Генетическая обусловленность полевой всхожести семян у межвидовых гибридов Triticum aestivum x T. durum Desf

Автор: Жарков Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетические основы селекции
Статья в выпуске: 3 т.47, 2012 года.
Бесплатный доступ
В полевых и лабораторных опытах изучали жизнеспособность семян у сортов яровой мягкой и яровой твердой пшеницы, у гибридных комбинаций F 1 и F 2, полученных от скрещивания Triticum aestivum x T. durum, а также у пентаплоидов F 1 и ВС 1. Выявлена достаточно сложная система генетического контроля анализируемого признака, происхождение которой может быть связано с особенностями эволюции геномов А и В твердой и мягкой пшеницы. На основании сравнительного анализа всхожести семян у гибридов первого и второго поколения выявлены два базисных гена, доминантные аллели которых принадлежат T. durum, рецессивные — T. aestivum. Сделано предположение о наличии специфических генов, связанных с сортовыми особенностями родительских форм, использованных при получении первого поколения гибридов.
Пшеница, сорт, линия, межвидовые гибриды, полевая всхожесть семян, корреляция, ген, геном
Короткий адрес: https://sciup.org/142133313
IDR: 142133313 | УДК: 633.11:631.522/.524:631.527.5:631.524.821
Genetic determination of field germination ability in seeds of interspecific hybrids of Triticum aestivum x T. durum Desf
Obtained from crossing Triticum aestivum x T. durum, and also in pentaploids F 1 and ВС 1. The sufficiently complicated system of genetic control of analyzed determinant was revealed, the origin of which can be relates with features of evolution of A and B genomes of durum and soft wheat. On the ground of comparative analysis of seed germinating ability in hybrids of the first and second generation the author revealed two basic genes, the dominant alleles of which belong to T. durum, but the recessive genes — to T. aestivum. The assumption was made about availability of specific genes, related with varietal features of initial parents.
Текст научной статьи Генетическая обусловленность полевой всхожести семян у межвидовых гибридов Triticum aestivum x T. durum Desf
При отдаленной гибридизации формирование невсхожих зерен может быть вызвано эмбриональной летальностью (8). Отмечается абортиро-вание гибридных зародышей. В результате в зрелых зерновках содержатся мертвые зародыши (9).
Дальнейшие исследования летальных генов связаны в основном с определением их географического распространения, степени влияния на элементы продуктивности колоса и растения, хромосомной локализации, с поиском новых генетических систем (10-12). Так, скрещивая мягкую пшеницу с рожью, Н.Д. Тихенко с соавт. (13) обнаружили наличие у ржи специфического гена Eml (Embryo lethality), блокирующего развитие гибридного зародыша у амфигаплоидов. Высказано предположение о комплементарном взаимодействии генов пшеницы и ржи, вызывающем отсутствие дифференциации зародыша и, как следствие, потерю всхожести семян у пшенично-ржаных гибридов F 1 . Позднее комплементарный ген Eml-
A1 у мягкой пшеницы был определен на хромосоме 6А (14).
Целью настоящей работы стало изучение особенностей генетического контроля полевой всхожести семян у межвидовых гибридов Triticum aestivum х T . durum .
Методика . Исследования проводили в лабораторных и полевых условиях (г. Омск). Объектами служили яровая мягкая пшеница (сорт Омская 37 и линия Г 695/01), яровая твердая пшеница (сорта Ангел, Омская янтарная, Омский кристалл) и шесть гибридных комбинаций F 1 и F2, полученных от скрещивания яровой мягкой (?) и яровой твердой (^) пшеницы. Кроме того, изучали пентаплоидные гибриды F 1 и ВС 1 по 13 гибридным комбинациям. Для получения пентаплоидных форм ВС | использовали сорта Памяти Азиева, ДИАС 2, Эритроспермум 59 яровой мягкой пшеницы и сорта Ангел, Саратовская золотистая яровой твердой пшеницы. Пентаплоидные гибриды ВС 1 получали посредством скрещивания выделенных из популяции F2 28-хромосомных растений с мягкой пшеницей и 42-хромосомных — с твердой пшеницей. Полевые опыты закладывали в 2008 году в оптимальные сроки по пару. Площадь питания растений составляла 10х20 см. В процессе появления всходов по каждой гибридной комбинации подсчитывали число проростков.
