Генетически разные органогенные постройки в изотопных величинах D13С и D18О

Автор: Антошкина А.И., Шмелева Л.А., Леонова Л.В., Добрецова И.Г., Смолева И.В.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 11 (323), 2021 года.
Бесплатный доступ
Исследован изотопный состав углерода и кислорода 165 образцов карбонатных пород генетически отличающихся органогенных построек: верхнеордовикского рифа Большая Косью, верхнедевонского микробиального холма Шаръю, скелетного холма Кожым, верхнемиоценового биогермного комплекса Казантип и глубоководных органогенно-карбонатных построек севера Срединно-Атлантического хребта. Для рифа и микробиального холма характерны значения d13СPDB в пределах, близких к значениям нормально-осадочных морских карбонатов (-0.33-3.13 ‰) и (0.8-3.0 ‰), но с существенными вариациями d18ОSMOW -(22.24-30.0 ‰) и (20.4-26.3 ‰) соответственно. Наиболее изотопно-тяжелым составом углерода (5.1-7.3 ‰) в сочетании с варьирующими значениями d18ОSMOW (22.4-30.0 ‰) характеризуются известняки и брахиоподы скелетного холма. Колебания значений d13СPDB и d18ОSMOW в карбонатных породах биогермного комплекса Казантип являются самыми выразительными: (-2.76-7.17 ‰) и (24.20-33.1 ‰) соответственно. Глубоководные органогенные постройки со дна приосевой зоны с активным вулканизмом, в отличие от других, показали устойчивые значения d13СPDB (-0.98-0.83 ‰) в пределах области нормально-осадочных морских карбонатов, тогда как для кислорода значения являются изотопно-тяжелыми (32.27-39.75 ‰). В итоге специфика формирования объектов исследования, установленная литолого-палеоэкологическими и химико-физическими методами, нашла свое обоснование в изотопных величинах, палеосолености и палеотемпературах, обусловленных палеогеографической и климатической ситуацией.
Риф, микробиальный и скелетный холмы, биогермный комплекс, глубоководные органогенно-карбонатные постройки, изотопный состав углерода и кислорода
Короткий адрес: https://sciup.org/149139289
IDR: 149139289 | УДК: 551.8.07+550.42 | DOI: 10.19110/geov.2021.11.3
Genetically different organogenic buildups in isotope values D13C and D18O
The isotopic composition of carbon and oxygen of 165 samples of carbonate rocks of genetically different organogenic structures: the Upper Ordovician Bol'shaya Kos'yu reef, the Upper Devonian Shar'yu microbial mound, the Kozhym skeletal mound, the Upper Miocene Kazantip bioherm complex and deep-water organogenic-carbonate structures of the north of the Mid-Atlantic Ridge was studied. The reef and microbial mound are characterized by d13CPDB values within the limits close to normal-sedimentary marine carbonates (-0.33-3.13 ‰) and (0.8-3.0 ‰), but with significant variations in d18OSMOW - (22.24-30.0 ‰) and (20.4-26.3 ‰), respectively. The most isotopically-heavy carbon composition (5.1-7.3 ‰) in combination with varying d18OSMOW values (22.4-30.0 ‰) is characteristic of limestones and brachiopods of the skeletal mound. Fluctuations of d13CPDB and d18OSMOW values in carbonate rocks of the Kazantip bioherm complex are the most expressive (-2.76-7.17 ‰) and (24.20-33.1 ‰), respectively. Deep-water organogenic structures from the bottom of the axial zone with active volcanism, in contrast to others, showed stable d13CPDB values (-0.98-0.83 ‰) within the area of normal-sedimentary marine carbonates, whereas for oxygen the values are istopic-heavy (32.27-39.75 ‰). As a result, the specificity of the development of the objects under study, established by lithological-paleoecological and chemical-physical methods, found its substantiation in isotopic values, paleosalinity and paleotemperatures caused by paleogeographic and climatic conditions.
Текст научной статьи Генетически разные органогенные постройки в изотопных величинах D13С и D18О
Ископаемые и современные органогенные простые и сложные постройки, в особенности рифы, являются важными индикаторами для реконструкций палеогеографической и палеотектонической обстановок. Древние организмы, как и современные, в процессе жизнедеятельности были приспособлены к среде обитания, и экологическая обстановка находила отражение в формах и адаптивных функциях ор-ганизмов-каркасостроителей, которые выражались в их облике, взаимном пространственном расположении, отношении к субстрату, в наборах жизненных форм, биотическом доминировании [21, 23, 29, 32, 40, 44, 47].
Кислород и углерод входят в число наиболее распространенных элементов Земли, они тесно связаны с процессами, происходящими в биосфере, гидросфере и литосфере, и являются одними из наиболее важных элементов для карбонатных пород, характеризуясь значительными природными вариациями в изотопном составе. Поэтому данные δ 13С и δ 18О в карбонатах разного генезиса являются полезным инструментом для реконструкции физико-химических условий осадконакопления. В последние десятилетия появилось много публикаций по изотопно-углеродным и изотопнокислородным характеристикам карбонатов биогенной и абиогенной природы: океанических, морских, лагунных, озерных, континентальных, пещерных и гидротермальных [5, 8, 13, 15—17, 20, 26, 33, 38, 48, 51]. Исследован изотопный состав углерода и кислорода в фоссилиях с целью определения палеотемператур, солености, палеоклимата [7, 9, 11, 39, 42, 48, 51].
Цель данной статьи состоит в демонстрации возможности использования изотопных характеристик генетически разных органогенных построек для обоснования их генезиса на примере изученных объектов.
Объекты и методы исследования
Объектами исследований были морфологически и генетически разнообразные органогенные сооружения: риф Большая Косью (Северный Урал), микробиальный холм Шаръю (гряда Чернышева), скелетный холм Кожым (Приполярный Урал), мшанковые био-гермные постройки Казантип (Крым), трубчатые и ветвистые органогенно-карбонатные постройки (Северная Атлантика). На изотопный состав углерода и кислорода проанализированы 165 проб пород, слагающих вышеупомянутые органогенные карбонатные постройки. При этом использовались методы аналитической сканирующей электронной микроскопии (СЭМ (JSM 6400 Jeol с энергодисперсионным и волновым спектрометрами, В. Н. Филиппов); электронного парамагнитного резонанса (В. П. Лютоев); изотопной масс-спектрометрии (DeltaVAvantage с аналитическим комплексом ThermoFisher). Значения изотопных коэффициентов δ 13С и δ 18О даны относительно стандартов соответственно PDB и SMOW, калиброванного по международному стандарту NBS 19 (TS-limestone). Ошибка определений как δ 13С, так и δ 18О не превышала ± 0.1 ‰ (1 σ ). Для отбора материала для исследований с наименее измененных участков пород применялись тонкие стоматологические сверла. Аналитическая рабо- 28
та была осуществлена в ЦКП «Геонаука» ИГ ФИЦ Коми НЦ УрО РАН.
Геологическое и палеогеографическое положение исследуемых объектов
Геологическая позиция, палеогеографическое положение и генезис исследуемых органогенных карбонатных образований существенно отличались по характеру морских обстановок и климатическим условиям в среде обитания. Верхнеордовикский (средний катий) риф Большая Косью мощностью 150 м в естественных выходах имеет протяженность до 2 км вдоль р. Большая Косью в бассейне р. Илыч на западном склоне Северного Урала. Палеогеографически этот участок располагался на окраине Тимано-Североуральского эвапорито-карбонатного мелководного шельфа в пределах рифовой отмели с подвижной гидродинамикой [1]. Исследованный франско-фаменский микробиальный холм мощностью 300 м прослеживается на расстоянии 2 км в естественных выходах на р. Шаръю в северной части гряды Чернышева, являющейся западным ограничением Предуральского краевого прогиба на северо-востоке Европейской платформы. Подобные микробиальные холмы в пределах расчлененного карбонатного шельфа обрамляли склоны изолированных карбонатных платформ, окруженных прогибами с относительно глубоководным некомпенсированным карбонатно-кремнисто-глинистым осадконакоплением [34]. Гжельско-сакмарский скелетный холм мощностью до 330 м наблюдается в виде крупных скальных выходов на протяжении 2 км вдоль р. Кожым на западном склоне Приполярного Урала. Такого рода образования развивались в указанное время на склонах впадин, возникших в результате деградации карбонатного шельфа во время формирования Предуральского краевого прогиба [35]. Верхнемиоценовый мшанко-вый биогермный комплекс , слагающий толщу мощностью до 60 м, расположен на территории Казан-типского природного заповедника, где породы образуют прерывистые в рельефе скальные гряды вокруг округлой котловины (по-тюркски — «казан»), давшей название мысу на севере Керченского полуострова. Верхнемиоценовые постройки обрамляют брахианти-клиналь, представляющую собой криптодиапировую складку с ядром нагнетания из сильно перемятых, раздробленных и перетертых глин майкопской серии [12]. Мшанковые биогермы формировались на склонах диапировой складки как пригидротермальные оазисы в мелководном бассейне с газофлюидным высачива-нием из грифонов древнего грязевого вулкана [36]. Голоцен-верхнеплейстоценовые глубоководные ор-ганогенно-карбонатные постройки размером, редко превышающим 15—20 см в длину, формировались на Срединном Атлантическом хребте (САХ) на глубинах около 3 км, во внеосевой зоне рифтовой долины с активным вулканизмом. Субстратом для них служили молодые пикробазальты и туффиты, породы габ-бро-перидотитового комплекса, участками перекрытые рыхлыми кокколито-фораминиферовыми илами. Трубчатые и ветвистые формы построек образовались путем нарастания на поверхности океанических вулканитов или непосредственно в среде относительно рыхлых пелагических осадков [2, 6].
