Генетические аспекты модернизации молочного скотоводства

Автор: Шендаков А.И., Шендакова Т.А.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Животноводство
Статья в выпуске: 2 (17), 2009 года.
Бесплатный доступ
В статье дан подробный анализ иммуногенетических особенностей молочных и комбинированных пород скота, наиболее распространённых в России. Затронуты вопросы селекции по генам и антигенам.
Короткий адрес: https://sciup.org/147123384
IDR: 147123384 | УДК: 636.2.03
Текст научной статьи Генетические аспекты модернизации молочного скотоводства
ФГОУ ВПО Орел ГАУ
ГЕНЕТИЧЕСКИЕ АСПЕКТЫ МОДЕРНИЗАЦИИ МОЛОЧНОГО СКОТОВОДСТВА
В статье дан подробный анализ иммуногенетических особенностей молочных и комбинированных пород скота, наиболее распространённых в России. Затронуты вопросы селекции по генам и антигенам.
В современной селекции сельскохозяйственных животных активно обсуждается вопрос об информативности эритроцитарных антигенов как маркеров высокой продуктивности в сравнении с ДНК-анализом. Доказано, что при небольших генетических различиях между внутрипородными структурами более информативны микросателлиты [10]. Однако в литературе также приводятся данные о различных причинах корреляции между группами крови и продуктивностью животных – плейотропии, сцеплении генов и гетерозисе [1]. Известно, что гены, кодирующие систему EAF (Erythrocyte Antigen F), находятся на 15 хромосоме, а открытие в этой системе антигена V′ позволяет лучше понять сущность эволюции скота [4].
При этом многие породы скота родственного происхождения имеют высокий индекс иммуногенетического сходства ( r i =0,64-0,89), хотя у черно-пёстрых и красно-пёстрых голштинов обнаружено существенное отличие по концентрации эритроцитарных антигенов B2, I1, I2, T1, Y1, E′2, G′, I′, I′ 2 , Q′, C 2 , W, X 1 , X 2 , L′, V, S 1 и S 2 . По данным С. Исламовой и др., у симменталов отсутствуют антигены P2, Y1, A′2, P′2, а T2 имеет наибольшую концентрацию [6], однако в Башкортостане также часто встречаются факторы W и F, не обнаружены Z′, R 1 , X 1 , M′, V′ 1 , R′ и пр. [14]. Эритроцитарные антигены B′, I′, Q′, T и С′ позволяют увеличить срок продуктивного использования палево-пёстрых коров на комплексе [2]. Факторы С′, C2, E, L, L′, R2, X1 и X2 встречаются у чёрно-пёстрых коров, выбраковываемых и по старости, и по причинам различных заболеваний, в то время как B2, H′, L – по причине мастита, С, R и X′ 2 – у коров с заболеваниями конечностей, С 1 – у больных лейкозом. Частота антигена A 1 у коров-долгожительниц составляет 0,0566, а при болезнях сердца, эндометрите и пневмонии – до 0,1818 [9]. Вместе с тем большинство научных работ в настоящее время посвящено изучению возможности маркировки тех или иных качеств у коров (высокого удоя, долголетия, заболеваемости и пр.), остаются недостаточно исследованными эволюционные процессы в породах, маркерная способность антигенов и генетических кластеров, сходство по аллелям сложных локусов и пр.
В связи с активным внедрением в скотоводство новых методов биотехнологии [7, 11, 16] исследованиями многих авторов доказано, что технологические свойства молока зависят от их генотипа по локусу гена каппа-казеина. Молоко коров с генотипом АВ и ВВ под действием сычужного фермента свёртывается раньше, чем молоко коров с генотипом АА. Используя быков, в геноме которых содержится аллель В каппа-казеина (генотип АВ и ВВ), можно не только поддерживать, но и увеличивать концентрацию аллеля В у маточного поголовья [8, 12, 13, 15]. Однако в настоящее время недостаточно изученным остаётся вопрос изменения 30
величины удоев при повышении концентрации гена В в стадах молочного скота.
Материалы и методика исследований
Для решения этих проблем под руководством ИНИЦ и лаборатории генетики Орловского ГАУ нами были начаты комплексные исследования групп крови у молочных и комбинированных пород крупного рогатого скота, наиболее распространённых в России. По каталогу быков-производителей ОАО «ЦСИО» нами был изучен антигенный состав и концентрация аллелей гена k-Cas у 8 пород: чёрно-пёстрой голштинской, айрширской, чёрно-пёстрой, симментальской, швицкой, красно-пёстрой голштинской, джерсейской и холмогорской (56, 16, 39, 7, 22, 8, 5 и 8 голов, соответственно). Частота встречаемости эритроцитарных антигенов (pi) была определена по методике Е. К. Меркурьевой, сходство по аллелям локусов групп крови (ri) – по Животовскому, общее иммуногенетическое сходство – по Майялу и Лингстрему, генетическая дистанция – по Нагаки. При определении достоверности силы влияния в однофакторном дисперсионном анализе применялся критерий Фишера [5]. Стандартная статистическая обработка данных проходила в компьютерной программе «Microsoft Excel».
