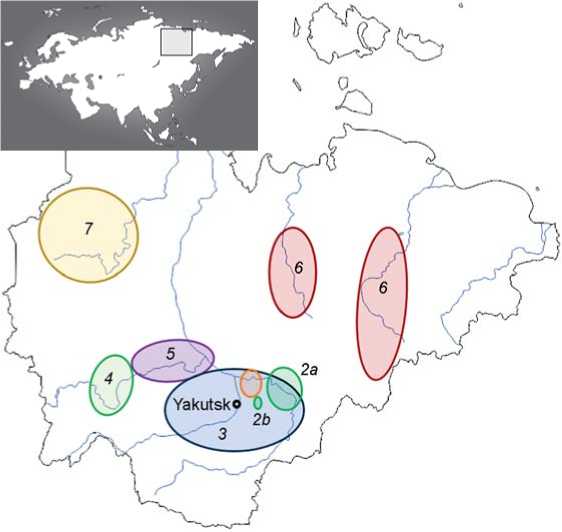
Fig. 1 . Geographic location of the analyzed territorial groups of Yakuts. 1 – Ust-Aldansky ulus (after (Kharkov et al., 2008)); 2a – Tattinsky and Tomponsky uluses; 2b – the villages of Tolon and Chyappara in Churapchinsky ulus (after (Karafet et al., 2018)); 3 – central Yakutia (after (Thèves et al., 2010)); 4 – southwestern Vilyuy basin (after (Zvénigorosky et al., 2020)); 5 – northeastern Vilyuy basin (after (Xu et al., 2014)); 6 – northeastern Yakutia (Verkhoyansky, Oymyakonsky, Momsky, Abyisky uluses); 7 – Olenek Sakha-speaking Evenki.
phenol-chloroform extraction method with proteinase K. Genotyping (fragment analysis) was carried out on an ABI 3130xl instrument (Applied Biosystems) with the Y-filer PCR Amplification Kit (Applied Biosystems) (a 17-locus panel of Y-chromosome STR markers). Four samples of buccal epithelium were genotyped in a commercial laboratory FTDNA (12 STR loci).
Determination of haplogroups. Most samples used in this study had information on the Y-SNP haplogroup, with varying degrees of detail regarding its position on the Y-chromosomal haplogroup tree. For 158 samples, which were provided only with Y-STR haplotype data (Thèves et al., 2010; Xu et al., 2014), the haplogroup was predicted based on Y-STR haplotypes. The haplogroups of these samples were established by comparing them with haplotypes from other studies where haplogroups had been tested. In rare cases, the online predictor Nevgen was used.
Multidimensional scaling. Nei’s genetic distances were calculated using the DJgenetic program (Balanovsky et al., 2008) based on the frequency of 13 Y-haplogroups: N3a2 (N-M2019/M2118), C-M407, C-M48, C-M217(xM504, M407, M48), N-F4205, on the sample size. The Vilyuy Yakuts are divided into two territorial groups: the southwestern Vilyuy basin – 48 samples (o.d.) from the Suntarsky and Nyurbinsky uluses; northeastern Vilyuy basin – 109 samples from the Vilyuysky, Verkhnevilyuysky, and Kobyaisky uluses (17 o.d., 92 (Thèves et al., 2010)). The Yakuts of northeastern Yakutia are represented by 89 samples: from Abyisky (3 o.d.), Verkhoyansky (1 o.d.), Momsky (50 (Zvénigorosky et al., 2020)), and Oymyakonsky (35 (Ibid.)) uluses. The group of central Yakutia includes 176 samples from the Ust-Aldansky ulus (6 o.d., 108 (Kharkov et al., 2008), 62 (Karafet et al., 2018)); 78 from the Tattinsky (6 o.d.), Tomponsky (4 o.d.) uluses, and western Churapchinsky (41 (Thèves et al., 2010), 27 (Zvénigorosky et al., 2020)); 61 samples from other uluses (36 o.d., 25 (Xu et al., 2014)). Data on 157 samples are presented for the first time: 135 from Yakutia and 22 samples from the Yakuts living in the northeastern part of the Khabarovsk Territory. Samples from o.d. were collected after signing a voluntary informed consent form for examination, which was approved by the ethics committee of the Bochkov Research Center for Medical Genetics.