Краткая литологическая характеристика объектов исследования
Риф Большая Косью (рис. 1, а) сложен комплексом биогермных (рис. 1, с, d) и биокластовых известняков, грубообломочных известняковых конглобрек-чий (рис. 2, b) и разнозернистых известняковых песчаников. Наибольшее участие в этом принимают известняки: биогермные, биокластовые, литокластовые и микросгустковые. Основными внешними признаками пород являются светло-серый цвет, массивность сложения, обилие фоссилий разнообразной бентосной фауны и флоры — цианобактерий, красных и зеленых водорослей, сфинктозойных губок (рис. 1, c), рецепта-кулитов, проблематических гидроидов, кораллов-ругоз, табулятоморфных кораллов, строматопороидей, гелеолитид (рис. 1, e), раковинная фауна и незначительное количество терригенного материала. Важным диагностическим признаком массивных рифовых пород является также их текстурная неоднородность, обусловленная пятнистым распределением крупных каркасостроящих организмов, межкаркасным заполнением иловым, зернистым и обломочным материалом (рис. 1, f). Кроме того, в этих породах широко развит инкрустационный раннедиагенетический цемент, обрастающий частицы обломочного и биокластового материала и заполняющий промежутки между каркасными организмами. Формирование такой ассоциации определялось гидродинамическими обстановками рифовой отмели, палеоландшафтом рифового плато и условиями формирования биогенного каркаса рифа Большая Косью [28,30].
Микробиальный холм Шаръю (рис. 3, а) представлен в целом светлыми массивными известняками, имеющими линзовидно-наслоенное строение (рис. 2, b). Разрез сложен ассоциациями строматолитовых (рис. 2, c, e) и микробиальных биогермных, спонгиа-строматовых, поростроматовых, микрокомковатых и фенестровых (рис. 2, i) известняков. Биогермные известняки включают прослои органогенных известняковых песчаников (рис. 2, d), несортированных поли-биокластовых известняков (рис. 2, h) и более редких их сортированных разностей. Среди известняков встречаются небольшие линзы брахиоподовых ракушняков и маломощные прослои известняковых псевдобрекчий с палеомикрокодиями. Последние характеризуют субаэральные обстановки. Основными каркасобразу-ющими организмами являлись скелетные, нарастающие друг на друга (рис. 2, f, g) и коркооблекающие (рис. 2, c, e) цианобактерии [34].
Скелетный холм Кожым (рис. 3, a) сложен массивно-плитчатыми известняками — биокластовыми, илово-биокластовыми, биогермными (мшанково-ту-бифитесовыми, тубифитесово-мшанковыми, реже па-леоаплизиновыми биоцементолитами) и брахиоподо-выми ракушняками (рис. 3, c, g). Отмечаются отчетливо слоистые пласты микрозернистых известняков с рассеянным тонким скелетным материалом и илово-биокластовых известняков комковатого сложения. Обломочные породы, за исключением палеоапли-зиновых гравелитов и биокластовых разностей, не обнаружены. В зависимости от положения в разрезе они либо линзовидно переслаиваются, либо чередуются. Органические остатки (целые и фрагменты) характеризуются широким таксономическим спектром: бра- хиоподы, мшанки, фораминиферы, палеоаплизины, гастроподы, остракоды, членики и фрагменты стеблей криноидей, иглы морских ежей, ортоцератиды, аммо-ноидеи, хвостовые щитки трилобитов, спикулы известковых губок, ругозы, табуляты (единичные), реже па-леоаплизины (рис. 3, b) цианобактерии, разнообразные известковые водоросли (зеленые, проблематические), микропроблематика и скелетный лом [35].
Мшанковый биогермный комплекс Казантип развит в бортах бухт до береговых уступов (рис. 4, а). Он включает довольно разнообразные типы пород, изученные нами в трех бухтах [36]. В биогермах пространство между колониями мшанок заполнено тонкобио-кластовым и тонкообломочным материалом (рис. 4, c). На поверхности колоний мшанок присутствуют крусти-фикационно-нарастающие с разной цветовой гаммой карбонатные корки толщиной от 5 мм до 3 см (рис. 4, d). Обнаружены биогермы со следами частичного растворения карбонатных построек мшанок и образованными в этих полостях корками гётит-лимонитового состава. На разных уровнях разреза в биогермах встречены желваки, линзы белых и черно-белых опалов. Межбиогермное пространство заполнено главным образом мшанковыми биокластовыми известняками (рис. 4, б), среди которых отмечаются линзовидные тонкозернистые известняки и микро- и мелкообломочные карбонатные брекчии. В ассоциации с ними встречаются скопления мелкой биокластики — целых раковин моллюсков и их фрагментов (рис. 4, e), а также трубки червей-полихет, гастроподовые банки и моллюсково-полихетовые минибиогермы. СЭМ-исследования выявили в составе корок минерализованные биопленки с бактериоморфными структурами (рис. 4, g) и изобилие аутигенных минералов — барита, гипса, галита, стронцианита, целестина, кутногорита, транзитно прослеживающихся не только в подстилающих глинистопесчаных отложениях, но и в минибиогермах и мшанковых известняках.
Глубоководные органогенно-карбонатные постройки Северной Атлантики (рис. 5, a), образовавшиеся в условиях активного вулканизма и постоянных струй газофлюидов, подразделяются на отдельные мор-фотипы [2], среди которых наиболее преобладают ветвистые колонии (рис. 5, b). В телах построек имеются приосевые каналы, которые могут быть заполнены биогенными карбонатными осадками. На поверхности тела часто покрыты корочкой карбонатно-железомарганцевого состава, а в поперечном срезе карбонатных тел отчетливо проявляется концентрически-зо-нальное строение (рис. 5, e). Редко встречаются совсем молодые постройки без корки, но тоже с концентрической зональностью и хорошо сохранившимися реликтами фауны. СЭМ-исследования выявили внутри и на поверхности построек фоссилии кокколитофо-рид, спикул губок, фораминифер, трубчатых червей, а также минерализованные биопленки с бактерио-морфными структурами, включениями нанофоссилий и аутигенных минералов [2] (рис. 5, c, d, f).
