Генетическая характеристика российской популяции немецких классических пони по STR-локусам и генам, влияющим на масть

Автор: Калинкова Л.В., Купцова Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика
Статья в выпуске: 6 т.59, 2024 года.
Бесплатный доступ
Немецкий классический пони - сравнительно молодая порода, официальная регистрация которой состоялась в 2000 году. Порода была создана в Германии с целью разведения лошадей, идеально подходящих для детского конного спорта. Немецкие классические пони умны, добронравны, энергичны, гармонично сложены и исключительно элегантны, выглядят как верховая лошадь в миниатюре. Порода характеризуется наличием нарядной игреневой масти различных оттенков. Игреневая масть у домашних лошадей - результат действия доминантной мутации гена PMEL17 , получившей название Silver, которая оказывает влияние на количество черного пигмента эумеланина, осветляя масть. Как правило, мутация Silver меняет черный цвет волос гривы и хвоста на белый, льняной или дымчатый. С 2010 года немецких классических пони разводят в России. В настоящее время российская популяция немецких классических пони крайне малочисленна: на начало 2023 года насчитывалось 30 маток и 9 жеребцов-производителей. Поголовье племенных пони сосредоточено в одном хозяйстве. Оригинальная и нарядная масть считается одним из важнейших селекционируемых в породе признаков. В настоящей работе впервые дана характеристика генетической структуры российской популяции немецких классических пони с применением ДНК-маркеров. Цель исследования состояла в оценке генетической вариабельности в популяции немецких классических пони с использованием 17 высокополиморфных STR-маркеров, а также в изучении полиморфизма 4 генов, влияющих на пигментацию волоса и имеющих важное селекционное значение. Всего протестировано 32 пони с использованием маркеров генов MC1R , ASIP , PMEL17 , MATP и 17 STR-маркеров: AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10, LEX3, VHL20. Образцы для исследования были отобраны от немецких классических пони в КФХ «Кошелев В.В.» (Тверская обл.). ДНК выделяли из волосяных луковиц с помощью коммерческого набора реагентов ExtraGene™ DNA Prep 200 (ООО «Лаборатория Изоген», Россия). Генотипирование по STR-маркерам проводили с использованием набора реагентов для мультиплексного анализа COrDIS Horse (ООО «ГОРДИЗ», Россия). По результатам тестирования определяли частоту встречаемости аллелей, число наблюдаемых аллелей в каждом локусе (А), эффективное число аллелей (Ae), ожидаемая (He) и наблюдаемая (Ho) гетерозиготность, коэффициент инбридинга (FIS). Результаты исследования продемонстрировали наличие достаточно высокой внутрипородной генетической изменчивости. Число аллелей в каждом STR-локусе варьировало от 3 до 8 при среднем значении 5,53±0,35, показатель наблюдаемой гетерозиготности (Ho), рассчитанный с учетом 16 аутосомных локусов, составил 0,7051±0,0434, коэффициент популяционного инбридинга Fis равнялся -0,0686±0,0196. Установлено, что частота встречаемости доминантного аллеля А гена ASIP и доминантного аллеля Е гена MC1R составила соответственно 0,313 и 0,688, частота мутантного аллеля Silver гена PMEL17 - 0,563. Мутация Cremello гена MATP у животных экспериментальной выборки не обнаружена. С учетом четырех генов, контролирующих пигментацию, у протестированных животных идентифицировано 14 различных вариантов генотипа, что свидетельствует о довольно высокой генетической вариабельности в популяции. Наибольшее распространение имели генотипы, ассоциированные с необычными и нарядными мастями.