DNA extraction and genotyping. DNA was extracted from 153 samples of venous blood using the
N-P43, N-M231(xM2019/M2118, F4205, P43), Q-M242, O-M175, I-M170, J-M304, R1a-M420, R1b-M343. The “other” category includes the rarest haplogroups: C-M504, E1b1b1-M35.1, G1-M285, G2a-P15, and others. The matrix of Nei’s genetic distances was visualized using multidimensional scaling (MDS) in Statistica version 7.1.
Phylogenetic networks. For a more detailed analysis of the structure of the Yakut haplogroup N-M1982, phylogenetic networks were constructed by median-joining (median networks) (Bandelt, Forster, Röhl, 1999) for the 17-marker haplotypes of the Y-filer set. The weight of each marker was set to 10, with a parameter ε = 0. Median networks were constructed using Network v.10.2.0.0 and visualized in Network Publisher v.2.1.2.5.
The number of tandem repeats at locus DYS389B is defined as the difference in the number of repeats between loci DYS389II and DYS389I. There is a high probability that the N-M1991 lineage corresponds to 16 or more repeats at locus DYS392, while N-M1933 corresponds to 15 (cluster Ht2) (Adamov, 2022). The branches N-M1983 and N-Y25011 within haplogroup N-M1991 are separated by the alleles of locus DYS389B. In the N-M1983 lineage, 18 or more repeats are observed (cluster Ht1). The N-Y25011 branch shows 17 or 16 repeats (cluster Ht3). However, a portion of haplotypes with DYS392 = 16 and DYS389B = 17 belongs to the N-M1983 lineage owing to a back mutation from 18→17 (Ibid.). The frequency ratio of branches N-Y25011 and N-M1983 within cluster Ht3 is currently unknown.
Results and discussion
Gene pools of local Yakut populations based on Y-SNP haplogroup data
Among the 15 Y-SNP haplogroups identified in the Yakuts (Table 1), N-M1982 predominates (85 %); it includes two main branches: N-M1991 (70 %) and N-M1933 (15 %). This dominance influences the multidimensional scaling plots based on Nei’s genetic distance matrices across all haplogroups (Table 2; Fig. 2). For comparability with data from different publications, the “Yakut” haplogroup N-M1982 is included in the parent N3a2 one (also referred to as N-M2019/M2118).
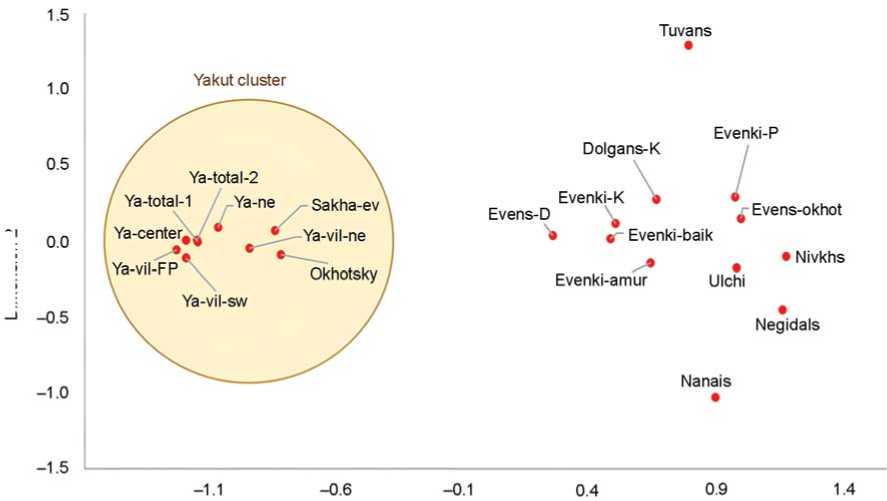
In the gene pools of peoples of the Far East and Siberia, the Yakut cluster is clearly identified (Fig. 2), uniting all Yakuts and Sakha-speaking Evenki (see Fig. 1). This is due to the high frequency of haplogroup N3a2 (68–95 %), which ensured their remoteness from other peoples of Siberia and the Far East (Dolgans, Nanais, Negidals, Nivkhs, Tuvans, Ulchi, Evenki, Evens), where the N3a2 frequency varies from 0 to 21 % (on average 6 %). The values of genetic distances from the averaged Yakut population (see Fig. 2) clearly demonstrate its remoteness from other populations. The closest to the Yakuts are the Evens (Duggan et al., 2013). However, the genetic distance to them ( d = 0.90) is significantly greater than the average genetic distances between Yakut groups ( d = 0.01).