Изотопная характеристика исследуемых объектов
Разными авторами вариации величин δ 13С для осадочных морских карбонатов определяются в пре- 29


Рис. 1 . Особенности строения рифа Большая Косью:
a — характер обнаженности пород; b — грубообломочные известняковые конглобрекчии; c — фоссилии красных водорослей Kozhymella dembovskia Sh.(K) с заполнением межкаркасных полостей остатками цианобактерий Renalcis sp. (R) и сфинктозой-ными губками Corymbospongia sp. (C); d — фоссилии предположительно зеленых сифониковых водорослей; e — колонии Heliolitoidea; f — грубопятнистая текстура, характерная для рифовой толщи с неравномерным распределением илового, зернистого и обломочного материала в межкаркасных полостях
Fig. 1. Features of the structure of the Bol’shaya Kosyu reef:
a — fragments of outcrops; b — coarse limestone conglobreccias; c — red algae Kozhymella dembovskia Sh. (K) with filling of interframe cavities with cyanobacteria Renalcis sp. (R) and sphinctozoa sponges Corymbospongia sp. (C); d — possible green siphonic algae; e — a colony of Heliolitoidea; f — coarse-spotted texture, characteristic of the reef thick, represented by the distribution of silt, granular and detrital material in interframe cavities
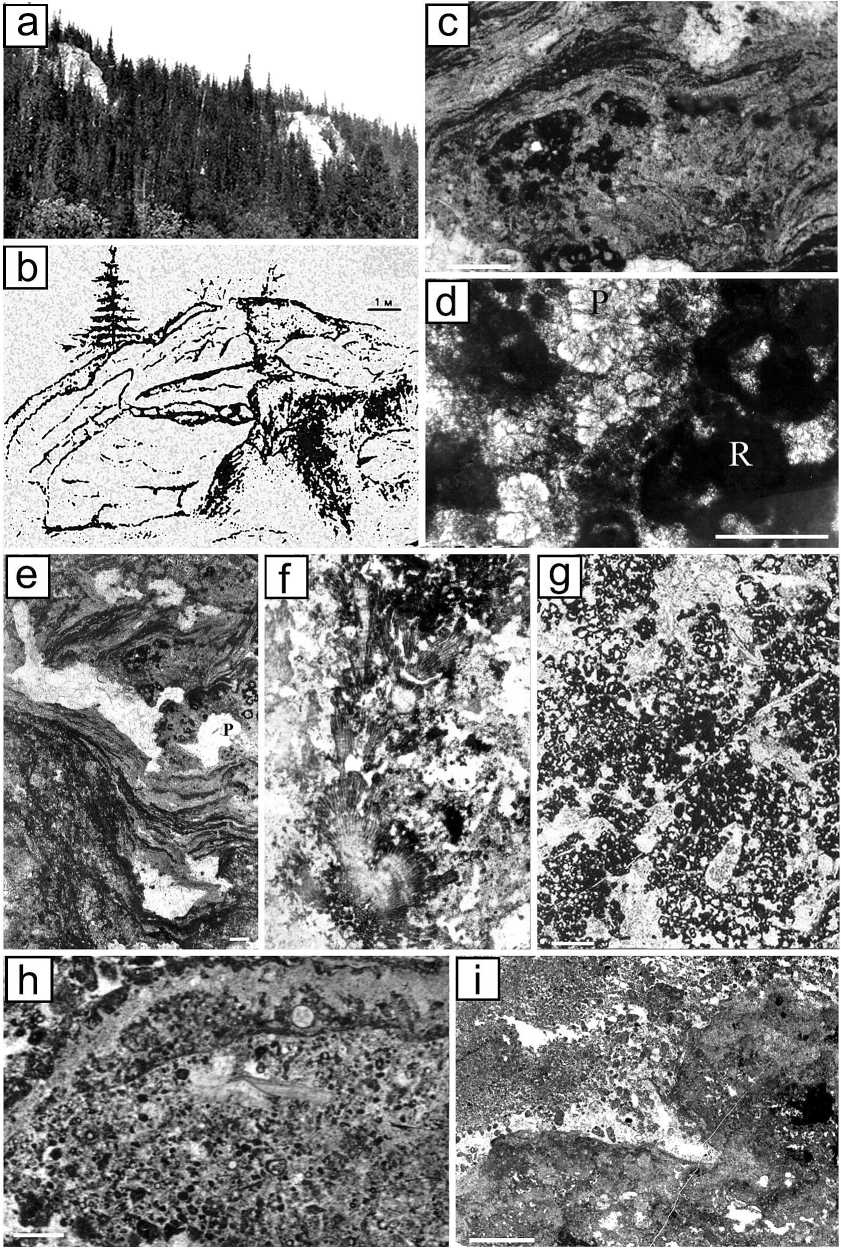
Рис. 2. Характерные типы пород и представители биогермных ассоциаций среднефранско-фаменского микробиального холма Шаръю:
a — фрагмент выходов на р. Шаръю, обн. 86; b — биогермный строматолитовый известняк, образованный чередованием цианобактерий Sphaerocodium и Girvanella вокруг скоплений Renalcis и Shuguria , шлиф, р. Шаръю; с — полевая зарисовка характера строения микробиального холма; d—i — шлифы: d — розетки Paleomicrocodium в биогермном микробиальном известняке, R — Renalcis ; e — строматолитовый биогермный известняк с Rothpletzella-Girvanella-Renalcis-ассоциацией и крупная фенестровая пористость, которая может образовываться в субаэральных средах при выщелачивании вадозной(?) пресной водой; f — ветвистые колонии эпифи-тоноподобных Shariuphyton в биогермном микробиальном известняке; g — микробиальный биогермный известняк, сложенный Renalcis-Shuguria-Chabakovia- ассоциацией; h — известняк несортированно-биокластовый с обилием микробиальных комков; i — псевдобрекчия с цементом вадозного алеврита. Масштабная линейка — 1 мм
Fig. 2. Typical types of rocks and representatives of bioherm associations of the Sharyu Middle Frasnian-Famennian microbial mound: a — a fragment of outcrop on the Sharyu River, outcrop 86; b—f — thin sections: b — bioherm stromatolite limestone formed by alternating cyanobacteria Sphaerocodium and Girvanella around Renalcis and Shuguria clusters, Sharyu Riv.; c — field sketch of the nature of the structure of the microbial mound; d — Palaeomicrocodium rosettes in a bioherm microbial limestone, R — Renalcis ; e — stromatolite boundstone constructed by Rothpletzella-Girvanella-Renalcis association and larger fenestral porosity that could form in subaerial environments by vadose(?) freshwater leaching; f — branched colonies of epiphyton-like Shariuphyton in a bioherm microbial limestone; g — a microbial bioherm limestone composed by the Renalcis-Shuguria-Chabakovia association; h — unsorted bioclastic limestone with an abundance of microbial lumps;. i — vadose silt in pseudibreccia. Scale bar — 1 mm

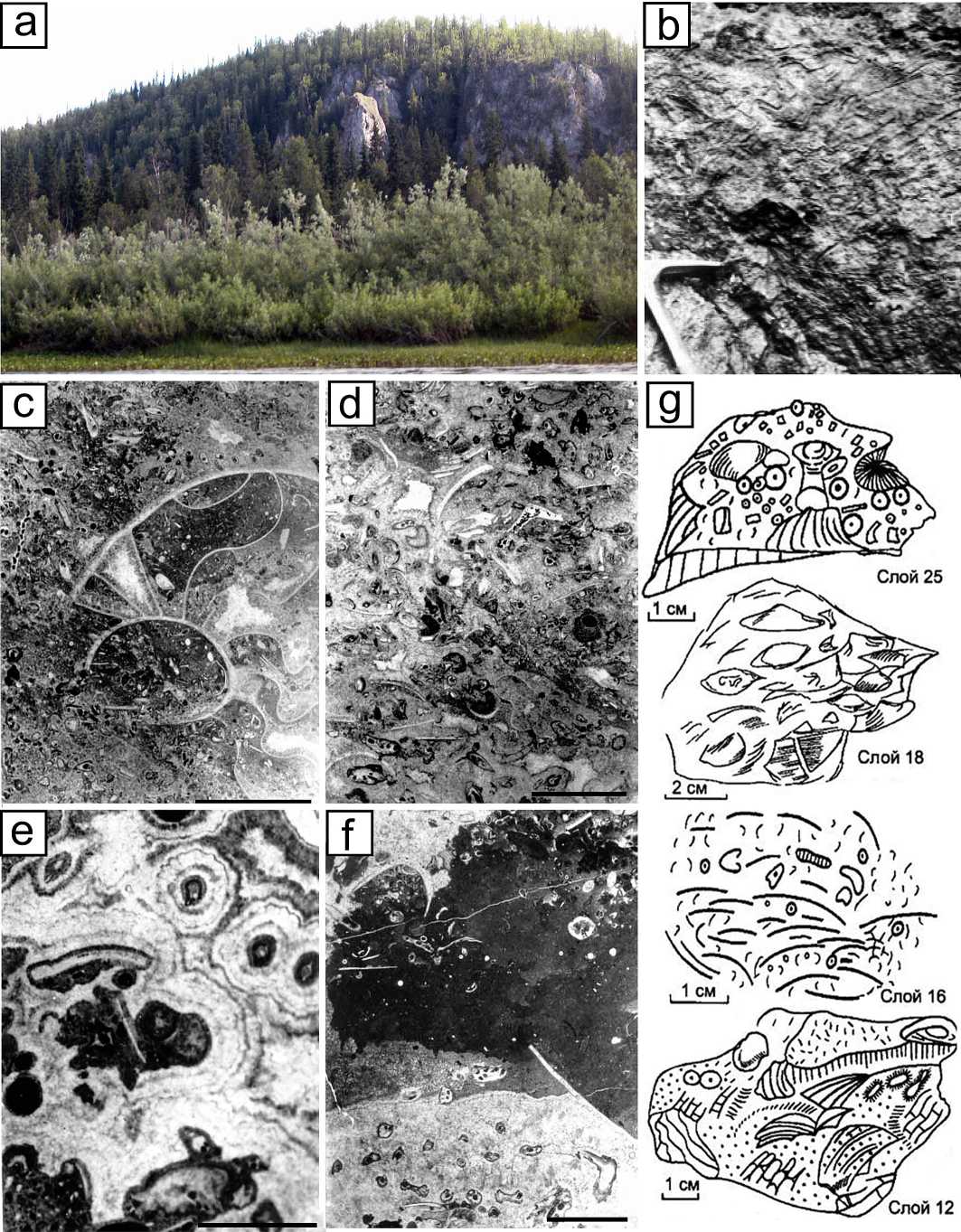
Рис. 3 . Характерные типы известняков скелетного холма Кожым:
а — выходы пород среднеассельской части разреза; b — фрагмент поверхности сакмарского палеоаплизинового биогерма в верхней части разреза; c—f — шлифы: c — микрозернистый известняк с несортированным биокластовым материалом с крупным фрагментом аммонита; d — мшанково-биокластовый известняк с тонкофибровой крустификацией вокруг мшанок, черное — примазки битума; e — мшанково-тубифитесовый биогермный известняк с биоиндуцированным крустификационным цементом в виде корок вокруг зооеций мшанок и трубочек Tubiphytes ; f — тубифитесово-мшанковый биогермный известняк с участками илового и микробиально-сгусткового кальцита; g — полевые зарисовки брахиоподовых ракушняков. Масштабная линейка — 1 мм
-
Fig. 3. Characteristic types of limestones of the Kozhym skeletal mound:
a — outcrops of the Mid-Assellian part in the section; b — a fragment of the surface of the Sakmarian Palaeoaplysina bioherm in the upper part of the section; c—f — thin sections: c — a micro-grained limestone with unsorted bioclastic material with a large fragment of ammonite; d — a bryozoan-bioclastic limestone with fine-fibrous crustification around bryozoans, black — smears of bitumen; e — a bryozoan- Tubiphytes bioherm limestone with bioinduced cement in the form of crusts around zooecia of bryozoans and tubules of Tubiphytes ; f — a Tubiphytes -bryozoan bioherm limestone with areas of silt and microbial-clotted calcite; g — field sketches of bra-chiopod shells. Scale bar — 1 mm