Результаты и их обсуждение
Из рисунка 1 следует, что из 73 антигенов их наибольшее число было характерно для быков-производителей чёрно-пёстрой голштинской и швицкой пород – 59 и 46, соответственно, (в целом 798 и 264 положительной реакции гемолиза). Быки остальных пород отличались меньшим разнообразием антигенного состава – 33-36, что в отдельных случаях объяснимо небольшим поголовьем. Чёрно-пёстрая порода, несмотря на большую численность быков, также имела низкое иммуногенетическое разнообразие.
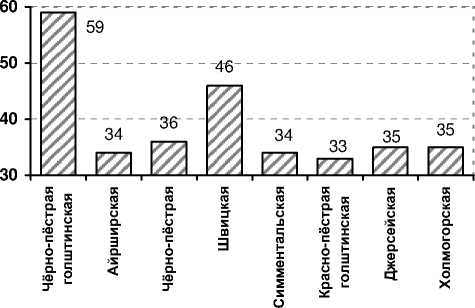
Рис. 1. Иммуногенетическое разнообразие быков-производителей молочных и комбинированных пород ОАО «ЦСИО»
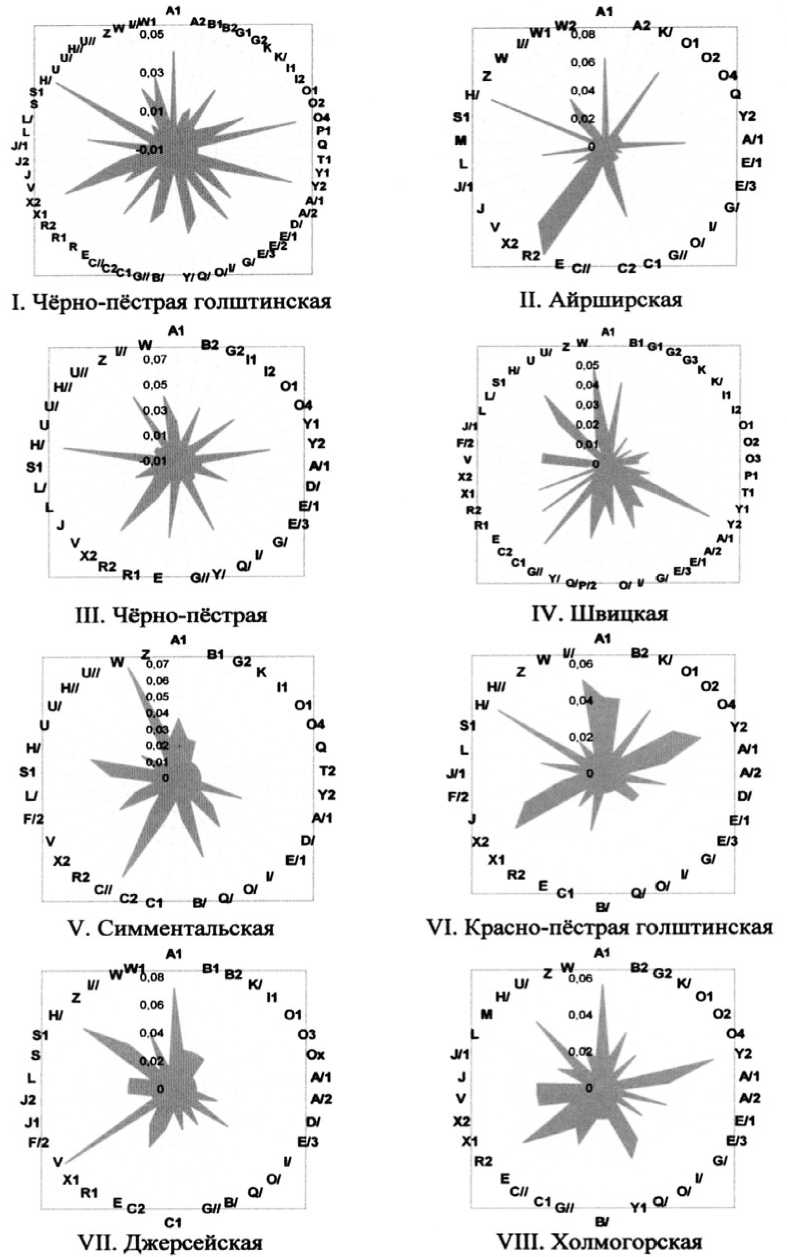
Рис. 2. Частота встречаемости эритроцитарных антигенов у быков молочных и комбинированных пород ОАО «ЦСИО»
Из рисунка 2 следует, что у всех исследованных пород наибольшую концентрацию показали разные антигены. У чёрно-пёстрых голштинских быков-производителей (I) выделялись эритроцитарные факторы A 1 , O 4 , Y 2 , X 2 и H′ – 0,04130, 0,04887, 0,04762, 0,04511 и 0,05514, соответственно. Антигены G 2 , O 1 , E′ 3 , Q′ и L имели частоту 0,02631, 0,02631,
0,03383, 0,03383 и 0,03007, соответственно. У айрширских быков (II) наблюдалось преимущество по A 1 , O 1 , A′ 1 , С′, R 2 , X 2 и H′ – 0,06286, 0,06286, 0,05143, 0,05143, 0,08571, 0,05634 и 0,08000, соответственно. Чёрно-пёстрые животные (III) отличались высокой частотой встречаемости антигенов G 2 , O 1 , Y 2 , Q′, E, X 2 , H′, Z и W – 0,03521,
0,04225, 0,05634, 0,04930, 0,04930, 0,05634, 0,07042, 0,04930 и 0,04225, соответственно. Наибольшая частота встречаемости у швицкого скота (IV) была характерна для факторов B1, Y2, C1, H′ и W – 0,04167, 0,05303, 0,04924, 0,04545 и 0,04924, соответственно. Антигены Q′, С′′, H′ и W у симменталов (V) характеризовались концентрацией 0,04938-0,07407, в то время как красно-пёстрые голштинские быки (VI) имели предпосылку к идентификации по антигенам A 1 , B 2 , O 1 , O 4 , Y 2 , X 2 , J, H′ и I′′ – 0,04166-0,06250. У джерсейского скота (VII) наибольшее значение проявили факторы A1, V и H′ (до 0,08695), у холмогорского (VIII) – A1, Y2, Q′, R2 и H′ (до 0,05691). При этом все породы имели высокую концентрацию антигенов F и H′.