Полученные данные обрабатывали математически методами корреляционного и дисперсионного анализов (15, 16).
Результаты . Весенне-летний период в год проведения опытов характеризовался контрастными погодными условиями. В мае I и II декады оказались засушливыми (среднесуточная температура воздуха превышала норму соответственно на 0,1 и 5,0 ° С, количество выпавших осадков составило 38 и 25 % от среднего многолетнего), III декада — прохладной и дождливой. В сложившихся агроклиматических условиях наблюдалась относительно низкая полевая всхожесть семян как у родителей, так и у их гибридов (табл. 1). По сортам она варьировала от 51,4 до 63,3 %, по гибридам F 1 — в пределах 35,0-65,0 %, F2 — 24,8-47,1 %.
1. Полевая всхожесть семян у сортов и межвидовых гибридов Triticum aestivum х T . durum (Сибирский НИИ сельского хозяйства, г. Омск, 2008 год)
|
Сорт, линия, гибридная комбинация |
Потомство |
Высеяно зерен, шт. |
Число всходов, шт. |
Всхожесть семян, % |
|
Омская 37 |
Р |
60 |
34 |
56,7 |
|
Линия Г 695/01 |
Р |
70 |
37 |
52,9 |
|
Ангел |
Р |
60 |
38 |
63,3 |
|
Омская янтарная |
Р |
70 |
37 |
52,9 |
|
Омский кристалл |
Р |
70 |
36 |
51,4 |
|
Омская 37 х Ангел |
F 1 |
60 |
35 |
58,3 |
|
F 2 |
210 |
81 |
38,6 |
|
|
Омская 37 х Омская |
F 1 |
60 |
39 |
65,0 |
|
янтарная |
F 2 |
210 |
99 |
47,1 |
|
Омская 37 х Омский |
F 1 |
60 |
38 |
63,3 |
|
кристалл |
F 2 |
210 |
89 |
42,4 |
|
Линия Г 695/01 х Ангел |
F 1 |
60 |
21 |
35,0 |
|
F 2 |
210 |
54 |
25,7 |
|
|
Линия Г 695/01 х Ом- |
F 1 |
60 |
24 |
40,0 |
|
ская янтарная |
F 2 |
210 |
52 |
24,8 |
|
Линия Г 695/01 х Ом- |
F 1 |
60 |
28 |
46,7 |
|
ский кристалл |
F 2 |
210 |
65 |
30,9 |
|
Суммарное значение |
F 1 |
360 |
185 |
51,4 |
|
F 2 |
1260 |
440 |
34,9 |
|
|
П р и м е ч а н и е. Р — родительская форма. |
||||
Согласно результатам трехфакторного дисперсионного анализа, основной вклад (53 %) в общую генотипическую изменчивость анализи- руемого признака у межвидовых гибридов вносила мягкая пшеница. Несколько меньшее влияние (42 %) оказывали особенности гетерозиготного состояния гибридов F1 и F2. Твердая пшеница и взаимодействие факторов обусловливали соответственно 2 и 3 % от общей совокупности генотипической изменчивости.
Гибриды F1, полученные на основе одного сорта (линии) мягкой пшеницы, имели близкие показатели. В то же время гибридные комбинации от разных материнских сортов существенно различались между собой. Известно, что межвидовые гибриды первого поколения (T. aestivum х T. durum ) гомогенны по хромосомно-генетическому составу. При этом в гетерозиготном состоянии они имеют хромосомные наборы геномов А и В (1 n мягкой пшеницы + 1 n твердой пшеницы), а в гемизиготном — геном D, который полностью принадлежит мягкой пшенице. Поскольку твердая пшеница не оказывала существенного влияния на характер варьирования полевой всхожести семян по комбинациям, можно предположить, что этот признак у межвидовых гибридов определялся в основном хромосомами генома D. Однако результаты анализа популяции гибридов F2 свидетель ствуют об обратном.