Рис. 4 . Характерные представители карбонатных пород, слагающих комплекс мшанковых биогермов Казантипа: a — мшанковые биогермные известняки в береговом клифе (бухта Широкая); b — межбиогермные мшанковые известняки (там же); c — фрагмент мшанкового биогерма с фоссилизированной биопленкой на мшанках и межкаркасным заполнением (Сенькина бухта); d — крупная мощная карбонатная корка со сложным минеральным составом на мшанковом биогермном известняке (бухта Шарабай); e — скопление несортированных фрагментов двустворок, участками отчетливо видна покрывающая их минерализованная биопленка (там же); f — СЭМ-изображение в режиме вторичных электронов строения минерализованной биопленки на мшанковом биогермном известняке (бухта Широкая)
-
Fig. 4. Typical representatives of carbonate rocks in the Kazantip complex of bryozoan bioherms:
a — a fragment of an outcrop of bryozoan biohermal limestones in the coastal cliff (the Shirokaya Bay); b — interbioherm beds of bryo-zoan limestones in the same place; c — a fragment of a bryozoan bioherm with fossilized biofilm on bryozoans and interframe filling (the Sen’kina Bay); d — a large carbonate crust with a varied mineral composition on bryozoan bioherm limestone (the Sarabaj Bay); e — an accumulation of unsorted fragments of bivalves, in some areas the mineralized biofilm covering them is clearly visible, in the same place; g — SEM-image in the mode of secondary electrons of the structure of a mineralized biofilm on bryozoan biohermal limestones (the Shirokaya Bay)
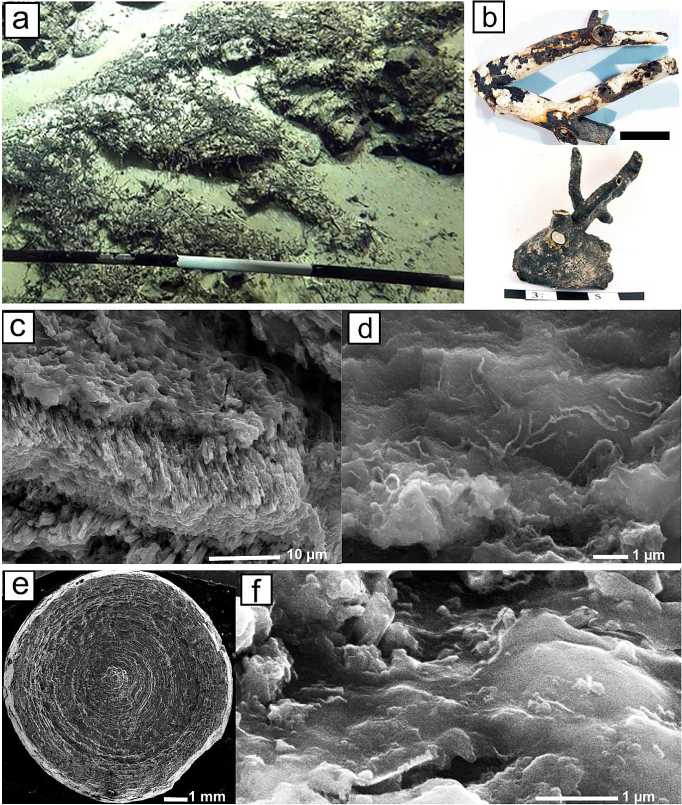
Рис. 5 . Глубоководные органогенно-карбонатные постройки САХ:
a — фрагмент видеосъёмки дна рядом с молодым вулканом, молодые базальты и туффиты лежат на осадках и покрыты твёрдыми бактериальными образованиями различных форм, преимущественно в виде палочек и веточек (съемка И. Г. Добрецовой); b — ветвистые формы построек; c—e — СЭМ-изображения во вторичных электронах: c — тонкофибровые кристаллы аутигенного Mg-кальцита под Fe-Mn-коркой с бактериоморфными структурами; d—f — типы бактериоморфных структур в железомарганцевых корках построек; e — зонально-концентрическая структура ветвистой постройки с Fe-Mn-коркой (светлая зона)
-
Fig. 5. Deep-sea organogenic-carbonate buildups of the MAR:
a — a fragment of video footage of the bottom near a young volcano, young basalts and tuffites lie on sediments, they are covered with solid bacterial formations of various forms, mainly in the form of sticks and twigs (video footage by I. G. Dobretsova); b — branched forms of buildups; c—e — SEM-images in secondary electrons: c — fine-fiber crystals of authigenic Mg-calcite under a Fe-Mn crust with bacte-riomorphic structures; d—f — types of bacteriomorphic structures in Fe-Mn crusts of buildups; e — zonal-concentric structure of a branched buildup with a Fe-Mn crust (light zone)
делах как от –3 до 3 ‰, так и от –2 до 2 ‰ [4, 14, 18, 20]. Установлено, что первичный изотопный состав углерода в морских карбонатолитах сохраняется, несмотря на перекристаллизацию и доломитизацию пород, если относительное содержание активной углекислоты в подземных водах было невелико [41]. В статье [39] результатами изучения фанерозойских брахиопод было подтверждено, что в большинстве случаев изотопный состав кислорода в кальците второго слоя раковин брахиопод вполне отражает условия их существования. Разница в значениях δ 18О морской воды и равновесного с ней δ 18О в карбонате близка к 30 ‰. С повышением температуры воды на 1 °С она уменьшается примерно на 0.2 ‰, к уменьшению значений δ 18О в карбонатах приводит также опреснение мор- 34
ской воды [3]. Исследованиями [51] доказывается, что изотопный состав кислорода морской воды на протяжении всего фанерозоя оставался стабильным благодаря существованию стационарного баланса между гидротермальной активностью на морском дне и выветриванием на земной поверхности. Изотопный состав кислорода морской воды в фанерозое принимается в пределах 28—30 ‰, что по Тейс и Найдину [27] приводит к оценке палеосолёности в 29.5—33.5 ‰. Снижение значения изотопного коэффициента по кислороду и соответственно уменьшение солёности морской воды может быть обусловлено как соответствующими изменениями обстановки осадкообразования в самом бассейне, например опреснением воды вследствие континентального речного стока, так и вторич-
Изотопный состав углерода и кислорода в карбонатном веществе исследованных органогенных построек Isotopic composition of carbon and oxygen in the carbonate matter of the investigated buildups
|
№ п/п |
δ 13С PDB |
δ 18O SMOW |
№ п/п |
δ 13С PDB |
δ 18O SMOW |
№ п/п |
δ 13С PDB |
δ 18O SMOW |
|
Риф Большая Косью Bolshaya Kosyu |
86/154 |
2.6 |
25.8 |
Шир/26а-18 |
1.92 |
28.79 |
|||
|
12/2-2 |
2.20 |
26.32 |
86/156 |
1.6 |
23.8 |
Шир/26b-18 |
2.69 |
29.03 |
|
|
12/2-3 |
1.52 |
25.56 |
86/163 |
1.7 |
24.0 |
Шир/30а-18 |
0.64 |
27.40 |
|
|
12/2-4 |
0.35 |
22.36 |
86/141 |
1.3 |
24.7 |
Шир/31-18 |
–0,15 |
24.20 |
|
|
11/13 |
2.90 |
27.45 |
86/164 |
1.9 |
25.2 |
Шир/30б-18 |
1,05 |
25.72 |
|
|
5/30-1 |
0.90 |
28.25 |
86/165 |
1.6 |
23.0 |
Шир/27-18 |
2.11 |
28.10 |
|
|
5/30-2 |
1.21 |
27.78 |
Ск S |
елетный хо keletal moun |
м d |
Шир/29-18 |
–0.32 |
26.35 |
|
|
5/44-1 |
1.24 |
26.53 |
пм/1 |
6.8 |
29.3 |
САХ Mid-Atlantic Ridge |
|||
|
5/44-2 |
1.09 |
26.99 |
пм/2 |
5.7 |
27.0 |
1 |
–0.29 |
33.03 |
|
|
5/44-3 |
1.73 |
23.78 |
пм/3 |
6.7 |
28.6 |
2 |
0.10 |
33.58 |
|
|
5/67-1 |
1.11 |
27.36 |
пм/4 |
6.0 |
27.8 |
3 |
–0.45 |
33.74 |
|
|
5/67-2 |
1.30 |
26.72 |
пм/6 |
6.8 |
29.3 |
4 |
–0.37 |
33.72 |
|
|
5/76-1 |
1.29 |
24.88 |
пм/7 |