Из таблицы 1 следует, что для чёрно-пёстрой голштинской породы специфическими являются антигены E′ 2 и R, для швицкой – G 3 и P′ 2 , для симментальской – T 2 , для джерсейской – O x и J 1 . У чёрно-пёстрой, красно-пёстрой голштинской и холмогорской пород специфические антигены обнаружены не были, в то время, как только у айрширов не было антигена Q′, только у чёрнопёстрого скота – O′, у джерсеев – J2, E′1, R2 и X2, у холмогорского скота – S1. При этом некоторые антигены отсутствовали у всех пород – Z′, P2, I′2, B′′ и S2, у большинства пород нет антигенов A2, P1, E′2, T1, R, J 2 , J′ 2 , M, S, W 2 и др.
Из таблицы 2 следует, что наибольшее сходство по аллелям локуса EAA было получено между джерсейской и айрширской, джерсейской и холмогорской, холмогорской и айрширской – 0,067, 0,064 и 0,060, соответственно. Высокое сходство наблюдалось у джерсеев с чёрно-пёстрым и краснопёстрым голштинским скотом – 0,055, а также у чёрно-пёстрых и красно-пёстрых голштинов с айрширами, джерсеев и симменталов (0,052).
По системе EAB выделялось сходство чёрнопёстрых голштинов с холмогорскими, швицкими и красно-пёстрыми голштинскими быками – 0,333, 0,319 и 0,315, соответственно. Низкое сходство было у джерсейского и чёрно-пёстрого скота – 0,146, а также у айрширского с чёрно-пёстрым и симментальским – 0,154.
Локус EAС отличался высоким сходством холмогорских и айрширских, чёрно-пёстрых голштинских и айрширских, айрширских и швицких быков – 0,219, 0,183 и 0,178, соответственно. Невысокое сходство показали симменталы с краснопёстрым голштинским и чёрно-пёстрым скотом (0,050-0,052). Большинство пород имели средний индекс сходства.
В системе EAJ средним сходством отличались холмогорские и айрширские, чёрно-пёстрые голштинские и айрширские, холмогорские и чёрнопёстрые голштинские животные – 0,027, 0,023 и 0,021, соответственно, высоким – айрширский и красно-пёстрый голштинский скот (0,039), холмогорский и красно-пёстрый голштинский (0,037), чёрно-пёстрый голштинский и красно-пёстрый голштинский (0,032). По большинству сочетаний сходство не было обнаружено или было низким (0,006-0,012).