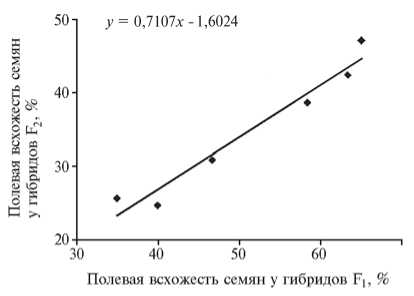
Корреляционная зависимость между полевой всхожестью семян у межвидовых гибридов Triticum aestivum х T. durum в F i и F2 (Сибирский НИИ сельского хозяйства, г. Омск, 2008 год) .
Межвидовые гибриды второго поколения по полевой всхожести семян значительно уступали таковым первого. Разница между средними показателями у двух поколений составила 16,5 % (Р < 0,001). Существенное снижение всхожести гибридных зерен F2 может быть обусловлено вы-щеплением генотипов с пониженной жизнеспособностью семян.
По результатам корреляционного анализа между гибридами F 1 и F2 проявилась достаточно высокая прямо пропорциональная зависимость (рис.).
Коэффициент корреляции в этом случае составил 0,976 (Р < 0,001).
В отличие от пентаплоидных форм F1 (2n = 35) у растений второго поколения геном D мог быть представлен 0-14 хромосомами, а общее их число варьировало в пределах 28-42. Зависимости между характером рас- щепления популяции по плоидности генотипов и полевой всхожестью семян не обнаружили. Сохранение в F2 тех же закономерностей изменчивости анализируемого признака, что и в F1, вряд ли возможно только за счет сортовой специфичности хромосом генома D. Очевидно, решающую роль в детерминации полевой всхожести играют гены, расположенные в хромосомах геномов А и В. Это предположение подтверждается результатами анализа полевой всхожести семян у пентаплоидных форм F1 и BC1 (табл. 3).
3. Полевая всхожесть семян у межвидовых гибридов Triticum aestvum х T durum F i и BC 1 (Сибирский НИИ сельского хозяйства, г. Омск, 2008 год)
|
Гибридная комбинация сортов |
Пентаплоиды гибридов |
d |
|||
|
F 1 |
BC 1 |
||||
|
высеяно зерен, шт. |
полевая всхожесть, % |
высеяно зерен, шт. |
полевая всхожесть, % |
||
|
П р я м ы е с к р е щ и в |
а н и я ( $ 2 n = |
42; 5 2 n = 28) |
|||
|
(Памяти Азиева х Саратовская |
|||||
|
золотистая) х Саратовская золо- |
|||||
|
тистая |
60 |
51,7 |
50 |
76,0 |
24,3* |
|
(Памяти Азиева х Ангел) х Ангел |
60 |
53,3 |
30 |
53,3 |
0 |
|
(ДИАС 2 х Ангел) х Ангел |
60 |
43,3 |
60 |
56,7 |
13,4 |
|
(Ангел х ДИАС 2) х Ангел |
30 |
76,7 |
33,4** |
||
|
(Эритроспермум 59 х Ангел) х Ангел |
60 |
68,3 |
50 |
70,0 |
1,7 |
|
О б р а т н ы е с к р е щ и в а н и я ( $ 2 n |
= 28; 5 2 n = 42) |
||||
|
(Саратовская золотистая х Памяти |
|||||
|
Азиева) х Памяти Азиева |
60 |
53,3 |
50 |
0 |
- 53,3** |
|
(Памяти Азиева х Саратовская |
|||||
|
золотистая) х Памяти Азиева |
60 |
0 |
- 53,3** |
||
|
(Саратовская золотистая х ДИ- |
|||||
|
АС 2) х ДИАС 2 |
60 |
36,7 |
10 |
0 |
- 36,7* |
|
(ДИАС 2 х Саратовская золоти- |
|||||
|
стая) х ДИАС 2 |
40 |
0 |
- 36,7** |
||
|
Ангел х ДИАС 2 |
60 |
33,3 |
|||
|
(ДИАС 2 х Ангел) х ДИАС 2 |
60 |
5,0 |
- 28,3** |
||
|
Саратовская золотистая х Эри- |
|||||
|
троспермум 59 |
60 |
25,0 |
|||
|
(Эритроспермум 59 х Саратовская |
|||||
|
золотистая) х Эритроспермум 59 |
60 |
21,7 |
- 3,3 |
||
|
(Ангел х Эритроспермум 59) х Эри- |
|||||
|
троспермум 59 |
60 |
18,3 |
60 |
8,3 |
- 10,0 |
|
(Эритроспермум 59 х Ангел) х Эри- |
|||||
|
троспермум 59 |
60 |
20,0 |
- 1,7 |
||
П р и м е ч а н и е. При получении семян F i использовались только два первых компонента скрещивания указанной гибридной комбинации. При сравнительном анализе гибридов F i и BC i между собой учитывалось направление скрещивания. Пропуск означает, что гибридную комбинацию не анализировали.