5.7 |
27.0 |
5 |
–0.98 |
36.07 |
|
|
5/76-2 |
1.17 |
24.43 |
пм/8 |
6.7 |
28.6 |
6 |
–0.86 |
38.27 |
|
|
5/76-3 |
–0.33 |
22.24 |
пм/9 |
6.0 |
27.8 |
7 |
0.40 |
39.75 |
|
|
5/76-4 |
2.19 |
25.07 |
пм/10 |
6.9 |
28.6 |
8 |
–0.96 |
37.52 |
|
|
5/85-2 |
3.13 |
25.68 |
пм/11 |
6.9 |
28.5 |
9 |
–0.46 |
37.41 |
|
|
1/14-1 |
1.56 |
27.81 |
пм/12 |
7.1 |
28.3 |
10 |
–0.47 |
34.24 |
|
|
1/14-2 |
1.46 |
28.66 |
пм/4а |
6.2 |
26.6 |
11 |
0.26 |
35.49 |
|
|
5 |
1.50 |
28.84 |
пм/4б |
6.1 |
27.5 |
12 |
–0.71 |
34.14 |
|
|
20 |
1.30 |
28.10 |
пм/5а |
6.5 |
27.7 |
13 |
–0.47 |
34.24 |
|
|
27 |
1.39 |
29.52 |
пм/6б |
6.6 |
27.4 |
14 |
0.67 |
33.40 |
|
|
29 |
1.29 |
29.86 |
пм/38а |
6.8 |
30.0 |
15 |
0.78 |
32.72 |
|
|
101 |
1.54 |
27.50 |
пм/38б |
5.1 |
27.2 |
16 |
0.76 |
32.65 |
|
|
103 |
1.67 |
27.70 |
пм/38в |
5.6 |
25.2 |
17 |
0.83 |
32.56 |
|
|
105 |
1.58 |
27.73 |
пм/46а |
7.3 |
29.3 |
18 |
0.26 |
35.49 |
|
|
7 |
1.50 |
28.84 |
пм/46б |
5.6 |
26.4 |
19 |
–0.71 |
34.14 |
|
|
107 |
1.53 |
27.60 |
пм/46в |
5.6 |
25.4 |
20 |
0.40 |
32.15 |
|
|
109 |
1.37 |
27.49 |
пм/46г |
6.1 |
26.3 |
21 |
0.05 |
33.76 |
|
|
111 |
1.27 |
26.98 |
пм/47а |
5.5 |
26.4 |
22 |
0.13 |
32.80 |
|
|
113 |
1.89 |
27.92 |
пм/47б |
5.4 |
25.4 |
23 |
0.49 |
32.61 |
|
|
115 |
1.25 |
27.90 |
пм/47в |
5.1 |
22.5 |
24 |
0.82 |
32.27 |
|
|
231 |
0.68 |
28.45 |
пм/53а |
5.5 |
24.6 |
25 |
0.37 |
32.24 |
|
|
241 |
1.17 |
28.45 |
пм/53б |
6.5 |
26.6 |
26 |
0.67 |
33.40 |
|
|
219 |
0.83 |
28.98 |
пм/53в |
6.4 |
25.1 |
27 |
0.78 |
32.72 |
|
|
Микробиальный холм Microbial mound |
пм/53г |
6.4 |
25.8 |
28 |
0.76 |
32.65 |
|||
|
86/85 |
1.7 |
20.4 |
пм/53д |
5.7 |
26.5 |
29 |
0.83 |
32.56 |
|
|
86/91 |
1.6 |
22.6 |
пм/5 |
6.9 |
28.6 |
30 |
0.40 |
32.15 |
|
|
86/92 |
1.4 |
24.0 |
Казантип Kazantip |
31 |
0.05 |
33.76 |
|||
|
86/109 |
2.0 |
23.2 |
СБ/4-18 |
7.17 |
32.76 |
32 |
0.13 |
32.80 |
|
|
86/121 |
1.7 |
24.2 |
СБ/5-18 |
-1.40 |
26.49 |
33 |
0.49 |
32.61 |
|
|
86/122 |
2.2 |
24.1 |
СБ/6-18 |
3.97 |
31.14 |
34 |
0.82 |
32.27 |
|
|
86/123 |
2.0 |
24.6 |
СБ/8-18 |
4.51 |
30.22 |
35 |
0.37 |
34.24 |
|
|
86/126 |
2.0 |
23.2 |
Шар/2b-18 |
3.12 |
32.26 |
36 |
0.40 |
32.15 |
|
|
86/136 |
3.0 |
25.6 |
Шар/17-18 |
6.20 |
32.69 |
37 |
0.05 |
33.76 |
|
|
86/138 |
1.3 |
24.7 |
Шир/1к-18 |
4.22 |
32.12 |
38 |
0.13 |
32.80 |
|
|
86/139 |
1.3 |
26.0 |
Шир/1с-18 |
3.71 |
32.04 |
39 |
0.49 |
32.61 |
|
|
86/142 |
1.1 |
24.4 |
Шир/3-18 |
3.99 |
30.94 |
40 |
0.82 |
32.27 |
|
|
86/147 |
1.3 |
26.3 |
Шир/12-18 |
4.53 |
33.01 |
41 |
0.37 |
34.24 |
|

ными процессами, такими как перекристаллизация, доломитизация, карстование, протекающими с участием опресненных вод [14, 22].
Приведенные в таблице и на рис. 6 данные позволяют сделать следующие выводы.
Карбонатные породы рифа Большая Косью в целом характеризуются варьированием δ 13С в пределах нормы для морской воды — от –0.33 до 3.13 ‰, статистически (среднее ± СКО) это составляет (1.41 ± 0.63) ‰. Значения изотопного коэффициента по кислороду статистически колеблется в пределах δ 18О = (26.75 ± 2.87) ‰, что отвечает уровню палеосолености морской воды в 20—26.2 ‰. Последнее несколько уступает средней солёности морской воды по фанерозою (29.5—33.5 ‰). Между значениями изотопных коэффициентов углерода и кислорода обнаружена слабая положительная связь (r = 0.16), коэффициенты их вариации составляют соответственно 45 и 11 %, что свидетельствует об умеренной изотопной однородности. Имеющиеся отклонения изотопного состава углерода и кислорода от средних значений могут свидетельствовать о колебаниях количества органического вещества (ОВ) в осадке — основного источника изотопно-легкого углерода. В целом для рассматриваемого рифа можно предполагать образование в обстановках разной подвижности вод, солености и количества ОВ: рифового плато с отмелями (подвижная гидродинамика), межбиогерм-ного и межкаркасного пространства, а также внутри-рифовой лагуны (относительно спокойная гидродинамика). Более детальная углеродно-изотопная кривая по рифовой толще выявила семь уровней с различающимися значениями δ 13С, соответствующих разным стадиям развития рифовой экосистемы — стабилизации, колонизации, диверсификации и доминации, для
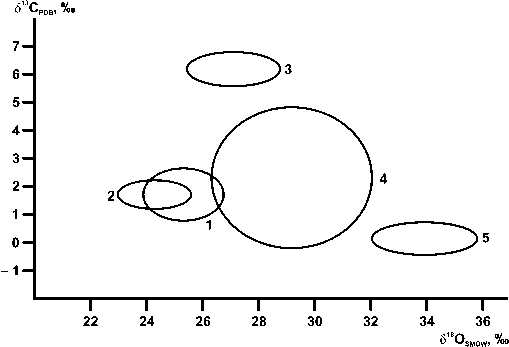
Рис. 6 . Статистические поля (среднее ± СКО) варьирования изотопного состава углерода ( δ 13С) и кислорода ( δ 18О) в карбонатах исследованных органогенных построек: 1 — риф Большая Косью, 2 — микробиальный холм Шаръю, 3 — скелетный холм Кожым, 4 — мшанковый биогермный комплекс Казантип, 5 — постройки в зоне рифтовой долины севера Срединно-Атлантического хребта
-
Fig. 6. Static fields (mean ± RMS) of variations in the isotopic composition of carbon ( δ 13С) and oxygen ( δ 18О) in carbonates of the studied organogenic buildups: 1 — Bol'shoj Kos'yu reef, 2 — Shar’yu microbial mound, 3 — Kozhym skeletal mound, 4 — Kazantip bryozoan biohermal complex, 5 — buildups in the rift valley zone of the northern Mid-Atlantic Ridge
которых характерны собственные типы организмов и карбонатных отложений [30, 31]. Смещение значений δ 18О в область 28.3—30.0 ‰ при массовом развитии рифостроящих каркасных организмов и разнообразных биогермных известняков отражает нормальноморские условия развития рифа на стадии диверсификации.