В EAL-локусе выделялось сочетание чёрнопёстрых голштинов со швицким и айрширским скотом (0,036 и 0,035), красно-пёстрых голштинских быков с айрширами – 0,035, чёрно-пёстрых голштинских и красно-пёстрых голштинских, холмогорских и айрширских быков (0,031). Не выявилось сходство симментальских быков-производителей с айрширами, красно-пёстрыми голштинами, джерсейской и холмогорской породой.
Таблица 1 – Специфические и нехарактерные антигены у быков-производителей
|
Порода |
Специфические антигены |
Нехарактерные антигены |
|
Чёрно-пёстрая голштинская |
E′ 2 , R |
Z′, G 3 , O 3 , O x , P 2 , T 2 , I′ 2 , P′ 2 , B′′, J 1 , S 2 , W 2 |
|
Айрширская |
W 2 |
Q′ (нет только у айрширов); Z′, B1, B2, G1, G2, G3, K, I1, I2, O3, O x , P 1 , P 2 , T 1 , T 2 , Y 1 , A′ 2 , D′, E′ 2 , I′ 2 , P′ 2 , B′′, B′, R, R 1 , X 1 , J 1 , J 2 , L, S, S 2 , U, U′, H′′, U′′ |
|
Чёрно-пёстрая |
Не обнаружены |
O′ (нет только у чёрно-пёстрой породы); A 2 , Z′, B 1 , G 1 , G 3 , K, K′, O 2 , O 3 , O x , P 1 , P 2 , T 1 , T 2 , Q, A′ 2 , E′ 2 , I′ 2 , P′ 2 , B′′, B′, C 1 , C 2 , C′′, R, X 1 , J 1 , J 2 , J′ 1 , M, S, S 2 , W 1 , W 2 |
|
Швицкая |
G 3 , P′ 2 |
A 2 , Z′, B 2 , I′′, O x , P 1 , P 2 , T 2 , Q, D′, E′ 2 , I′ 2 , B′′, B′, R, J 1 , J 2 , J′ 1 , M, S, S 2 , H′′, U′′, W 1 , W 2 |
|
Симментальская |
T 2 |
A 2 , Z′, B 2 , I′′, G 1 , G 3 , K′, I 2 , O 2 , O 3 , O x , P 1 , P 2 , T 1 , Y 1 , A′ 2 , E′ 2 , E′ 3 , I′ 2 , P′ 2 , B′′, G′′, R, R 1 , X 1 , J, J 1 , J 2 , J′ 1 , M, S, S 2 , W 1 , W 2 |
|
Красно-пёстрая голштинская |
Не обнаружены |
A 2 , Z′, B 1 , G 1 , G 2 , G 3 , K, I 1 , I 2 , O 3 , O x , P 1 , P 2 , Q, T 1 , T 2 , Y 1 , E′ 2 , I′ 2 , P′ 2 , B′′, G′′, C 2 , C′′, R, R 1 , V, J 1 , J 2 , L′, M, S, S 2 , U, U′, U′′, W 1 , W 2 |
|
Джерсейская |
O x , J 1 |
J 2 , E′ 1 , R 2 , X 2 (нет только у джерсеев); A 2 , Z′, G 1 , G 2 , G 3 , K, I 1 , I 2 , O 4 , P 1 , P 2 , Q, T 1 , T 2 , Y 1 , E′ 2 , G′, I′ 2 , P′ 2 , Y′, B′′, C′′, R, J, J′ 1 , J′ 2 , L′, M, S 2 , U, U′, H′′, U′′, W 2 |
|
Холмогорская |
Не обнаружены |
S1 (нет только у холмогоров); A2, Z′, B1, I′′, G1, G3, K, I1, I2, O3, O x , P 1 , P 2 , T 1 , T 2 , Y 1 , D′, E′ 2 , I′ 2 , P′ 2 , B′′, C 2 , R, R 1 , F′ 2 , J 1 , J 2 , J′ 2 , L′, S, S 2 , U, H′′, U′′, W 1 , W 2 |
Таблица 2 – Иммуногенетическое сходство быков-производителей молочных и комбинированных пород
|
Породы |
Сходство по аллелям основных локусов, r |
Число общих антигенов |
Корреляция, ( rAB ) |
Общее сходство, r i |
||||||||
|
EAA |
EAB |
EAC |
EAJ |
EAL |
EAF |
EAS |
EAM |
EAZ |
||||
|
ЧПГ-А |
0,052 |
0,225 |
0,183 |
0,023 |
0,035 |
0,159 |
0,075 |
0,004 |
0,012 |
33 |
0,814 |
0,851 |
|
ЧПГ-ЧП |
0,029 |
0,226 |
0,109 |
0,010 |
0,028 |
0,142 |
0,096 |
0 |