The positioning of populations within the Yakut cluster (see Fig. 2) indicates the predominant contribution of haplogroup N3a2 to its structure. In groups that are more distant from the center, the frequency of this haplogroup is lower than in core populations, but it remains majoritarian: the combined contribution of other haplogroups does not exceed 30 %. The Sakha-speaking Evenki are the only nonYakut group within the Yakut cluster, having joined it owing to the high frequency of N3a2, although their gene pool also includes haplogroups characteristic of the Evenki (C-M48, 15 %) and other populations neighboring the Yakuts: N-P43 (6 %), R1a-M420 (6 %).
When analyzing the positioning of populations in the multidimensional genetic space, all STR haplotypes of the Yakut haplogroup N-M1982 were considered as a whole. The next step will be a more detailed analysis of individual territorial groups of the Yakuts, using data on the distribution of various STR haplotypes of the dominant haplogroup N-M1982 within them.
Genetic history of the Yakuts based on STR haplotypes of dominant haplogroup N-M1982
To construct median networks, 338 samples of haplogroup N-M1982 were used, whose tandem repeats are known in 17 STR loci of the Y-filer panel. The estimation of the time to the most recent common ancestor of main lines N-M1991 and N-M1933 of haplogroup N-M1982 based on STR markers gives an estimate of 1200 ± 480 years. Since these lines are young, STR haplotypes within them generally coincide, except for two STR loci: DYS392 and DYS389B. In the consolidated median network, the STR haplotypes of N-M1982 are distinctly divided into three clusters: Ht1, Ht2, and Ht3 (Fig. 3, a ; Table 2).
The legendary Turkic-speaking ancestor of the Yakuts, Elley (Istoriya Yakutii, 2020: Vol. I, p. 174), is conditionally associated with the Ht1 branch, which is observed in half of the modern Yakuts (see Table 1) (Adamov, 2022). The haplotype of this branch was identified in ancient human sample SS1 from the burial site of Sytygan Sihe, believed to be a descendant of Tygyn Darkhan (Zvénigorosky et al., 2020). The latter, according to legends, was a direct descendant of Elley.
Central Yakutia. According to legends, the second ancestor of the Yakuts, Omogoy Baai, the father-in-law of Elley, was from the Batulintsy tribe (Istoriya Yakutii, 2020: Vol. I, p. 192). A participant of the Second Kamchatka Expedition, G.F. Miller, in 1736–1737, recorded a message from the Yakut shaman Sotoy Trekin about the composition of this tribe: “Baksintsy, Batulintsy, Silangtsy, Igideitsy, Dzhokdzokhontsy, Torasintsy, Orgoty, Baigantaytsy” (cited after (Elert, 2001: 112–113)). The naslegs formed from the names of the Batulintsy clans are currently located in different uluses of Yakutia, including Churapchinsky, Tattinsky, and Tomponsky (see Fig. 1). The Bakhsytsky (village of Tolon) and Alagarsky (village of Chyappara) naslegs border each other to the west of Churapchinsky ulus. In the 17th–18th centuries, the Alagarsky clan was part of the Sylanskaya volost (Dolgikh, 1960: 376). According to legends, the Bakhsytsy and Sylantsy are descendants of Omogoy (Aprosimov, Popov, 2015: 38).