* и ** Соответственно Р < 0,05 и Р < 0,001.
При получении пентаплоидов ВС 1 посредством скрещивания выделенных из F2 42-хромосомных форм с рекуррентным родителем твердой пшеницы (2 n = 28) сохранялась достаточно высокая полевая всхожесть семян межвидовых гибридов. В отдельных случаях она значительно превосходила этот показатель у гибридов первого поколения (см. табл. 3). Однако ситуация менялась, если в качестве материнского растения использовали 28-хромосомные, а отцовского — 42-хромосомные формы (обратные скрещивания). По ряду гибридных комбинаций высеянные семена оказались невсхожими. При этом результат не зависел от первоначального направления скрещивания родительских форм, использованных при получении первого поколения гибридов (см. табл. 3).
Известно, что гибридизация эффективнее, если в качестве материнского используется вид с меньшим числом хромосом, а всхожесть выше, когда меньшее число хромосом имеет отцовский компонент скрещивания (17). Считается, что это обусловлено особенностями двойного оплодотворения (17). В наших опытах полная потеря всхожести семян при бек-кроссировании в обратных скрещиваниях и ее сохранение — в прямых могут быть связаны с особенностями эволюции геномов А и В у мягкой и твердой пшеницы. Так, если у T. aestivum формирование их генетических комплексов проходило в присутствии третьего генома — D, то у T. durum процесс осуществлялся без его участия. Благодаря этому стал возможен перенос генома D от мягкой пшеницы в твердую (18), однако при обратных скрещиваниях, когда происходило замещение хромосом геномов А и В твердой пшеницы на аналогичные геномы мягкой, они (в силу особенностей эволюционного развития геномов А и В у T. aestivum) при гемизи-готном состоянии хромосом генома D не могли обеспечить формирование жизнеспособных генотипов. Тем не менее, у мягкой пшеницы существуют анеуплоидные линии, генотип которых ограничивается хромосомным набором двойных моносомиков. Создать же на собственном генофоне T. aestivum пентаплоидные формы, а тем более выделить в чистом виде первые два генома оказывается весьма проблематично.
У растений в F 1 и F 2 не было обнаружено признаков хлороза, некроза или гибридной карликовости, что исключает возможность влияния этих летальных генов на жизнеспособность семян у межвидовых гибридов. Очевидно, характер проявления анализируемого признака определялся другими генами и связан со спецификой межгеномного взаимодействия хромосом.
Как отмечалось ранее, межвидовые гибриды F2 по полевой всхожести семян значительно уступали F 1. Разница между ними может быть принята как параметр, определяющий степень утраты жизнеспособности семян из-за особенностей гетерозиготного состояния генотипа во втором расщепляющемся поколении. В результате проведенных расчетов было установлено, что его значение варьировало по гибридным комбинациям от 9,3 до 20,9 % (табл. 4).