0,035 |
36 |
0,878 |
0,902 |
|
ЧПГ-Ш |
0,022 |
0,319 |
0,133 |
0 |
0,036 |
0,128 |
0,082 |
0 |
0,024 |
41 |
0,714 |
0,815 |
|
ЧПГ-С |
0,039 |
0,261 |
0,095 |
0 |
0,003 |
0,153 |
0,114 |
0 |
0,025 |
29 |
0,640 |
0,753 |
|
ЧПГ-КПГ |
0,041 |
0,315 |
0,125 |
0,032 |
0,031 |
0,130 |
0,079 |
0 |
0,023 |
32 |
0,870 |
0,901 |
|
А-ЧП |
0,036 |
0,154 |
0,142 |
0,013 |
0,029 |
0,160 |
0,082 |
0 |
0,017 |
21 |
0,734 |
0,788 |
|
А-Ш |
0,027 |
0,182 |
0,178 |
0 |
0,012 |
0,138 |
0,073 |
0 |
0,011 |
24 |
0,590 |
0,689 |
|
А-С |
0,048 |
0,154 |
0,151 |
0 |
0 |
0,166 |
0,077 |
0 |
0,012 |
19 |
0,603 |
0,694 |
|
А-КПГ |
0,052 |
0,233 |
0,162 |
0,039 |
0,035 |
0,158 |
0,078 |
0 |
0,011 |
25 |
0,851 |
0,882 |
|
ЧП-Ш |
0,015 |
0,288 |
0,099 |
0 |
0,015 |
0,124 |
0,085 |
0 |
0,033 |
28 |
0,705 |
0,777 |
|
ЧП-С |
0,028 |
0,214 |
0,052 |
0 |
0,010 |
0,149 |
0,113 |
0 |
0,035 |
23 |
0,669 |
0,749 |
|
ЧП-КПГ |
0,030 |
0,220 |
0,098 |
0,018 |
0,026 |
0,136 |
0,084 |
0 |
0,032 |
23 |
0,766 |
0,818 |
|
Ш-С |
0,022 |
0,238 |
0,079 |
0 |
0,010 |
0,134 |
0,104 |
0 |
0,024 |
26 |
0,638 |
0,756 |
|
Ш-КПГ |
0,022 |
0,227 |
0,116 |
0,006 |
0,011 |
0,110 |
0,071 |
0 |
0,022 |
25 |
0,572 |
0,692 |
|
С-КПГ |
0,039 |
0,207 |
0,050 |
0 |
0 |
0,134 |
0,086 |
0 |
0,023 |
20 |
0,553 |
0,670 |
|
Д-КПГ |
0,055 |
0,248 |
0,076 |
0 |
0,030 |
0,092 |
0,080 |
0 |
0,030 |
23 |
0,402 |
0,570 |
|
Д-С |
0,052 |
0,192 |
0,097 |
0 |
0 |
0,141 |
0,083 |
0 |
0,033 |
18 |
0,484 |
0,650 |
|
Д-Ш |
0,029 |
0,215 |
0,146 |
0 |
0,010 |
0,121 |
0,078 |
0 |
0,031 |
25 |
0,446 |
0,645 |
|
Д-ЧП |
0,039 |
0,146 |
0,100 |
0 |
0,025 |
0,122 |
0,082 |
0 |
0,046 |
19 |
0,459 |
0,601 |
|
Д-А |
0,067 |
0,161 |
0,133 |
0 |
0,034 |
0,131 |
0,085 |
0 |
0,016 |
19 |
0,416 |
0,561 |
|
Д-ЧПГ |
0,055 |
0,247 |
0,119 |
0,012 |
0,029 |
0,133 |
0,086 |
0 |
0,033 |
31 |
0,518 |
0,683 |
|
Д-Х |
0,064 |
0,211 |
0,101 |
0 |
0,027 |
0,128 |
0,059 |
0 |
0,033 |
21 |
0,451 |
0,636 |
|
Х-КПГ |
0,049 |
0,280 |
0,149 |
0,037 |
0,028 |
0,119 |
0,055 |
0 |
0,022 |
27 |
0,756 |
0,820 |
|
Х-С |
0,046 |
0,267 |
0,148 |
0 |
0 |
0,144 |
0,063 |
0 |
0,024 |
21 |
0,647 |
0,757 |
|
Х-Ш |
0,025 |
0,307 |
0,171 |
0 |
0,010 |
0,121 |
0,058 |
0 |
0,023 |
29 |
0,688 |
0,796 |
|
Х-ЧП |
0,035 |
0,267 |
0,141 |
0,016 |
0,023 |
0,133 |
0,066 |
0 |
0,035 |
24 |
0,802 |
0,847 |
|
Х-А |
0,060 |
0,200 |
0,219 |
0,027 |
0,031 |
0,149 |
0,062 |
0,010 |
0,012 |
26 |
0,770 |
0,819 |
|
Х-ЧПГ |
0,048 |
0,333 |
0,170 |
0,021 |
0,027 |
0,138 |
0,056 |
0,006 |
0,025 |
35 |
0,870 |
0,913 |
Примечание: ЧПГ - чёрно-пёстрая голштинская, А - айрширская, ЧП - чёрно-пёстрая, Ш - швицкая, С – симментальская, КПГ – красно-пёстрая голштинская, Д – джерсейская, Х – холмогорская порода.