Изотопный состав углерода и кислорода в карбонатных породах микробиального холма Шаръю колеблется в пределах соответственно 0.8—3.0 и 20.4—26.3, что статистически дает (1.71 ± 0.51) и (24.23 ± 1.36) ‰. Коэффициенты вариации изотопных коэффициентов определяются в 30 и 6 %, что свидетельствует о несколько большей однородности вещества холма по сравнению с вышерассмотренным рифом. Парной корреляции между значениями изотопных коэффициентов не выявлено. Вариации значений δ 13СPDB в известняках франской части холма составляют 2.2 ‰ (0.8—3 ‰), а δ 18О — 2.5 ‰ (23.8—26.3 ‰), что соответствует выводу о значительной изотопной однородности карбонатного вещества. Для фаменских карбонатов характерны меньшие вариации величин δ 13С — 0.8 ‰ (1.6— 2.4‰), тогда как для кислорода они более существенны — 4.2 ‰ (20.4—24.6 ‰). В целом значения δ 13С, полученные для микробиального холма, соответствуют морским карбонатам, а значения δ 18O, напротив, заметно ниже типичных для нормально-морской обстановки. Последнее может свидетельствовать о формировании исследованного микробиального холма на мелководье и в опресненных условиях. В формировании каркаса микробиального холма важную роль играли кальцимикробы и цианобактерии, но при этом карбонаты аутигенного происхождения чередовались в разрезе биогерма с карбонатами аллохтонными, привнесенными в составе фрагментов фоссилий, пелитоморфного и микробиального материала. Самые низкие значения изотопного состава кислорода в пределах 20.4—23.2 ‰ характерны для карбонатов в псевдоконгломератах с палеомикрокодиями. Значения δ 18O и δ 13СPDB в раковинах брахиопод рода Cyrtospirifer , широко распространенного в верхнедевонских рифогенных толщах Тимано-Североуральского региона, соответствуют оценкам палеотемпературы в 6—22 °С, а солености вод — от 27 до 29 ‰ [7]. Последние значения на 3—3.5 ‰ ниже, чем таковые для других низкоширотных разрезов [53]. По данным [48], во франском веке в морских бассейнах Северной Америки, Европы и Австралии палеотемпература воды составляла 30 °C, а в течение фамена она лишь несколько снизилась. Палеосоленость морских вод по изотопным характеристикам брахиопод Cyrtospirifer в позднем девоне снизилась, что соответствует условиям крайнего мелководья на склонах карбонатных платформ с притоком пресных вод. Аналогичный результат был получен и при изучении верхнедевонских рифогенных толщ в Хорейверской впадине [19]. Отметим, что вариации палеотемператур и солености по разрезу вполне могли быть обусловлены чередованием трансгрессивно-регрессивных фаз развития бассейна, отраженных в строении микробиального холма сменой биогермных фаций и более мелководных фаций биокластовых песков и органогенных песчаников, вплоть до субаэральных [34].
Реконструкция палеотемператур и палеоклимата позднедевонской эпохи в целом вызывает многочис- ленные дискуссии [50]. Среднее значение δ18О в карбонатах микробиального холма Шаръю является аномально низким, свидетельствуя о специфике условий развития этой постройки. В этой связи действительно может идти речь о поступлении в морской бассейн речных вод, что уже обсуждалось в публикациях по верхнему девону [45, 46].
Результаты исследования биогермных и биокла-стовых известняков и раковин брахиопод скелетного холма Кожым показали высокие значения изотопного коэффициента δ 13С: для биогермных известняков 3.5—5.5 ‰, для биокластовых известняков 6.9—7.3 ‰, для раковин брахиопод 5.1—7.3 ‰. Статистические данные по холму составляют (6.18 ± 0.61) ‰. Это значительно превышает аналогичные характеристики нормально-морских карбонатов, свидетельствуя об испарительном режиме. Некоторое облегчение изотопного состава углерода от 5.5 до 3.5 ‰ в кальците биоцементолитов характеризует переход от цемента ранней генерации (радиально-лучистый кальцит) к цементу поздних генераций (блоковый кальцит). В па-леоаплизинах изотопный состав карбонатного углерода достигает 5—5.2 ‰ [37]. Разброс изотопных величин на углеродной кривой в биогермных разностях составляет 2 ‰, в биокластовых– 0.4 ‰, а в раковинах брахиопод — 2.2 ‰. Следовательно, мы видим больший размах колебаний величины δ 13С именно в аутигенных карбонатах, тогда как в биокластовых карбонатах она по углероду в 5—6 раз изотопно-однородней. Можно предположить, что образование биоинду-цированных цементов происходило в более изменчивых по физико-химическим параметрам условиях [43].
Значения изотопного коэффициента для карбонатного кислорода в рассматриваемом скелетном холме демонстрируют весьма широкий разброс по разрезу — от 22.5 до 30 ‰. При этом выявляется очень сильная положительная корреляция изотопного состава кислорода и углерода (r = 0.75). Статистические (среднее ± СКО) оценки δ 18О определяются как (27.1 ± 1.66) ‰. Известняки с величинами δ 18О 28—30 ‰ и соответствующими значениями солености 29.2—33.6 отвечают нормально-морским обстановкам со среднефане-розойской солёностью вод. Образцы карбонатов с более низкими значениями коэффициента δ 18О = 25— 27.7 ‰ показывают некоторое опреснение до значений солености 22.5—28.5 ‰. Самые низкие величины δ 18О = 22.5—24.6 ‰ в карбонатах соответствуют палеосолёности в 17.5—21.5 ‰, что указывает на довольно опресненные морские воды.
Изотопное исследование аммоноидей Среднего Урала показало, что палеотемпература вод в мелководных морских бассейнах составляла для гжельского времени 13.7—18.9 °С, а для сакмарского — 17.4—23.7 °С [10]. Это соответствует умеренно прохладному климату. Таким образом, формирование скелетного холма Кожым происходило в условиях глобального поздне-каменноугольного-раннепермского похолодания [25] на фоне уральских орогенных процессов, захвативших окраину карбонатной платформы [24]. Моря в это время характеризовались мезотрофностью вод и повышенной биопродуктивностью [25, 43], что объясняет существенно высокие значения изотопного состава углерода как в фоссилиях, так и в органогенных кар- бонатах — от 3.5 до 7.1 ‰. На это указывает и развитие в составе холма преимущественно сессильных организмов, биоиндуцированного цемента и обилие скелетной биокластики.
Исследования изотопного состава карбонатных пород биогермного комплекса Казантип показали максимальные вариации изотопных значений: δ 13C = = от –2.76 до 7.17 ‰; δ 18O = от 24.20 до 33.01 ‰. Статистические (средние ± СКО) данные составляют соответственно (2.31 ± 2.51) и (29.18 ± 2.86) ‰. При этом, судя по коэффициентам вариации, степень изотопной неоднородности по углероду оказалась в 10 раз выше, чем по кислороду (109 против 10 %). Корреляция между значениями изотопных коэффициентов здесь отсутствует.
В пределах комплекса карбонатные образования по величинам δ 13C и δ 18O подразделяются на три кластера. Первый кластер образуют пелитоморфные известняки с неравномерно распределенной биокласти-кой, вадозной пористостью, звездочками гематита и зернами глауконита: δ 13C = от –2.76 до –1.05 ‰; δ 18O = = 24.20—27.53 ‰; палеосолёность составляла 20.8— 28.15 ‰. Незначительность колебаний изотопного состава отражает относительную устойчивость условий образования карбонатов, характеризовавшихся обогащением осадков органическим веществом и опреснением морских вод. Второй кластер характеризуют биогермные мшанковые известняки с биоиндуциро-ванным цементом и гастроподами: δ 13C = 1.92—2.69 ‰; δ 18O = 28.19—29.03 ‰. Судя по приведенным значениям изотопного состава, условия образования мшанковых известняков были еще однообразнее, солёность морской воды была близка к среднефанерозойской. В третий кластер объединены гораздо более изотопно-неоднородные образования: карбонатные корки на мшанковых биогермах, моллюсково-полихетовые минибиогермы и известняки глинисто-доломитовые, вплоть до доломита со значениями δ 13С = 3.12—7.17 ‰; δ 18О = 30.22—33.01 ‰. Палесолёность морских вод оценивается в 33.8—41 ‰, что значительно превышает среднефанерозойские данные по морской воде.
Таким образом, проведенный изотопный анализ выявил отчетливые изменения условий карбонато-образования во время формирования биогермного комплекса Казантип. Породы первого кластера, распространенные на разных уровнях изученного разреза, образовались в обстановке мелководного моря с опресненной водой и с обогащением осадочной среды ОВ на фоне регрессии, неравномерного поступления терригенного материала с суши и проявления субаэральных процессов. Для пород второго кластера (средняя часть разреза) была характерна обстановка с солоноватой морской водой, позволявшая мшанкам строить каркас за счет раннедиагенетического биоин-дуцированного цемента, а гастроподам образовывать поселения. Породы третьего кластера, характеризующие разные интервалы разреза, формировались в более экстремальных условиях, сочетающих повышенную солёность морской воды и повышенный режим испарения. Электронно-микроскопические исследования карбонатных корок и моллюсково-полихетовых минибиогермов выявили в них фоссилизированные биопленки и нетипичную для нормально-осадочных карбонатов аутигенную минерализацию. Эти разноо- 37

бразные карбонаты формировались в обстановке мелководного внутриконтинентального морского бассейна с участием в осадкообразовании локальных газофлюидных высачиваний.
Голоцен-верхнеплейстоценовые глубоководные органогенно-карбонатные постройки Северной Атлантики характеризуются аномальными изотопно-геохимическими свойствами — сочетанием относительно изотопно-лёгкого углерода — δ 13С = от –0.96 до 0.83 ‰, характерного для нормально-морских карбонатов, и экстремально изотопно-тяжелого для морских обстановок кислорода — δ 18О = 32.24—39.75 ‰. Причем этих свойства практически тождественно характерны как для кальцитовых, так и для железомарганцевых карбонатов. Статистические оценки значений изотопных коэффициентов определяются как δ 13С = (0.14 ± 0.58) ‰ и δ 18О = (33.92 ± 1.88) ‰. Между значениями изотопных коэффициентов обнаруживается сильная отрицательная корреляция (r = –0.57).