Лошади, немецкий классический пони, днк-маркеры, полиморфизм, str-маркеры
Короткий адрес: https://sciup.org/142244124
IDR: 142244124 | УДК: 636.16.082:575 | DOI: 10.15389/agrobiology.2024.6.1108rus
Genetic characterization of the Russian population of German Classic ponies by STR loci and genes affecting coat colour
The German classic pony is a relatively new breed officially registered in 2000. The German classic pony was created in Germany with the aim of breeding horses that are ideal for children. The ponies are smart, good-tempered and energetic, elegant and handsome, looking like a riding mini-horse. The breed is characterized by presence of attractive silver dapple coat colour of various shades. The silver dapple coat colour in horses is the result of the dominant Silver mutation of the PMEL17 gene that affects the black pigment eumelanin. Typically, the Silver gene changes the black colour of hairs of mane and tail to white, flaxen or grey. German classic ponies have been bred in Russia since 2010. At present, the Russian population of German classic ponies is extremely small. At the beginning of 2023, there were 30 purebred brood mares and 9 stallions. The breeding stock is concentrated on one farm. The unique silver coat colour of the ponies is considered one of the most important traits of selection in the breed. The aim of the study was to evaluate the genetic variability using 17 STR markers and to investigate polymorphism of the 4 genes associated with the coat colours in German classic ponies of the Russian population. A total number of 32 animals were genotyped by using markers of genes MC1R , ASIP , PMEL17 , MATP and 17 STR markers: AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10, LEX3, VHL20. Hair samples were collected from the German classic ponies of the farm Koshelev V.V. in 2019. DNA was isolated from hair follicles using the commercial reagent kit ExtraGene™ DNA Prep 200 (Laboratory Isogen, Russia). Genotyping for STR markers was performed using the reagent kit for multiplex analysis COrDIS Horse (GORDIZ, Russia). Based on the testing results, the following parameters were calculated: allele frequencies, the number of observed alleles in each locus (A), the effective number of alleles (Ae), expected (He) and observed (Ho) heterozygosity, and the inbreeding coefficient (FIS). The results revealed a relatively high level of genetic variability in the most of studied STR loci. The number of alleles identified across all evaluated STR loci was 94. The number of alleles per locus varied from 3 to 8 with a mean value of 5.53±0.35. The level of the observed heterozygosity (Ho) for 16 autosomal loci was 0.7051±0.0434, the FIS coefficient was -0.0686±0.0196. It was found that the frequencies of the dominant A allele of the ASIP gene and the dominant E allele of the MC1R gene were 0.313 and 0.688, respectively. The frequency of the mutant Silver allele of the PMEL17 gene was 0,563. The mutant Cremello allele of the MATP gene was not found in the group of genotyped ponies. Given four genes that control pigmentation, 14 different genotype variants were identified in the tested animals, indicating a high level of genetic variability in the population. The most common genotypes in the studied population were associated with the most unusual and attractive coat colors.
Текст научной статьи Генетическая характеристика российской популяции немецких классических пони по STR-локусам и генам, влияющим на масть
Немецкий классический пони — сравнительно молодая порода, официальная регистрация которой состоялась в 2000 году. Порода создавалась в Германии на конном заводе семьи Grober с использованием шетлендских пони европейской селекции и более элегантных по типу американских классических пони. Родоначальником новой породы стал импортированный в Германию в 1965 году караковый жеребец по кличке Jiggs 1958 года рождения, имевший титул чемпиона США 1961 года. Немецкие классические пони умны, добронравны, энергичны, гармонично сложены и исключительно элегантны, имея рост в холке до 112 см, вы- глядят как верховая лошадь в миниатюре, идеально подходящая для детского конного спорта (1).
Один из ярких отличительных признаков породы — широкое распространение нарядной игреневой масти разнообразных оттенков. Игреневая масть у домашней лошади Equus caballus характеризуется специфическим ослаблением интенсивности пигментации покровных и защитных волос. А.С. Красников в 1957 году описывал две основных вариации игреневой масти у лошадей: темно-игреневую, характеризующуюся туловищем шоколадного цвета, часто в яблоках, с белым или дымчатым хвостом и гривой, и светло-игреневую, отличающуюся окраской туловища близкой к светло-коричневой (2, c. 246).
Игреневая масть считается редкой и не встречается в большинстве существующих конских пород, генетический механизм ее наследования изучался с 1950-х годов. В 1953 году американские исследователи W.E. Castle и F.H. Smith дали подробное описание оригинальных мастей silver dapple (серебристая в яблоко) и dappled chestnut (рыжая в яблоко), достаточно часто встречавшихся у шетлендских пони, разводимых в США. Они предположили, что эти фенотипы связаны с неким доминантным геном Silver (серебро) (3). В 1886 году Американским клубом шетлендских пони была зарегистрирована кобылка по кличке Trot, рожденная в США от родителей, привезенных из Великобритании, и имевшая уникальную масть, схожую с палевым окрасом олененка, при этом волосы гривы и хвоста были очень светлыми, почти белыми. Внук кобылы Trot — жеребец по кличке Chestnut, родившийся в 1897 году, стал очень популярным в США производителем. Принято считать, что Chestnut — прародитель всех современных американских пони, обладающих различными вариантами игреневой масти (4).