The high frequency of the N-M1933 lineage (approximately 58 %) observed in the Tattinsky and Tomponsky uluses, as well as in the western part of the Churapchinsky ulus, may indicate the contribution of modern descendants of bootulu (Batulintsy) clans to the gene pool of the region. Notably, the
Table 1. Frequency of Y-chromosome haplogroups in regional populations of the Yakuts, %
|
S)n>1BA Лвдсадо рив ‘Д>|5ив>|!ШП1ю -ojnBnj. ‘Л>|5Лв|Л|-оивЛу |
см см II с |
CN N г- О) М Г- LO ID СО СО т- ID оз со от о со от Р 1 р 1 СО 1 1 со 1 от 1 Р 1 1 1 СО СО Ю ^ -^ т- Т- |
|
Э|ОЦМ В SB (вЛцП^ВА) Bl|>|BS 10 □nqnday ащ 11104 S)n>|BA |
со II с |
OTOTOTOTOOCOLDb- -<- ч- -<- Г- СО LO т- ID Tf т- СО т-рротоотююРР cxi ю Р о cxi о Р о о о см о 00 СО СО ID т- т- |
|
(9002 ‘ |B)9jjopua>|Bd) !>|иалз 6ui>|Bads-Bq>|BS >|эиэю |
со со II с |
h- Г- Ю ^ -г- CXJ т- ^ т- Т- т- CMCxiPlDcdPcdlcO 1 СО 1 I ID 1 I I I 1 СО । S N Ю М- с\ т- Т- |
|
(9002 ‘ |B)9jjopua>|Bd) sn|n Л^зиеЛоц^эд ‘ецп^ед шэ^зеэцрои |
см со II с |
ID 1О р ^ ^ V- V- f^ Р • ; : • 1 1 1 1 со 1 1 cd cd 1 1 1 со со 1 00 со |
|
(£102 "IB (a BAOJopaj) ецп^ед ujejsesLipoN |
со со II с |
см cxj cxj o со ip со ip ср id ^ р Р 00 СО ID : СО СО Р О Р cd Р 1 со со 1 1 1 1 Р 1 СО СО СО Т- ^ т- |
|
ецп^ед ujejsesLipoN |
о со II с |
Г^Г'-Г'-CXJIDOOTI'-CO СО CM CXJ т-; СМ Р Р ототосоРотРсоот id см 1 1 cxi 1 Р см 1 1 со Р СР СО СО ОТ т- |
|
> |
(9002 ' |B)ajjopua>iBd) |
СО от II с |
см об о |
CXJ со о |
1 1 |
00 1 1 р 1 |
1 1 |
||||||||||
|
(£102 "IB )Э BAOJopaj) |
от II с |
см от |
CXJ от |
CXJ со |
о |
°? т- |
со |
со со |
от 1 со |
00 1 р 1 1 |
00 р 1 |
||||||
|
uiSBq 1пЛ|!д iua)SBamJON |
со о II |
2 |
к |
со от от |
CXJ со |
со см |
со со |
со |
со |
от о |
со |
00 см т— |
со |
от ^ о о |
S 00 1 CO I V-' |
со 1 |
|
|
с |
|
uiSBq 1пЛ|!д ша^амщпод |
II С |
срсо1Лсо!'*--<-счт- т- q т- -<- т- OTOTi^ibPcxiTtcxil cxi Tt 1 cxi cxi I I I n l l l 00 CO co ^ ^ |
|
(9002 ' |B)ajjopua>|Bd) |
ю от II с |
U’P....T7PP'^T7'^ ^ 6 6 : : : : ^ r^ -^ CM CM CM 1 1 1 1 1 1 1 сб । ОТ от |
|
(£102 "IB )Э BAOJopaj) |
см от II с |
°0 °? °? . . P H ^ P ^ ^ ^ ^ ^ ^ ^ cxi : cxi co ^ cxi 1 P ^ 1 1 IcmI 1 1 co cm 00 CO 1"- ^ |
|
SQSn|D J9L|J0 |
со II с |
CM CXJ о ^ ОТ CXJ CP co ID ОТ CO P ^ ID Ю r< ^ cxi CO T-‘ 1 P 1 cd p P 1 1 p P 1 1 1 1 00 CO r- ID CXJ |
5П|П A^suiqadBjnqg uja)sa/v\ ‘sasnin Л>|suodшol ‘Л^зищв!