Из приведенных данных видно, что в карбонатах построек пририфтовой долины с активным вулканизмом сочетается весьма большая изотопная неоднородность по углероду (коэффициент вариации — 414 %) с редко встречающейся изотопной однородностью по кислороду (5.5 %). Соленость среды образования этих карбонатов определяется в 38.6—48.2 ‰, что на 30— 45 % превышает среднефанерозойскую соленость морских вод. Таким образом, обнаруживается, что океанические пририфтовые органогенно-карбонатные постройки характеризуются принципиально разными изотопными типами углерода и кислорода — нормально-морским для углерода и аномальным изотопно-тяжелым для кислорода. Эта разнотипность объясняется тем, что углерод в карбонатах рассматриваемых построек имеет нормально-морское происхождение, а вот кислород по происхождению, возможно, не бикарбо-натный, а сульфатный, обусловленный жизнедеятельностью особых микроорганизмов.
Заключение
По результатам изотопных ( δ 13C, δ 18O) исследований карбонатного вещества органогенных построек можно сделать следующие выводы.
Генетическое разнообразие объектов исследования, установленное литолого-палеоэкологическими и химико-физическими методами, нашло свое подтверждение и обоснование в изотопных величинах углерода и кислорода, палеосолености и палеотемпературах.
Известняки среднекатийского рифа Большая Косью как сложно построенного рифового комплекса характеризуются существенным варьированием величин δ 13СPDB: от –0.33 до 3.13‰ и δ 18ОSMOW — от 22.5 ‰ до 30.0 ‰. Значения δ 13С и δ 18О, выходящие за пределы области нормально-осадочных морских карбонатов, соответствуют условиям внутририфовой лагуны. В область нормально-осадочных морских карбонатов попадают карбонаты с наивысшим таксономическим, морфологическим и количественным распространением каркасостроящих организмов.
В карбонатных породах среднефранско-фамен-ского микробиального холма Шаръю изотопный состав углерода и кислорода колеблется от 0.8 до 3.0 ‰ и от 20.4 до 26.3 ‰ соответственно. Факторами, обу- 38
словившими низкие значения δ 18OSMOW, были поступление изотопно-легких вод с речным стоком (обмеление в периоды регрессии вплоть до субаэральных) и аридизация климата в связи с похолоданием в фа-менское время.
Известняки и раковины брахиопод гжельско-сак-марского скелетного холма Кожым показали высокие значения δ 13СPDB — от 3.5 до 7.3 ‰ и варьирование значений δ 18OSMOW в пределах 7.5 ‰ (22.5—30.0 ‰). Среднеассельские известняки и брахиоподы с величинами 27.5—30 ‰ характеризуют область нормальноосадочных морских карбонатов. А верхнегжельские и сакмарские карбонаты и брахиоподы (22.5—27.4 ‰) — морские условия с меняющейся соленостью и температурой. Утяжеление значений δ 13С и δ 18O в карбонатах и брахиоподах могло быть связано как с ингресси-ями морских вод, насыщенных ОВ, так и усилением процессов испарения при аридизации климата.
Образование кластеров карбонатных пород био-гермного комплекса Казантип с отличающимися значениями δ 13C и δ 18O отражают группы карбонатов по химическому составу и условиям образования в полу-изолированном мелководном бассейне с колебанием уровня моря и, соответственно, солености и температуры: опресненный морской водоем δ 13CPDB (–2.76— 1.05 ‰) и δ 18OSNOW (24.20—27.5 3‰), морской бассейн с солоноватыми водами δ 13CPDB (1.92—2.69 ‰) и δ 18OSNOW (28.19—29.03 ‰) и бассейн с влиянием газофлюидных высачиваний и усилением испарительного режима: δ 13CPDB (3.12—7.17 ‰) и δ 18OSNOW (30.22— 33.01 ‰).
Изотопный состав δ 13C и δ 18O карбонатов глубоководных органогенно-карбонатных построек севера Срединно-Атлантического хребта показал, что они характеризуются однородностью, но имеют принципиально разные изотопные типы углерода и кислорода. Изотопный состав углерода кальцитовых тел построек не выходит за пределы интервала значений (–1… +1 ‰) морских осадочных карбонатов, но имеет аномально изотопно-тяжелые значения кислорода (33— 40 ‰); в железомарганцевых карбонатах — значения –3…+1‰ и 32—35 ‰ соответственно. Абсолютное отсутствие корреляции изотопных составов в единой постройке могло быть связано с особенностями изотопного обмена в придонных водах внеосевой зоны Срединно-Атлантического хребта.
Работа выполнялась в рамках государственного задания ИГ Коми ФИЦ НЦ УрО РАН (номер государственной регистрации № AAAA-A17-117121270034-3) и государственного задания ФГУН ИГГ УрО РАН (номер государственной регистрации АААА-А18-118053090045-8).
Авторы признательны рецензентам за полезные рекомендации по расшифровке аналитических данных и подготовке статьи к публикации.
Список литературы Генетически разные органогенные постройки в изотопных величинах D13С и D18О
- Антошкина А. И. Раннепалеозойское рифообразо-вание на севере Урала как пример его взаимосвязи с геобиосферными изменениями // Рифогенные формации и рифы в эволюции биосферы. М.: ПИН РАН, 2011. С.116— 141. (Геобиологические системы в прошлом).
- Антошкина А. И., Добрецова И. Г., Силаев В. И., Киселёва Д. В., Чередниченко Н. В. Особенности состава и строения карбонатных построек в северной зоне Срединного Атлантического хребта // Геология рифов: Всерос. литолог. совещ. Сыктывкар: Геопринт, 2020. С. 19—22.
- Виноградов В. И. Изотопный состав углерода и кислорода венд-кембрийских карбонатных пород и палеоэкологические реконструкции // Литол. и полез. ископ. 2008. № 1. С. 51—65.
- Галимов Э. М. Природа биологического фракционирования изотопов. М.: Наука, 1981. 247 с.
- Голубцов В. А., Черкашина А. А., Пустовойтов К. Е., Штар К. Стабильные изотопы углерода и кислорода педо-генных карбонатных кутан в черноземах Южного Прибайкалья как индикаторы локальных экологических изменений // Почвоведение. 2014. № 10. С. 1215—1227.
- Добрецова И. Г., Оськина Н. С. О взаимодействии базальтов с карбонатными осадками в районе 13—20° с.ш. Срединно-Атлантического хребта // ДАН. 2015. Т. 461. № 3. С. 307—311.
- Дорофеева Л. А., Сорикан Е. В., Малковский К. Изотопно-геохимические данные раковин брахиопод ВосточноЕвропейской платформы: значение для реконструкции условий формирования и корреляции позднедевонских отложений // Геология девонской системы: Материалы меж-дунар. симпоз. Сыктывкар, 2002. С. 153—156.
- Дубинина Е. О., Крамчанинов А. Ю., Силантьев С. А., Бортникова Н. С. Влияние скорости осаждения на изотопный состав (518О, 513С и S88Sr) карбонатов построек поля Лост-Сити (Срединно-Атлантический хребет, 30° с.ш.) // Петрология. 2020. Т. 28. № 4. С. 413—430.
- Захаров Ю. Д., Соколова Е. А., Смышляева О. П., Шигэта Я, Танабэ К., Маэда Х., Веливецкая Т. Г., Попов А. М., Игнатьев А. В., Афанасьева Т. Б. Новые данные по изотопам кислорода и углерода органогенных карбонатов и проблема низких изотопных палеотемператур тропиков в маастрихте // Тихоокеан. геолог. 2004. Т. 23. № 4. С. 54—72.
- Захаров Ю. Д., Борискина Н. Г., Попов А. М. Реконструкция условий морской среды позднего палеозоя и мезозоя по изотопным данным (на примере севера Евразии). Владивосток: Дальнаука, 2001. 112 с.
- Захаров Ю. Д., Смышляева О. П., Попов А. М., Шигэта Я. Изотопный состав позднемезозойских органогенных карбонатов Дальнего Востока (стабильные изотопы кислорода и углерода, основные палеоклиматические события и их глобальная корреляция) // Владивосток: Дальнаука, 2006. 204 с.
- Клюкин А.А. Факторы, определяющие биоразнообразие Казантипского природного заповедника // Тр. Никитского ботанич. сада — нац. науч. центра. 2006. Т. 126. С. 133—148.
- Кравчишина М. Д., Леин А. Ю., Саввичев А. С., Рейхард Л. Е., Дара О. М., Флинт М. В. Аутигенный Mg-кальцит на метановом полигоне в море Лаптевых // Океанология. 2017. Т. 57. № 1. С. 194—213.
- Кулешов В.Н. Эволюция изотопных углекислот-но-водных систем в литогенезе. Сообщение 1. Седименто-генез и диагенез // Литол. и полез. ископ. 2001. № 5. С. 491— 508.
- Кулешов В. Н., Арефьев М. П., Покровский Б. Г. Изотопные особенности (813С, 518О) континентальных карбонатов из отложений рубежа перми и триаса северо-востока Русской плиты: палеоклиматические и биотические причины, хемостратиграфия // Литол. и полез. ископ. 2019. № 6. С. 568—591.