По EAF-системе было обнаружено высокое иммуногенетическое сходство айрширской породы с симментальской, чёрно-пёстрой и чёрно-пёстрой голштинской – 0,166, 0,160 и 0,159, соответственно. Низкое сходство было получено между джерсейским и красно-пёстрым голштинским скотом – 0,092. По остальным сочетаниям значительных различий не наблюдалось.
В системе EAS большинство сочетаний значительно не отличалось между собой (0,06-0,08), в то время как симментальские быки показали наивысшее сходство с чёрно-пёстрым и чёрно-пёстрым голштинским скотом – 0,113-0,114. Также выделялось сравнение симментальских и швицких, чёрно-пёстрых и чёрно-пёстрых голштинских быков – 0,104 и 0,096, соответственно. Наименьшее сходство было у холмогорского скота с голштинами (0,055-0,056).
Несмотря на отсутствие сходства в 89,3% сочетаний по EAM-системе, изучение локуса EAZ открыло некоторые закономерности в иммуногенетическом сходстве пород. Так, чёрнопёстрый скот имел высокое сходство с джерсейским, симментальским и чёрно-пёстрым голштинским – 0,046, 0,035 и 0,035, соответственно. Айрширский скот имел меньшее сходство со всеми остальными породами (0,011-0,017).
Общее иммуногенетическое сходство было максимальным между холмогорскими и чёрнопёстрыми голштинскими быками (0,913) в связи со скрещиванием этих пород, между чёрно-пёстрыми голштинскими и чёрно-пёстрыми быками (0,902), а также между голштинами разной масти (0,901) по причине родственного происхождения. Невысокое сходство было обнаружено у джерсейских быков с быками остальных пород (0,561-0,683), что закономерно, поскольку долгое время эту породу разводили в изолированном состоянии на острове, и что также отразилось на наличии у неё 4 специфических антигенов. При этом в наших исследованиях высокую степень совпадения показала корреляция ( r AB ) между концентрацией антигенов у пород и способом определения общего иммуногенетического сходства по методике Майяла и Лингстрема (98,6%). Это позволяет предложить более простой и надежный способ определения иммуногенетического сходства по формуле: г , = гав+0,05, где r i - общее иммуногенетическое сходство, rAB – корреляция между частотами встречаемости антигенов у сравниваемых пород, 0,05 – поправочный коэффициент для более точного вычисления. Кроме того, между остальными методами было получено совпадение 80-85%.
Фактор " антиген " э Фактор " порода "
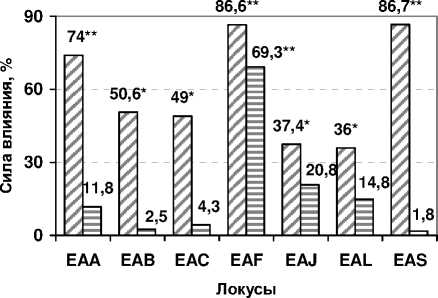
Рис. 3. Сила влияния факторов на разнообразие быков-производителей по локусам групп крови (h2 A , %), * – α<0,05, **– α<0,01 (по Фишеру)
Из рисунка 3 следует, что фактор «антиген» оказывает достоверное влияние на разнообразие быков-производителей исследованных пород по большинству простых и сложных локусов (36,086,7%), в то время как фактор «порода» не оказывает существенного влияния на разнообразие по локусам, за исключением системы EAF (69,3%). Полученные данные подтверждают необходимость тестирования быков-производителей по всем эритроцитарным антигенам.
Нами также получены данные, в которых маркерная способность эритроцитарных антигенов возрастает с увеличением частоты встречаемости от 0,02 до 0,04. При этом наибольшим удоем обладали дочери чёрно-пёстрых голштинских быков-производителей при частоте встречаемости антигенов больше 0,03 – 5194 кг молока, наблюдалось увеличение удоев у коров с частотой встречаемости антигенов ниже 0,01 – 4601 кг молока по первой лактации. Это, по-видимому, объяснимо тем, что в популяции чёрно-пёстрого голштинизированного скота Московской области в настоящее время идёт активное внедрение желательных эритроцитарных антигенов, маркирующих высокую молочную продуктивность.