В 2006 году E. Brunberg c соавт. (5) и в 2007 году M. Reissman c со-авт. (6) в двух независимых исследованиях установили, что игреневая масть у домашней лошади ассоциирована с мутантным вариантом гена PMEL17 , локализованного на 6-й хромосоме. Действие мутантного аллеля, названного Silver, выражается в осветлении покровных и защитных волос за счет уменьшения количества черного пигмента эумеланина, при этом влияние на количество желтого пигмента феомеланина практически отсутствует (7). Эффект ослабления пигментации под действием мутантного аллеля наиболее заметен в длинных волосах гривы и хвоста (8). Согласно D.P. Sponen-berg с соавт. (9), масти носителей мутации демонстрируют обширную фенотипическую изменчивость от ярко выраженного до минимального воздействия доминантного аллеля Silver. По свидетельству ряда исследователей, идентификация мастей, определяемых геном Silver, зачастую бывает затруднена, особенно у молодых животных, при этом масть некоторых лошадей может быть описана как темно-бурая, рыжая с осветленной гривой и хвостом, соловая различных оттенков и т.д. (5, 10).
Вариабильность оттенков масти, наблюдаемая у носителей гена Silver, базируется главным образом на полиморфизме генов MC1R и ASIP (11). Ген MC1R, кодирующий меланокортиновый рецептор 1-го типа, имеет два основных аллеля: доминантный аллель дикого типа контролирует выработку черного пигмента эумеланина, мутантный рецессивный аллель в гомозиготном состоянии подавляет синтез эумеланина и обеспечивает синтез красно-желтого пигмента феомеланина, обусловливая тем самым рыжую масть (9, 10). В настоящее время известно, что рецессивная мутация у лошадей рыжей масти представляет собой однонуклеотидную замену C > T в гене MC1R (11). Ген ASIP, кодирующий агути-сигнальный белок, влияет на синтез и характер распределения эумеланина. Доминантный аллель дикого типа контролирует гнедую масть, значительно ограничивая синтез эумеланина. У гнедых лошадей черный пигмент может накапливается только в определенных местах: в волосах ног, гривы и хвоста. Рецессивная мутация гена ASIP, представляющая собой делецию размеро 11 п.н., не оказывает влияния на синтез и распределение эумеланина и в гомозиготном состоянии определяет вороную масть (12).
В ряде заводских и локальных пород лошадей, в том числе у шет-лендских пони, встречается мутантный вариант гена MATP , получивший название Cremello, который ослабляет пигментацию кожи и волос, действуя по типу неполного доминирования, осветляя в основном красножелтый пигмент феомеланин и определяя буланую, соловую и изабелло-вую масти (8, 9). Мутация Cremello представляет собой однонуклеотидную замену G > A в экзоне 2 гена MATP (13). В отличие от гена Cremello, мутантный ген Silver осветляет волосы за счет уменьшения количества черного пигмента эумеланина, практически не оказывая влияния на количество красно-желтого пигмента феомеланина (14).
С 2010 года немецких классических пони стали разводить в России (1). В настоящее время российская популяция немецких классических пони крайне малочисленна: на начало 2023 года насчитывалось 30 чистопородных кобыл, используемых в разведении, и 9 жеребцов-производителей. Производящий состав породы сосредоточен в одном племенном хозяйстве — КФХ «Кошелев В.В.» (Тверская обл.). Оригинальная и нарядная масть племенных пони считается одним из важнейших селекционируемых в породе признаков.
Для успешной селекционной работы в популяциях животных с ограниченным генофондом особую важность приобретает проблема поддержания генетического разнообразия. Для оценки параметров генетического разнообразия в популяциях широко используются высокополиморфные STR-маркеры (short tandem repeat) (15, 16). В начале 1990-х годов в научной литературе появились первые описания STR-маркеров у домашней лошади (17, 18). К настоящему времени анализ ДНК с использованием STR-маркеров признан одним из наиболее эффективных инструментов для проведения генетических экспертиз по индивидуальной идентификации и подтверждению достоверности происхождения племенных и спортивных лошадей, в практике коннозаводства тестирование применяется повсеместно в качестве обязательной процедуры при регистрации племенного поголовья (14, 19, 20).