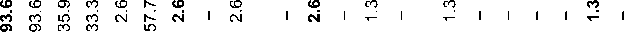
зп|П A>|SUBpiv-)sn
|
co II |
p cd 00 |
ОТ ^ CXI cd cxi cd co co co |
p |
p |
ID |
CO b- О p |
cd |
^ p ID о |
1 cd |
1 1 |
co I I CM I |
|
c |
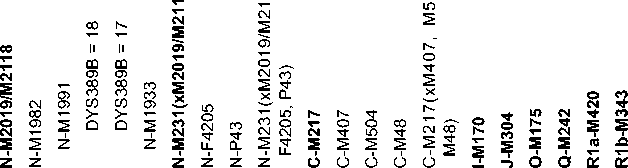
Table 2. Genetic distances of populations from the aggregate Yakut sample
|
Abbreviation |
Population |
Genetic distance * |
Sample size |
|
Ya-total-2 |
Yakuts |
0.000 |
995 |
|
Ya-total-1 |
Yakuts from the Republic of Sakha (Yakutiya) as a whole |
0.000 |
758 |
|
Ya-center |
Yakuts from central Yakutiya |
0.000 |
502 |
|
Ya-vil-FP |
Vilyui Yakuts |
0.001 |
113 |
|
Ya-vil-sw |
Yakuts from southwestern Vilyui basin |
0.002 |
48 |
|
Ya-ne |
Yakuts from Northeastern Yakutia |
0.004 |
187 |
|
Ya-vil-ne |
Yakuts from northeastern Vilyui basin |
0.006 |
109 |
|
Sakha-ev |
Sakha-speaking Evenki (Pakendorf et al., 2006) |
0.018 |
33 |
|
Okhotsky |
Okhotsky Yakuts |
0.023 |
22 |
|
Evens-D |
Evens (Duggan et al., 2013) |
0.893 |
89 |
|
Evenki-baik |
Evenki from Transbaikalia |
1.35 |
50 |
|
Evenki-K |
Western Evenki (Karafet et al., 2018) |
1.37 |
91 |
|
Evenki-amur |
Evenki from the Amur region |
1.64 |
81 |
|
Dolgans-K |
Dolgans, Taimyr (Karafet et al., 2018) |
1.79 |
57 |
|
Ulchi |
Ulchi |
2.59 |
52 |
|
Tuvans |
Tuvans |
2.99 |
450 |
|
Evenki-P |
Evenki, Podkamennaya Tunguska (Pakendorf et al., 2006) |
3.12 |
40 |
|
Nanais |
Nanais |
3.15 |
267 |
|
Negidals |
Negidals |
3.22 |
14 |
|
Evens-okhot |
Evens |
3.25 |
88 |
|
Nivkhs |
Nivkhs |
3.35 |
63 |
*The color scale reflects genetic proximity (green) to the Yakuts or distance (red) from them.
Dimension 2
Dimension 1
Fig. 2 . Position of nine united groups of Yakuts in the genetic space of populations of Siberia and the Far East (multidimensional scaling plot, stress coefficient 0.07, alienation 0.09).
See Table 2 for explanations.
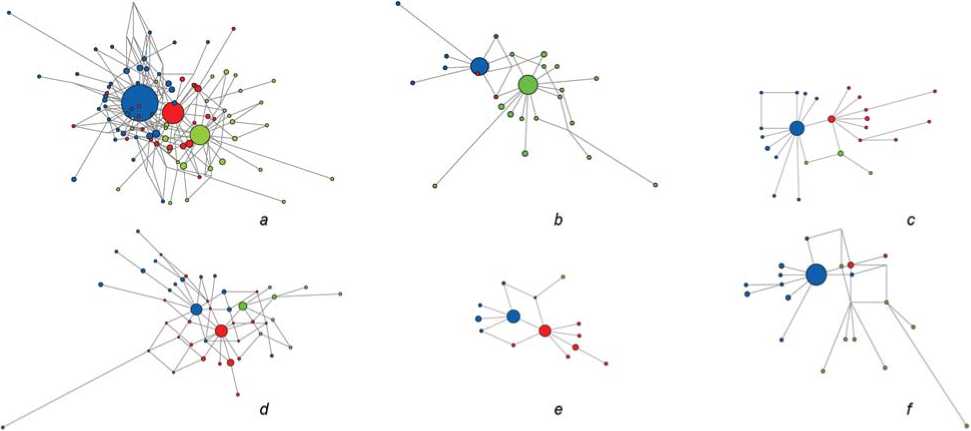
Fig. 3 . Median networks of STR haplotypes of haplogroup N-M1982.