4. Сравнительный анализ полевой всхожести семян у межвидовых гибридов Tri-ticum aestivum х T . durum (Сибирский НИИ сельского хозяйства, г. Омск, 2008 год)
|
Гибридная комбинация сортов и линий |
Полевая всхожесть семян, % |
d |
|
|
F 1 I |
F 2 |
||
|
Омская 37 х Ангел |
58,3 |
38,6 |
- 19,7* |
|
Омская 37 х Омская янтарная |
65,0 |
47,1 |
- 17,9* |
|
Омская 37 х Омский кристалл |
63,3 |
42,4 |
- 20,9** |
|
Линия Г 695/01 х Ангел |
35,0 |
25,7 |
- 9,3 |
|
Линия Г 695/01 х Омская янтарная |
40,0 |
24,8 |
- 15,2* |
|
Линия Г 695/01 х Омский кристалл |
46,7 |
30,9 |
- 15,1* |
|
* и ** Соответственно Р < 0,05 и P < 0,01. |
|||
По найденному проценту различий легко определить соотношение величин в натуральном выражении. Число зерен ( n ), утративших жизнеспособность в результате перехода генотипа в следующую стадию гетерозиготного состояния (F2), можно найти по формуле: n = (N х ПР)/100, где N — число высеянных зерен F2; ПР — разница между полевой всхожестью семян в F 1 и F2, %. Число зерен, в которых летальное действие этой генетической системы не проявилось, составит разницу между числом высеянных зерен и найденным значением n . Сложившиеся соотношения между двумя группами анализируемых генотипов представлены в таблице 5.
Анализ соответствия фактического расщепления теоретически ожидаемому показал, что почти во всех гибридных комбинациях оно происходило по двум генам, причем в одном случае (линия Г 695/01 х сорт Ангел) составило 15:1, во всех остальных — 13:3. Основываясь на характере проявления анализируемого признака у пентаплоидных форм первого поколения, можно утверждать, что жизнеспособность семян определялась до- минантными аллелями генотипа твердой пшеницы.
5. Характер расщепления в популяции межвидовых гибридов F 2 Triticum aestivum х T . durum по генам полевой всхожести семян (Сибирский НИИ сельского хозяйства, г. Омск, 2008 год)
|
Гибридная комбинация сортов и линий |
Фактическое соотношение двух групп генотипов |
Значение х 2 при теоретически ожидаемом расщеплении (данные по критическим вариантам) |
|
|
3:1 | 15:1 |
13:3 |
||
|
Омская 37 х Ангел |
168,6:41,4 |
3,13 |
0,12 |
|
Омская 37 х Омская янтарная |
172,4:37,6 |
0,10 |
|
|
Омская 37 х Омский кристалл |
166,1:43,9 |
1,96 |
0,63 |
|
Линия Г 695/01 х Ангел |
196,8:13,2 |
0,00 |
|
|
Линия Г 695/01 х Омская янтарная |
178,1:31,9 |
1,76 |
|
|
Линия Г 695/01 х Омский кристалл |
176,8:33,2 |
1,20 |
|
|
П р и м е ч а н и е. х 2 0,05 = 3,84. Указаны только значения х 2, не превышающие теоретически ожидае- |
|||
|
мые и, следовательно, соответствующие одному из трех видов расщепления. |
|||
Появление у 1 / 16 части генотипов F2 невсхожих зерен могло быть вызвано гомозиготным состоянием аллелей двух рецессивных генов мягкой пшеницы. Полученное расщепление 13:3 свидетельствует о неполном доминировании одного из двух генов T . durum при гомозиготном состоянии рецессивных аллелей второго гена.
Исходя из одинаковой величины отклонения показателей всхожести семян у гибридов F2 от F 1 , можно сделать вывод, что определяющие ее гены относятся к базисным (19), обусловливающим специфику видовой принадлежности растений.
В наших опытах полевая всхожесть семян существенно варьировала в тех гибридных комбинациях (см. табл. 1), где основной вклад в общую генотипическую изменчивость вносила мягкая пшеница. Из этого следует, что T . aestivum , кроме базисных генов, имеет и специфические (19), определяющие сортовые особенности детерминации анализируемого признака. Это предположение подтверждается результатами анализа пентаплоидных форм ВС 1 . В гибридных комбинациях обратных скрещиваний, где в качестве рекуррентного родителя использовался сорт яровой мягкой пшеницы Эритроспермум 59, полевая всхожесть семян в той или иной степени сохранялась (см. табл. 3). Наличие высокой корреляционной зависимости между полевой всхожестью семян у гибридов F 1 и F2 (см. рис.) указывает на то, что расщепление по специфическим генам во втором поколении практически отсутствовало либо проявлялось незначительно.