В большинстве современных генетических лабораторий, как правило, используются стандартизированные коммерческие наборы реагентов для генотипирования лошадей (StockMarks® Kit for Horses, «Applied Bio-systems», США; COrDIS Horse, ООО «ГОРДИЗ», Россия), включающие 17 STR-маркеров (21-23).
В настоящей работе впервые дана характеристика генетической структуры российской популяции немецких классических пони с применением ДНК-маркеров.
Цель исследования состояла в оценке генетической вариабельности в популяции немецких классических пони с использованием 17 высокополиморфных STR-маркеров, а также в изучении полиморфизма генов MC1R , ASIP , PMEL17 и MATP , влияющих на пигментацию волос и имеющих важное селекционное значение.
Методика . Образцы волос были отобраны от 32 немецких классических пони, используемых в разведении в Российской Федерации (КФХ 1110
«Кошелев В.В.», Тверская обл. 2019 год).
ДНК выделяли из волосяных луковиц с помощью коммерческого набора реагентов ExtraGene™ DNA Prep 200 (ООО «Лаборатория Изоген», Россия). Генотипирование по 17 STR-маркерам (AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10, LEX3, VHL20) проводили с использованием набора реагентов для мультиплексного анализа COrDIS Horse (ООО «ГОРДИЗ», Россия) согласно рекомендациям производителя.
При генотипировании биологических образцов по маркерам генов, детерминирующих масть, применяли коммерческие наборы реагентов GenPak® PCR Core (ООО «Лаборатория Изоген», Россия) в соответствии с рекомендациями производителя.
Мутацию гена ASIP (делеция 11 п.н.) детектировали согласно S. Rieder с соавт. (12) с использованием опубликованных последовательностей праймеров (F: 5´-CTTTTGTCTCTCTTTGAAGCATTG-3´, R: 5´-GAGAAG-TCCAAGGCCTACCTTG-3´) с последующим разделением полученных фрагментов электрофорезом в 3 % агарозном геле.
Генотипирование по SNP-маркерам C > T в гене MC1R , G > A в экзоне 2 гена MATP и C > T в экзоне 11 гена PMEL17 проводили методом PCR-RFLP (polymerase chain reaction—restriction fragment length polymorphism) с использованием праймеров, дизайн которых был выполнен с помощью программ Primer-BLAST (27): F: 5´-CTGCACTCACCCATGTAC-TA-3´, R: 5´-GCAAGATTGCCATCTCCAGC-3´ для MC1R ; F: 5´-CTGGGCC-ATAACCATCACCA-3´, R: 5´-TCCCTGTCTGGGGAGCTAAA-3´ для MATP ; F: 5´-CTGCCCTTGTCCATTGCTTAC-3´, R: 5´-GCAGCCAGTGGGTTCT-ACC-3´ для PMEL17 . Детекцию однонуклеотидных полиморфизмов в выделенных фрагментах ДНК проводили с использованием эндонуклеаз рестрикции соответственно TaqI , TasI и HhaI с последующим разделением полученных фрагментов в 3 % агарозном геле.
Для обозначения аллелей в исследованных локусах была использована буквенная номенклатура согласно M. Reissmann (8): ген MC1R — аллели E (доминантный аллель дикого типа) и e (рецессивный мутантный аллель); ген ASIP — аллели A (доминантный аллель дикого типа) и a (рецессивный мутантный аллель); ген PMEL17 — аллели Z (доминантный мутантный аллель Silver) и z (рецессивный аллель дикого типа); ген MATP — аллели C (аллель дикого типа, неполное доминирование) и cCr (мутантный аллель Cremello).
Статистическую обработку полученных результатов проводили в программе Microsoft Excel. Рассчитывали частоту встречаемости аллелей, число наблюдаемых аллелей в каждом локусе (А), эффективное число аллелей (Ae), ожидаемую (H e ) и наблюдаемую (H o ) гетерозиготность, коэффициент инбридинга (F is ). Вычисляли средние значения ( M ) и стандартные ошибки средних (±SEM).