a – Yakuts in general ( n =338); b – Tattinsky, Tomponsky, and western Churapchinsky (Chyappara, Tolon) uluses ( n =73); c – central Yakutia, excluding the above-mentioned uluses ( n =54); d – northeastern Vilyuy basin ( n =80); e – southwestern Vilyuy basin ( n =42); f – northeastern Yakutia ( n =62).
Branch Ht1 (blue) – haplotypes N-M1991, DYS389B = 18; Ht3 (red) – haplotypes N-M1991, DYS389B = 17; branch Ht2 (green) – haplotypes N-M1933.
Table 3. Main (modal) 17-marker STR haplotypes of Yakut samples with haplogroup N-M1982
17-marker haplotypes, constructed from samples from the above-mentioned uluses (Fig. 3, b ). Ht3 haplotypes are rarely found in these areas.
A large group of uluses in central and southern Yakutia—Khangalassky, Megino-Kangalassky, Gorny, Namsky, eastern part of Churapchinsky, Amginsky, Olekminsky, and Aldansky—yielded 61 samples. Clearly, further and more in-depth study is required. At this stage, the distribution of branches of haplogroup N-M1982 among these samples does not significantly differ from the general Yakut distribution (see Table 1).
The ratio of modern haplotypes Ht1 to Ht3 is approximately 2.5 to 1. The dominance of branch Ht1 is also noted in ancient Yakut samples: in six out of seven samples from the Megintsy Yakuts, in eight out of ten samples from the Erkeeni valley in Khangalassky ulus (Keyser et al., 2015; Zvénigorosky et al., 2020), and in seven out of nine from burials in the eastern part of Churapchinsky ulus (Crubézy et al., 2010).
A separate median network for the Yakuts of Ust-Aldansky ulus cannot be constructed owing to the small number ( n =5) of 17-marker haplotypes N-M1982. However, the high frequency (68 %) of haplotypes from cluster Ht1 aligns with the legends of the Dyupsyuntsy and Borogontsy clans about their descent from the legendary ancestor Elley (Aprosimov, Popov, 2015: 28, 31).
The combined median network of Yakut haplotypes from other uluses in central Yakutia (see Fig. 3, c ) shows similarity to the overall Yakut distribution (Fig. 3, a ).
Vilyui Basin . The Vilyui Yakuts are dominated by lineage N-M1991. The frequencies of 17 or 18 repeats at locus DYS389B are approximately equal, indicating an increased frequency specifically for branch N-Y25011. However, in the northeastern Vilyui basin, the dominance of haplogroup N-M1991 is less pronounced (about 60 %) than in the Yakuts living upstream, in the Nyurbinsky and Suntarsky uluses (about 87 %). This distribution is related to the fact that at the time of the Russian pioneers’ arrival, lower reaches of the Vilyui were populated by various groups of “hiking Yakuts” of unknown origin— Kyrgydai, Kirikei, Ontuly, Olesy, etc. (Dolgikh, 1960: 462–469). As a result of mass migration in the 17th–18th centuries of various populations from central Yakutia, some local Tungus clans transitioned to pastoralism and became assimilated.
The ancestor of some naslegs in the northeastern Vilyui basin is believed to be Omogoy Baai, according to local legends (Aprosimov, Popov, 2015: 50–58). The Batulintsy (bootulu) occur among the Vilyui and northern Yakuts (Ushnitsky, 2015). This may explain the presence of samples from haplogroup N-M1933 (14 %) in the dataset. The median network of haplotypes N-M1982 for the northeastern Vilyui basin (Fig. 3, d ) shows a minor proportion of haplotypes Ht2.