Кроме того, Н. Г. Букаров c соавторами приводит данные, в которых сказано, что к 1990-му году в селекции чёрно-пёстрых голштинов вводился определённый ряд аллелей [3]. В наших исследованиях групп крови быков «ЦСИО», родившихся в этот период, выявлена высокая концентрация большинства антигенов, составляющих вводимые аллели. Три антигена, из пяти рекомендуемых М. М. Боевым [2] в качестве маркеров долголетия (B′, I′, Q′) у симментальских быков «ЦСИО», имеют среднюю концентрацию (0,024-0,049), что открывает перспективу для их ввода в палево-пёстрые стада. У чёрно-пёстрых быков «ЦСИО» лишь один антиген (Е) имеет высокую концентрацию в числе тех, которые А.А. Коровушкин приводит как нежелательные [9]. Также подтверждаются результаты исследований других учёных.
Из таблицы 3 следует, что у 112 быков-производителей, семя которых в 2004 году реализовывало ОАО «ЦСИО», 58,0% голов имели генотип АА, 30,3 – АВ, 11,6% – генотип ВВ. При этом у быков швицкой, джерсейской и костромской пород была наибольшая частота генотипа ВВ – 38,1, 50,0 и 33,3%, соответственно. У чёрно-пёстрых голштинов лишь 2,1% быков имели генотип ВВ, а большинство пород вообще не отличалось предрасположенностью к высокой сыропригодности молока. Красно-пёстрые голштинские быки не имели даже генотипа с одним геном В, что, очевидно, может затруднить селекцию на сыропригодность молока у красно-пёстрого поголовья России. Между тем большинство пород характеризовались предрасположенностью к повышению качества молока при правильно организованном подборе в стадах. При этом в 2008 году, как и в 2004, наибольшей концентрацией гетерозиготных генотипов у чёрно-пёстрых голштинских быков отличалась линия Уес Идеал. Частота генотипа ВВ к 2008 году у этой породы повысилась до 7,2%. Наибольшая частота гена В была получена у швицких быков – 0,55.
Таблица 3 – Частота генотипов у быков разных пород ОАО «ЦСИО»
|
Порода |
n |
Частота, % |
||
|
κ-Cas AA |
κ-Cas AB |
κ-Cas BB |
||
|
Чёрно-пёстрая голштинская |
47 |
59,7 |
38,2 |
2,1 |
|
Красно-пёстрая голштинская |
7 |
100 |
0 |
0 |
|
Швицкая |
21 |
28,6 |
33,3 |
38,1 |
|
Джерсейская |
6 |
33,3 |
16,7 |
50,0 |
|
Холмогорская |
8 |
62,5 |
37,5 |
0 |
|
Айрширская |
16 |
81,5 |
18,5 |
0 |
|
Симментальская |
4 |
50,0 |
50,0 |
0 |
|
Костромская |
3 |
66,6 |
0 |
33,3 |
|
Всего |
112 |
58,0 |
30,3 |
11,6 |
Следует отметить, что в родословных чёрнопёстрых голштинских быков разных линий с изменением АА→АВ, у материнских предков прослеживалась тенденция к уменьшению жирности молока на 0,01-0,25%, однако при этом у коров линии Р. Соверинг намечалось увеличение удоев по 1 лактации – до 12340 кг. У матерей и матерей матерей в линии У. Идеал при изменении АА→АВ→ВВ удой понизился с 9694 до 9108 и с 9636 до 6448 кг. Меньшим потенциалом молочной продуктивности отличались коровы линии С. Т. Рокит. Аналогичные тенденции наблюдались по селекционным признакам молочной продуктивности у материнских предков быков-производителей за наивысшие лактации, а следовательно, происходило понижение генетического потенциала быков.
Выводы
Таким образом, быки-производители разных пород могут иметь как высокие, так и низкие индексы общего иммуногенетического сходства, что сопровождается наличием специфических антигенов или отсутствием целого ряда антигенов, характерных для других пород. По аллелям простых и сложных локусов, как правило, прослеживается значительное отличие сходства в большинстве сравнений. Наиболее информативны при изучении аллельного состава системы EAB, EAC и EAS, определённый интерес в маркерной селекции представляют системы EAA, EAF и EAZ, отдельные локусы между тем не представлены у исследуемых пород ни одним антигеном. В связи с разработкой новых компьютерных программ существует возможность создания современных способов определения генетического сходства и моделирования популяционных изменений в аллельном и кластерном составе по большинству пород, разводимых в России. С целью оптимизации селекционного процесса, внедрения метода BLUP и матричной алгебры в скотоводство приобретает значение определение силы влияния различных факторов на аллельный состав в локусах и маркерную способность антигенов. Однако при изучении сходства между линиями в породе методы маркерной селекции с использованием эритроцитарных антигенов не всегда информативны, что требует дополнительного генетического анализа.