Ðåçóëüòàòû. Анализ STR-профилей немецких классических пони показал достаточно высокую генетическую изменчивость. В целом у 32 животных в 17 локусах было обнаружено 94 аллеля (табл. 1). Число аллелей в каждом локусе варьировало от 3 до 8 при среднем значении 5,53±0,35, показатель наблюдаемой гетерозиготности (Ho), рассчитанный с учетом 16 аутосомных локусов, составил 0,7051±0,0434. Коэффициент популяционного инбридинга Fis в большинстве исследованных локусов имел отрицательное значение при среднем (отрицательном) значении -0,0686±0,0196, что свидетельствует о преобладании в популяции гетерозиготных генотипов. По показателям степени полиморфности и гетерозиготности российская популяция немецких классических пони несколько превосходила исследованные ранее L.H.P. Van de Goor с соавт. (23) популяции шетлендских пони, разводимых в странах Западной Европы. Следует отметить, что в связи с малочисленностью популяции немецких классических пони в России может быть полезным регулярное проведение мониторинга с использованием STR-локусов, рекомендованных в качестве эффективного инструмента при оценке параметров генетического разнообразия (24-26).
1. Показатели генетического разнообразия в популяции немецких классических пони по STR-локусам ( n = 32; КФХ «Кошелев В.В.», Тверская обл., 2019 год)
|
STR-локусы |
Показатель |
||||
|
A 1 |
A e |
H e |
H o |
Fis |
|
|
AHT4 |
8 |
5,13 |
0,8052 |
0,8750 |
-0,0867 |
|
AHT5 |
6 |
4,29 |
0,7671 |
0,7500 |
0,0223 |
|
ASB17 |
8 |
4,25 |
0,7646 |
0,8125 |
-0,0626 |
|
ASB2 |
6 |
4,03 |
0,7520 |
0,7500 |
0,0026 |
|
ASB23 |
4 |
3,72 |
0,7310 |
0,7188 |
0,0167 |
|
CA425 |
4 |
2,05 |
0,5122 |
0,5625 |
-0,0982 |
|
HMS1 |
4 |
1,77 |
0,4341 |
0,4688 |
-0,0799 |
|
HMS2 |
4 |
3,27 |
0,6938 |
0,7500 |
-0,0809 |
|
HMS3 |
5 |
2,14 |
0,5327 |
0,6250 |
-0,1732 |
|
HMS6 |
6 |
4,10 |
0,7559 |
0,8750 |
-0,1576 |
|
HMS7 |
7 |
5,15 |
0,8057 |
0,8750 |
-0,0861 |
|
HTG10 |
7 |
4,39 |
0,7725 |
0,7813 |
-0,0114 |
|
HTG4 |
6 |
2,66 |
0,6245 |
0,7500 |
-0,2009 |
|
HTG6 |
5 |
2,48 |
0,5962 |
0,5625 |
0,0565 |
|
HTG7 |
3 |
1,34 |
0,2515 |
0,2500 |
0,0058 |
|
LEX3 |
6 |
3,57 |
|||
|
VHL20 |
5 |
4,02 |
0,7515 |
0,8750 |
-0,1644 |
|
M ±SEM |
5,53±0,35 |
3,43±0,28 |
0,6594±0,0393 |
0,7051±0,0434 |
-0,0686±0,0196 |
Примечание. A — число наблюдаемых аллелей, A e — число эффективных аллелей, H e — ожидаемая гетерозиготность, H o — наблюдаемая гетерозиготность, F IS — коэффициент инбридинга. LEX3 локализован на Х-хромосоме, мужские особи гемизиготны, поэтому показатели гетерозиготности и FIS для этого локуса не вычисляли.
По результатам генотипирования по генам MC1R , ASIP и PMEL17 в каждом исследованном локусе было выделено по 3 генотипа: гомозиготный по доминантному аллелю, гомозиготный по рецессивному аллелю и гетерозиготный (рис. 1). По гену MATP все протестированные нами пони имели гомозиготный по дикому аллелю генотип C / C , мутация Cremello у животных экспериментальной выборки не обнаружена. Подавляющее число пони (31 гол.) были носителями доминантного аллеля E гена MC1R , контролирующего синтез меланоцитами черного пигмента эумеланина. Только один пони имел гомозиготный по рецессивному аллелю генотип e / e , ассоциированный с рыжей мастью.