- Леин А. Ю., Галкин С. В., Масленников В. В., Богданов Ю. А., Богданова О. Ю., Дара О. М., Иванов М. В. Новый тип карбонатных пород на дне океана (Срединно-Атлантический хребет, 30°07' с.ш.) // ДАН. 2007. Т. 412. № 4. С. 535-539.
- Логвинова Е. А., Матвеева Т. В. Сравнение изотопного состава аутигенных карбонатов из различных районов мирового океана // Вестник СПбГУ. 2009. Сер. 7. Вып. 1. С. 48-56.
- Маслов А. В. Осадочные породы: методы изучения и интерпретации полученных данных: Учеб. пособие. Екатеринбург, 2005. 289 с.
- Матвеева Н. А. Условия образования фаменских рифогенных отложений Центрально-Хорейверского вала Тимано-Печорской провинции // Нефтегазовая геология. Теория и практика. 2017. Т. 12. № 2. URL: http://www.ngtp. ru/rub/2/16_2017.pdf
- Мизенс Г. А., Кулешов В. Н., Степанова Т. Н. Первые сведения об изотопном составе углерода и кислорода в каменноугольных известняках восточного склона Южного Урала // Литосфера. 2008. № 4. С. 4-10.
- Параев В. В., Молчанов В. И., Еганов Э. А. Парадоксальность теории эволюции органического мира и тенденции ее решения // Уральский геолог. журн. 2009. № 4б. Вып. 70. С. 3-30.
- Покровский Б. Г., Мележик В. А., Буякайте М. И. Изотопный состав C, O, Sr и S в позднедокембрийских отложениях патомского комплекса, Центральная Сибирь. Сообщение 1. Результаты, изотопная стратиграфия и проблемы датирования // Литология и полезные ископаемые. 2006. № 5. С. 505-530.
- Преображеский Б. В. Современные рифы. М.: Наука, 1986. 244 с.
- Пучков В. Н. Геология Урала и Приуралья (актуальные вопросы стратиграфии, тектоники, геодинамики и металлогении). Уфа: Дизайн ПолиграфСервис, 2010. 280 с.
- Реймерс А. Н., Алексеев А. С., Ермакова Ю. В. Позднекаменноугольно-раннепермские климатические колебания и биотические события // Бюл. Моск. об-ва исп-лей природы. 2013. Т. 88. Вып. 1. С. 41-48.
- Садыков С. А., Потапов С. С. Изотопный состав углерода в карбонатных спелеотемах // Литосфера. 2011. № 5. С. 102-110.
- Тейс Р. В., Найдин Д. П. Палеотермометрия и изотопный состав кислорода органогенных карбонатов. М.: Наука, 1973. 256 с.
- Шмелёва Л. А. Палеоландшафтная реконструкция верхнеордовикского рифа р. Б. Косью (р. Илыч, Северный Урал): 23-я науч. конф. Сыктывкар: Геопринт, 2014. С. 146149.
- Шмелёва Л. А. Литолого-палеоэкологическая характеристика верхнеордовикского рифа Большая Косью (бассейн р. Илыч, Северный Урал) // Литосфера. 2016. № 1. С. 154-162.
- Шмелёва Л. А. Биоразнообразие верхнеордовикского рифа Большая Косью, Северный Урал // Вестник ИГ Коми НЦ УрО РАН. 2018. № 11. С. 38-42.
- Шмелёва Л. А. Изотопно-геохимические вариации как отражение геобиологических факторов в эволюции позднеордовикской рифовой экосистемы Большая Косью (р. Илыч, Северный Урал) // Геологический вестник. 2020. № 2. С. 94-109.
- Эволюционный тренд палеозойской рифовой экосистемы как отражение эволюции геобиологических систем на примере севера Урала / Антошкина А. И., Пономаренко Е. С., Рябинкина Н. Н., Валяева О. В., Лютоев В. П., Салдин В. А, Сандула А. Н, Канева Н. А. Сыктывкар: Геопринт, 2010. 44 с. (Отчетная серия № 2 (83)).
- Anadon P., Utrilla R. Sedimentology and isotope geochemistry of lacustrine carbonates of the Oligocene Campins Basin, north-east Spain. Sediment., 1993, V. 40, pp. 699—720.
- Antoshkina A. I. Palaeoenvironmental implications of Palaeomicrocodium in Upper Devonian microbial mounds of the Chernyshev Swell, Timan-northern Ural Region. Facies, 2006, V. 52, pp. 611—625.
- Antoshkina A. Justification of the unit boundaries by isotope data in the Gzhelian-Sakmarian carbonate massif in the Subpolar Urals / Kazan Golovkinsky Stratigraphic Meeting, 2017: Advances in Devonian, Carboniferous and Permian Research: Stratigraphy, Environments, Climate and Resources. Filodiritto Intern. Procs., 2018. pp. 19—23.
- Antoshkina A. I., Leonova L.V., Simakova Yu. S. The Development of Miocene Biohermal Bryozoan Limestones of Kazantip Cape (Crimea): A New Insight. Doklady Earth Sc., 2020, V. 49. Part 2, pp. 195—198.
- 37.Antoshkina A. I., Ponomarenko E. S. Bioinduced Crustification as a Reaction of Late Carboniferous-Early Permian Reef Ecosystems to Biosphere Changes.Paleont. J., 2014, V. 48. no. 14. pp. 1565—1576.
- Bekker A., Holmden C., Beukes N. J., Kenig, F., Eglinton, B., Patterson, W. P. Fractionation between inorganic and organic carbon during the Lomagundi (2.22—2.1 Ga) carbon isotope excursion. Earth and Planetary Sc. Let., 2008, V. 271. No 1—4, pp. 278—291.
- Brand U., Logan A., Hiller N., Richardson J. Geochemistry of modern brachiopods: applications and implications for oceanography and paleoceanography. Chem. Geol., 2003, V. 198, pp. 305—334.
- Copper P. Reefs under stress: the fossil record. Cour Forsh Senc, 1994, V. 72, pp. 87—94.
- Halverson G. P., Hoffman P. F., Schrag D. P., Maloof A. C., Rice A. H. N. Toward a Neoproterozoic composite carbonisotope record. Bull. Geol. Soc. Amer., 2005, V. 117, no. 9/10, pp. 1181—1207.
- lngram B. L., lngle J. C., Conrad M. E. A 2000 yr record of Sacramento-San Joaquin river inflow to San Francisco Bay estuary, California. Geology, 1996, V. 24, no. 4, pp. 331—334.
- FlügelE. Microfacies of carbonate rocks: analysis, interpretation and application. Berlin Heidelberg SpringerVerlag, 2010, 976 p.
- Gorshkov V. G., Makarieva A. M., Gorshkov V. V.Revising the fundamentals of environmental knowledge: the biota-environment interaction. Ecolog. Compl., 2004, V. 1, P. 17—36.
- Kaiser, S. I., Becker, R. T., Hassani, A. E. Middle to Late Famennian successions at Ain Jemaa (Moroccan Meseta) — implications for regional correlation, event stratigraphy and synsedimentary tectonics of NW Gondwana / R.T. Becker, W.T. Kirchgasser (eds.). Devonian Events and Correlation. Geological Society, London, Spec. Publ., 2007, V. 278, pp. 237—260.46. Lakin J. A., Marshall J. E. A., Troth I. Harding I. C. Greenhouse to icehouse: A biostratigraphic review of latest Devonian-Mississippian glaciations and their global effects. Geological Society, London, Special Publications, 2016, V. 423, No 1, 439—464.
- James N. P. Bourque P. A. Reefs and Mounds / R.G. Walker, N.P. James (eds). Facies Models -Response to Sea-Level Change. Geological Association of Canada. 1992, pp. 323—347.
- Joachimski M. M., Breisig S., Buggisch W. Talent J. A., Mawson R., Gereke M., Morrow J. R., Day J, Weddige K.. Devonian climate and reef evolution: Insights from oxygen isotopes in apatite. Earth and Planetary Science Let., 2009, V. 284, pp. 599— 609.
- Oehlert A. M., Swar P. K. Rolling window regression of S13C and 818O values in carbonate sediments: Implications for source and diagenesis. Depositional Record. 2019, V. 5, P. 613— 630.
- Racki G. A volcanic scenario for the Frasnian-Famennian major biotic crisis and other Late Devonian global changes: More answers than questions? Global and Planetary Change, 2020, V. 189, pp. 103—174.
- Ryb U., Eiler J. M. Oxygen isotope composition of the Phanerozoic ocean and a possible solution to the dolomite problem. PNAS, 2018, V. 115, no. 26, pp. 6602—6607.
- Sarangi V., Kumar A., Sanyal P. Effect of pedogenesis on the stable isotopic composition of calcretes and n-alkanes: Implications for palaeoenvironmental reconstruction. Sedimentology, 2019, V. 66, pp. 1560—1579.
- Van Geldern R., Joachimski M .M., Day J., Jansen U., Alvarez F., Yolkin E. A., Ma X.-P. Carbon, oxygen and strontium isotope records of Devonian brachiopods shell calcite. Palaeogeogr., Palaeoclimat., Palaeoecol., 2006, V. 240, P. 47—67.