The territory of the modern Suntarsky, Nyurbinsky, and Verkhneviluysky uluses, during the 17th– 18th centuries, was settled by Bordontsy, Zharkhantsy, and other migrants from central Yakutia, considering themselves the descendants of Elley (Parnikova, 1971: 47–50, 60–61; Ushnitsky, 2017; Petrov, 2017). In the southwestern Vilyui basin, samples from the “Omogoy branch” N-M1933 are almost absent (see Table 1). A high frequency of lineage N-M1991 is also observed in samples from ancient Yakut burials in the Suntarsky and Nyurbinsky uluses (Keyser et al., 2015; Zvénigorosky et al., 2020), in eight out of nine cases. The median network for the southwestern Vilyui basin (Fig. 3, e ) clearly demonstrates the aforementioned predominance of lineage N-M1991 (with an equal ratio of haplotypes Ht3 and Ht1) and the almost complete absence of cluster Ht2.
Northeastern Yakutia. In this case, samples from different uluses were used (see Fig. 1). The dominance of lineage N-M1991 is accompanied by a relatively high frequency of haplogroup N-M1933 (approximately 9 %). This is probably related to the resettlement of some Bayagantaysky people to the Oymyakon area and along the Moma River. Overall, the frequency of the Yakut haplogroup N-M1982 remains high, around 70 %. All ancient samples (except for one) from burials in the Verkhoyansky (16 samples) and Oymyakonsky (3 samples) uluses belong to cluster Ht1 (Keyser et al., 2015; Zvénigorosky et al., 2020). The increased proportion of this branch is also evident in the median network (Fig. 3, f).
Northeastern Khabarovsk Territory. In the group represented by Ayano-Maysky, Tuguro-Chumikansky, and Okhotsky Yakuts (see Table 1), the predominance of lineage N-M1982 remains at 68 %. The other haplogroups (except for I-P37.2 (9 %))— C-M48 (14 %), N-P43 (4.5 %), and O-M268 (4.5 %)— have Tungus-Manchu or local pre-Yakut origins. Haplotype O-M268 matches that of the Verkhoyansky Evenk across nine STR markers, as reported by B. Pakendorf et al. (2007).
Following the Russian pioneers who founded Fort Okhotsk in 1647, the first Yakuts reached the Sea of Okhotsk in the late 17th to early 18th centuries (Parnikova, 1971: 57), as the rapid extermination of sables, necessary for paying tribute, in central Yakutia forced them to undertake long expeditions for hunting or purchasing sable furs. However, more mass migrations of the Yakuts, both voluntary and forced, occurred later owing to the transportation of goods along the Okhotsk (1731–1852) and Ayan (1852–1867) routes (Ibid.: 91–97). Various goods (fabrics, tea, foodstuffs, manufactured goods, etc.) were transported to ports, and then delivered by sea to Petropavlovsk-Kamchatsky and Alaska. Furs from America (beaver pelts, sea otter furs, etc.) and marine products were unloaded from ships in the ports. Some Yakuts were hired to work for the Russian-American Company. Haplotype N-M1991 of a descendant of one of these Yakuts—a Native American from the Athabaskan tribe (Alaska, USA)— has been published (Davis et al., 2011).
Genetic history of the Yakuts with other haplogroups
Other haplogroups are observed in 15 % of 114 samples of the Yakuts from the Republic of Sakha (see Table 1). According to legends, one of the ancestors of the Sakha, along with Elley and Omogoy, is the elder Uluu Khoro—the leader of the Khorinsky clans. The language of the Khorinsky people was not understood by the Yakuts; therefore, modern researchers believe that the Khorinsky represent a Mongolian component of the Sakha ethnogenesis. Haplogroups C-M407, N-F4205, and C-M504 (“starcluster”), predominant among the Buryats and Barguts, are observed among the Yakuts with an overall frequency of about 3 %. This aligns with the estimated population of Khorinsky people: about 2 % of the total Yakut population in the early 20th century (Ksenofontov, 1992: 161). Notably, one haplotype from the “starcluster” was identified in a resident of the village of Khoro in the Suntarsky ulus. Anthropological materials from Yakut burials of the 17th–19th centuries yielded two samples with haplogroup C-M407 and two with haplogroup N-F4205 (Crubézy et al., 2010; Zvénigorosky et al., 2020).