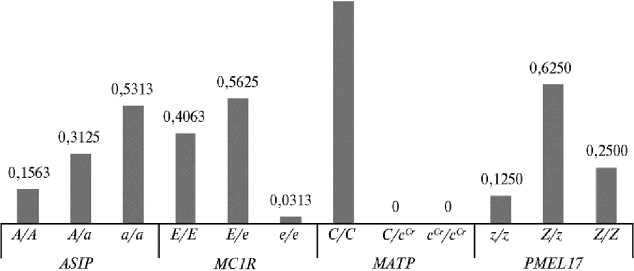
Рис. 1. Частота встречаемости различных генотипов в генах ASIP , MC1R , MATP и PMEL17 у немецких классических пони (КФХ «Кошелев В.В.», Тверская обл., 2019 год) .
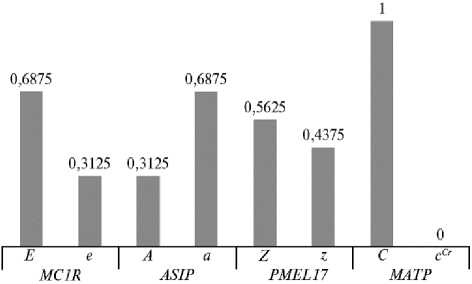
Рис. 2. Частота встречаемости аллельных вариантов генов MC1R , ASIP , PMEL17 и MATP в популяции немецких классических пони (КФХ «Кошелев В.В.», Тверская обл., 2019 год) .
Частота встречаемости доминантного аллеля E в популяции составила 0,688, частота встречаемости доминантного аллеля A гена ASIP — 0,313 (рис. 2). Носителями доминантного аллеля A были 15 животных, у 17 пони обнаружили гомозиготный по рецессивному аллелю генотип a/a. Большинство протестированных животных (28 пони) оказались носителями доминантного мутантного аллеля Z (Silver) гена PMEL17, в том числе 8 животных имели гомозиготный генотип Z/Z.
Частота встречаемости аллеля Z в производящем составе немецких классических пони, разводимых в России, составила 0,563 (рис. 2). Оче- видно, что оригинальные, осветленные по типу Silver масти служат одним из наиболее значимых селекционируемых в породе признаков.
2. Варианты генотипов немецких классических пони по полиморфизмам генов ASIP , MC1R , MATP и PMEL17 (КФХ «Кошелев В.В.», Тверская обл., 2019 год)
|
Генотипы |
Число животных |
Частота встречаемости в популяции |
|
a/a-E/E-C/C-Z/Z |
1 |
0,0313 |
|
a/a-E/E-C/C-Z/z |
7 |
0,2188 |
|
a/a-E/e-C/C-Z/Z |
3 |
0,0938 |
|
a/a-E/e-C/C-Z/z |
4 |
0,1250 |
|
a/a-E/e-C/C-z/z |
1 |
0,0313 |
|
A/A-E/E-C/C-Z/Z |
1 |
0,0313 |
|
A/A-E/e-C/C-Z/z |
2 |
0,0625 |
|
A/A-E/e-C/C-z/z |
2 |
0,0625 |
|
A/a-E/E-C/C-Z/Z |
1 |
0,0313 |
|
A/a-E/E-C/C-Z/z |
3 |
0,0938 |
|
A/a-E/e-C/C-Z/Z |
1 |
0,0313 |
|
A/a-E/e-C/C-Z/z |
4 |
0,1250 |
|
A/a-E/e-C/C-z/z |
1 |
0,0313 |
|
a/a-e/e-C/C-Z/Z |
1 |
0,0313 |
В целом по четырем генам, детерминирующим пигментацию, у 32 животных экспериментальной выборки было идентифицировано 14 различных вариантов генотипа (табл. 2).
Масти исследованных нами пони характеризовались широкой фенотипической изменчивостью и разлной степенью влияния мутации Silver на изменение интенсивности пигментации покровных и защитных волос (рис. 3). При этом масти гомозиготных носителей доминантного аллеля Z (Silver) выделялись большей степенью осветления.