Таким образом, полевая всхожесть семян у межвидовых гибридов Triticum aestivum х T . durum контролируется достаточно сложной генетической системой и связана с особенностями межгеномных взаимодействий хромосом у мягкой и твердой пшеницы. При этом в состав такой системы могут входить как базисные, так специфические гены. По нашим данным, доминантные аллели, определяющие жизнеспособность семян у межвидовых гибридов, принадлежат твердой пшенице, рецессивные — мягкой.
Л И Т Е Р А Т У Р А
-
1. Д е к а п р и л е в и ч Л.Л. О получении нежизнеспособных и полужизнеспособных комбинаций при скрещивании пшениц. Тр. Всесоюзного съезда по генетике, селекции, семеноводству и племенному животноводству, 1930: 221.
-
2. Д о р о ф е е в В.Ф., М е р е ж к о А.Ф. Проблемы гибридного некроза у пшеницы. Генетика, 1969, V(4): 161-167.
-
3. H e r m s e n J.G.Th. Hybrid dwarfness in wheat. Euphytica, 1967, 16(2): 134-162.
-
4. T s u n e w a k i K. A new tipe of hibrid chlorosis found in tetraploid wheats. Jap. J. Genet., 1968, 43(4): 279-288.
-
5. H а с к и д а ш в и л и П.П. Межвидовая гибридизация пшеницы. М., 1984.
-
6. П у х а л ь с к и й В.А., М а р т ы н о в С.П., Д о б р о т в о р с к а я Т.В. Гены гибридного некроза пшениц. Теория вопроса и каталог носителей летальных генов. М., 2002.
-
7. W o r l a n d A.J. Further studies on hybrid dwarfing in wheat. Indused Variability Plant Breeding Int. Symp. Sect. Mutat. and Polyploidy Eur. Assoc. Res. Plant Breed. EUCARPIA, Wageningen, Aug. 31-Sept. 4, 1981. Wageningen, 1982: 106-109.
-
8. Б а т ы г и н а Т.Б. Хлебное зерно. Атлас. Л., 1987.
-
9. R a i n a S.K. Crossability and in vitro development of hybrid embryos of Triticum durum x Secale
cereale . Indian J. Genet., 1984, 44(3): 429-437.
-
10. П у х а л ь с к и й В.А. Новые данные в изучении генов гибридного некроза в роде Triticum L. Тез. докл. 2-го Съезда Вавиловского общества генетиков и селекционеров. СПб, 2000, т. 1: 122-123.
-
11. S h a r m a R.K., C h o w d h u r y S., S e t h i A.P. Identification of hybrid necrosis genes among some bread wheat accessions. Wheat Inf. Serv., 2001, 92: 25-26.
-
12. T s u n e w a k i K. Aneuploid analysis of albino genes in tetraploid wheats. Wheat Inf. Serv., 2003, 96: 39.
-
13. Т и х е н к о Н.Д., Ц в е т к о в а Н.В., В о й л о к о в А.В. Генетический контроль эмбриональной летальности при скрещивании мягкой пшеницы с рожью. Генетика, 2005, 41(8): 1075-1083.
-
14. T i k h e n k o N., T s v e t k o v a N., D o b r o v o l s k a y a O., Z a y n a l i N e z-h a d K., R o d e r M.S., B o r n e r A. Embryo lethality in wheat x rye hybrids — mode of inheritance and the identification of a complementary gene in wheat. Euphytica, 2010, 176(2): 191-198.
-
15. Д о с п е х о в Б.А. Методика полевого опыта. М., 1979.
-
16. П л о х и н с к и й Н.А. Биометрия. М., 1970.
-
17. Б у д а ш к и н а Е.Б. Цитогенетика межвидовых гибридов пшеницы. В сб.: Цитогенетика пшеницы и ее гибридов. М., 1971: 196-221.
-
18. Ж и р о в Е.Г., Т е р н о в с к а я Т.К. Перенос генома D от мягкой пшеницы в твердую. Докл. АН СССР, 1987, 296: 1252-1254.
-
19. Р е й т е р Б.Г., Е в д о к и м о в М.Г. Моносомный анализ устойчивости к бурой ржавчине мягкой яровой пшеницы. Генетика, 1979, XV(6): 1067-1075.
ГНУ Сибирский НИИ сельского хозяйства Россельхозакадемии,