The overall frequency of haplogroups presumably of Tungus or local pre-Yakut origin (C-M48, N-P43, Q-M242) among the Yakuts does not exceed 5 %. Samples from the Early Iron Age in Yakutia have shown the presence of haplogroup N-P43 (individual from the village of Dyupsya) (Ma et al., 2021) and the absence of the Tat mutation (Pokrovsk man) (Amory et al., 2006).
The paragroup N-M231(xM1991, M1933, F4205, P43) is mainly represented by European branches of N-Tat. Together with other haplogroups atypical for Northeast Asia (I, J, R1a, R1b), it contributes about 6 %. The ancestors of the donors of samples O-P49 (two haplotypes) and O-M1706 (one haplotype) were apparently migrants from the Korean Peninsula.
On the origin of the Yakuts
The observed characteristics of the above territorial groups are explained by the mass migration of the Yakuts from central to more remote areas of Yakutia, and partly beyond its borders, in the 17th–18th centuries. The core of the Yakut people was formed in central Yakutia as a result of a demographic explosion, dated by the polymorphism of haplogroups N-M1991 and N-M1933 to the 14th century AD (Adamov, 2022). Calculations of the founder effect based on autosomal data also point to this date (Tournebize, Chu, Moorjani, 2022). Patrilinearity and patrilocality ensured the stable transmission of the Y-chromosome in the territories inhabited by Yakut clans. Marriages among the Yakuts were determined by exogamous traditions: brides were sought from other uluses or at least from other naslegs. Yakut men easily married Tungus women, but Yakut women rarely married Tungus men (Fedorova, 2008: 183; Fedorova, Khusnutdinova, 2022). These features of Yakut marriage traditions are reflected in the dominance of one relatively young Y-chromosome haplogroup, N-M1982, and a sufficiently high diversity of the mitochondrial DNA pool (Fedorova, 2008: 183–185; Fedorova, Khusnutdinova, 2022).
Conclusions
The combination of results from mass parallel sequencing of the Y-chromosome and the accumulated data on STR haplotypes allowed for the division of the young haplogroup N-M1982, which dominates among the Yakuts (at a level of 85 %), into three branches. The following features have been identified in the main Yakut ethno-local groups:
-
1. Throughout the territory inhabited by the Yakuts, branch N-M1991 with DYS389B = 18 is recorded with high frequency, which is associated with the legendary progenitor Elley. According to tradition, he is considered the male ancestor of most Yakuts: the “Elley line” is found in half of the men.
-
2. Branch N-M1933 is widespread in the areas inhabited by the descendants of the Batulintsy people; in the folk tradition, the latter are related to the second legendary ancestor of the Yakuts, Omogoy, and his tribesmen. In central Yakutia, a high frequency of the “Omogoy line” has been identified in the Tattinsky and Tomponsky uluses, and in the western part of Churapchinsky (Bakhsytsky and Alagarsky naslegs).
-
3. The high frequency of branch N-M1991 with DYS389B = 17 in the Vilyuy basin is apparently associated with the migration of the Bordontsy and Zharkhantsy clans from the middle Lena region in the 17th century, as recorded in Russian written sources. The dominance of haplogroups N-M1983 and N-Y25011 in the northeastern Vilyuy basin is less pronounced than that in the southwest.
-
4. The frequency of Yakut haplogroup N-M1982 decreases to about 70 % in the territorial groups of Yakuts in the northeastern Vilyuy basin, Yakutia, and Khabarovsk Territory.
-
5. The minor Yakut haplogroups have different origins. Haplogroups C-M407, N-F4205, C-M504 (“starcluster”) may reflect a Mongolian component in the Yakut ethnogenesis (frequency 3 %). The frequency of haplogroups of Tungus or local pre-Yakut origin—C-M48, N-P43, Q-M242—is also low (about 5 %) among the Yakuts. Haplogroups that joined the Yakut people at a later time (I, J, R1a, R1b, etc.) have a combined frequency of about 7 %.
Acknowledgment
This study was carried out under the State Assignment of the Ministry of Science and Higher Education of the Russian Federation, for the Bochkov Research Center for Medical Genetics.