У пони с наиболее ти- пичными для исследованной популяции генотипами a/a-E/E-C/C-Z/z и a/a-E/e-C/C-Z/z цвет покровных волос туловища варьировал от серокоричневого (оттенок сепии) до интенсивно-шоколадного, наблюдалось наличие яблок, волосы гривы и хвоста были белыми или дымчатыми. У пони с генотипами a/a-E/E-C/C-Z/Z и a/a-E/e-C/C-Z/Z туловище серокоричневого цвета с ярко-выраженными яблоками, волосы гривы и хвоста белые. Животные с генотипами A/A-E/E-C/C-Z/Z и A/a-E/E-C/C-Z/Z характеризовались светло-рыжим цветом туловища, светлыми волосами гривы и хвоста, значительным осветлением волос в нижней части ног, в паху и в области подпруги; участки темной пигментации на ногах имели серокоричневый оттенок. Цвет волосяного покрова туловища у пони с генотипами A/A-E/e-C/C-Z/z, A/a-E/E-C/C-Z/z, A/a-E/e-C/C-Z/z варьировал от рыжего до темно-рыжего, волосы гривы и хвоста были осветлены в различной степени (рыжеватые, дымчатые или белые). Пони — носитель ге- нотипа A/a-E/e-C/C-Z/Z отличался серо-коричневым (оттенок сепии) цветом туловища, светлыми волосами гривы и хвоста, незначительным осветлением волос в нижней части ног. Пони с генотипом a/a-e/e-C/C-Z/Z имел рыжую масть с осветлениями в нижней части ног, в паху и в области подпруги. Животные с генотипами A/a-E/e-C/C-z/z и A/A-E/e-C/C-z/z имели гнедую масть. Масть пони с генотипом a/a-E/e-C/C-z/z выглядела как вороная «в загаре».

Рис. 3. Вариации мастей у немецких классических пони в зависимости от генотипа: a — A/A-E / E-Ñ / Ñ-Z / Z , б — a / a-E / e-Ñ / Ñ-Z / Z , в — A / a-E / E-Ñ / Ñ-Z / z , г — a / a-E / E-Ñ / Ñ-Z / Z , д — a/a-e / e-Ñ / Ñ-Z / Z , е — a / a-E / e-Ñ / Ñ-Z / z (КФХ «Кошелев В.В.», Тверская область, 2019 год).
Согласно D.P. Sponenberg с соавт. (9), группа мастей, основанных на ослаблении пигментации по типу Silver, получила широкое распространение в таких породах, как лошадь скалистых гор (Rocky Mountain Horse) и исладская лошадь. Однако вследствие плейотропного действия мутации Silver у ее носителей встречаются так называемые множественные врожденные аномалии глаз (multiple congenital ocular abnormalities), в наиболее тяжелых случаях у животных может наблюдаться снижение зрительных функций (9). Среди владельцев исландских лошадей существует мнение, что носители аллеля Silver более нервные и пугливые (4, 28). Исследовав на ограниченном числе животных различия в поведенческих реакциях исландских лошадей в зависимости от масти, E. Brunberg с соавт. (28) пришли к выводу, что носители мутации Silver более осторожны в новых ситуациях, при этом не отличаются от других лошадей большей реактивностью при испуге. Предрасположенность носителей аллеля Silver к наруше- ниям зрения должна учитываться заводчиками лошадей в селекционной работе (4).
Таким образом, исследование российской популяции немецких классических пони с использованием 17 STR-маркеров продемонстрировало наличие достаточно высокой внутрипородной генетической изменчивости: при среднем значении числа аллелей на локус, равном 5,53±0,35, показатель наблюдаемой гетерозиготности с учетом 16 аутосомных локусов составил 0,7051±0,0434. Отрицательное значение коэффициента популяционного инбридинга F is ( - 0,0686±0,0196) свидетельствует о преобладании в популяции гетерозиготных генотипов. Изучение полиморфизма генов MC1R , ASIP, PMEL17 и MATP, контролирующих пигментацию кожи и волос, показало, что частота встречаемости доминантного аллеля A гена ASIP и доминантного аллеля E гена MC1R составила соответственно 0,3125 и 0,6875. Частота встречаемости доминантного аллеля Z гена PMEL17 составила 0,5625. Мутация Cremello гена MATP у животных экспериментальной выборки не обнаружена. В исследованной популяции преобладали аллельные варианты генов MC1R и ASIP , детерминирующих синтез темного пигмента эумеланина, а также мутантного варианта гена PMEL17 , уменьшающего количество этого пигмента в волосах и осветляющего масть. У протестированных нами пони чаще встречались генотипы a / a-E/ E-C / C-Z / z , a / a-E/ e-C/ C-Z/ z , A / a-E / e-C/ C-Z / z , a / a-E/ e-C/ C-Z / Z , A / a-E/ E-C / C-Z / z , ассоциированные с наиболее необычными и нарядными мастями.