Быки-производители большинства пород могут быть пригодны для увеличения концентрации гена В при правильно организованном подборе в популяциях крупного рогатого скота России, однако при этом следует вести одновременный отбор на сохранение высоких удоев. Целесообразно оценивать генетические корреляции между удоями и процентным содержанием основных компонентов молока, таких, как жир, белок и казеин. Необходимо отбирать при закреплении к стаду семя быка, который не только принесёт в дойное стадо ген В, но и будет способствовать одновременному увеличению основных признаков молочной продуктивности.
В целом, использование групп крови и ДНК-анализа в племенной работе повышает селекционную культуру, способствует раннему прогнозу племенной ценности животных и перспектив их дальнейшего использования, а внедрение в производство оптимизированных программ селекции позволяет во многих хозяйствах России за короткий срок повысить удой до 6000-8000 кг молока в среднем на корову.
Список литературы Генетические аспекты модернизации молочного скотоводства
- Бакай, А.В. Генетика/А.В. Бакай, И.И. Кочиш, Г.Г. Скрипниченко. -М.: КолосС, 2007. -448 с.
- Боев, М. М. Селекционно-генетические аспекты продления сроков хозяйственного использования крупного рогатого скота/М.М. Боев, А.О. Савин, М.М. Боев//Материалы международной научно-практической конференции 27-28 марта 2008 года «Трансферт инновационных технологий в животноводстве». -Орёл, издательство ОГАУ, 2008. -С. 27-30.
- Букаров, Н. Г. Генетика на службе молочного скотоводства / Н.Г. Букаров, И.М. Морозов // // Материалы международной научно-практической конференции 27-28 марта 2008 года «Трансферт инновационных технологий в животноводстве». - Орёл, издательство ОГАУ, 2008. - С. 30-33.
- Букаров, Н. Новый уровень познания маркерных генов групп крови у скота/Н.Букаров, Е.Лебедев, И.Морозов//Молочное и мясное скотоводство.-2005. -№7. -С. 39-41.
- Лакин, Г. Ф. Биометрия. -М.: Высшая школа, 1990. -352 стр.
- Исламова, С. Порода и антигенный состав крови быков-производителей/С. Исламова, Ф. Исламов//Молочное и мясное скотоводство.-2006. -№5. -С. 34-35.
- Исламова, С. Применение ДНК-технологии в селекции/С. Исламова, Ф. Исламов, И. Долматова, Р. Биккинин //Молочное и мясное скотоводство.-2007. -№5. -С. 2-4.
- Коновалова, Е. Н. Гены белков молока и микросателлитные профили в популяциях симментальского скота разного происхождения / Е. Н. Коновалова, О. А. Львина, В. И. Сельцов и др. // // Сельскохозяйственная биология. - 2007. - №:6. - С. 35-40.
- Коровушкин, А.А. Иммуногенетические маркеры устойчивости коров к заболеваниям//Зоотехния. -2004. -№6. -С. 9-11.
- Проскурина, Н. В. Сравнительный анализ информативности эритроцитарных антигенов и ДНК-микросателлтов как генетических маркеров в селекционно-племенной работе со свиньями канадской селекции/Н.В. Проскурина, Т.И. Тихомирова, Е.А. Гладырь и др.//Сельскохозяйственная биология. -2007. -№:6. -С. 41-46.
- Терлецкий, В.П. Оценка племенных животных по полиморфизму генов и ДНК/В.П. Терлецкий, Н.В. Дементьева, Е. С. Усенбеков//Зоотехния. -2001.-№1.-С. 14-16.
- Тинаев, А. Племенные ресурсы быков-производителей чёрно-пёстрой породы/А.Тинаев, Л. Калашникова, Т. Ганченкова//Молочное и мясное скотоводство.-2009. -№1. -С. 5-7.
- Тинаев, А. Хозяйственно-полезные признаки чёрно-пёстрого скота с разными генотипами каппа-казеина/А. Тинаев, Л. Калашникова, К. Аджибеков//Молочное и мясное скотоводство.-2005. -№5. -С. 30-32.
- Хайруллина, Н. Влияние сочетания локусов эритроцитарных антигенов быков-производителей на структуру стада крупного рогатого скота/Н. Хайруллина, Н. Фенченко, З. Ярмухаметова, Р. Ахмадуллин//Зоотехния. -2007. -№6. -С. 5-6.
- Черных, А. Генотип каппа-казеина и качество молока чёрно-пёстрых коров/А.Черных, Л. Калашникова//Молочное и мясное скотоводство.-2008. -№5. -С. 9-10.
- Эрнст, Л.К. Изучение влияния прилития крови голштинского скота на изменение генофонда крупного рогатого скота отечественных пород с использованием ДНК-микросателлитов/Л. К. Эрнст, Н. А. Зиновьева, Е. Н. Коновалова и др.//Зоотехния. -2007. -№12. -С. 2-5.
