Генетические дистанции как показатель микроэволюционных процессов при формировании енисейского типа красно-пестрого скота

Автор: Еремина Ирина Юрьевна
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Ветеринария и зоотехния
Статья в выпуске: 11, 2020 года.
Бесплатный доступ
Цель исследования - проанализировать микроэволюционные процессы при формировании енисейского типа красно-пестрого скота через структурную динамику генетических расстояний. Задачи исследования: 1) проанализировать динамику аллельного состава в простых системах групп крови (J, L, M, Z); 2) рассчитать генетические расстояния, провести кластерный анализ; 3) продемонстрировать направленность микроэволюционных процессов с помощью дендрограмм и многомерного шкалирования. Методы исследования: иммуногенетический, зоотехнический, методы математического моделирования, математической статистики, аппарата многомерного шкалирования (пакет STATISTICA). Объект исследования: стада животных красно-пестрой породы. Мониторинг генетической структуры, уровня полиморфности типов красно-пестрого скота проводили за десятилетний период по пяти стадам и в целом по енисейскому типу. Общее обследованное поголовье по типу - 1996 голов. Анализировали показатели: частота антигенных фактов и аллелей простых систем, индекс генетического (антигенного) сходства r. Внутристадная динамика оценивалась по периодам, один из них - базовое стадо (исходные данные), а другой - уже измененное в ходе селекции стадо с интервалом рождения животных через 2-3 года, условно обозначенные как период 1, 2 и 3. Установлено, что простые системы, имеющие ограниченное число аллелей, фиксируют изменения, протекающие под воздействием искусственного отбора. Кумулятивный эффект двух факторов, отражающих воздействие голштинского генофонда на изменчивость частотных характеристик антигенов групп крови, составил 49,5 %. Генетические дистанции отражают микроэволюционные процессы на разных уровнях биологической организации, происходящие при изменении генофонда местного скота под воздействием голштинских производителей в процессе формирования енисейского типа красно-пестрого скота. Следует шире использовать сложные математические методы применительно к сельскохозяйственным объектам, как на уровне породы в целом, так и к меньшим таксономическим популяционным единицам.
Селекция, полиморфизм, популяция, генетические расстояния, енисейский тип, молочный скот, красно-пестрая порода коров, иммуногенетические маркеры
Короткий адрес: https://sciup.org/140250551
IDR: 140250551 | УДК: 575.174.015.3: | DOI: 10.36718/1819-4036-2020-11-175-183
Genetic distances as the indicator of microevolutionary processes in the formation of Yenisei type of red-motley cattle
The purpose of the study was to analyze microevolutionary processes in the formation of the Yenisei type of red-motley livestock through structural dynamics of genetic distances. The research tasks were: 1) to analyze the dynamics of allelic composition in simple blood group systems (J, L, M, Z); 2) to calculate genetic distances, conduct cluster analysis; 3) to demonstrate the orientation of microevolution processes using dendrograms and multivariate scaling. The research methods were immunogenetic, zootechnical, methods of mathematical modeling, mathematical statistics, multivariate scaling apparatus (STATISTICA package). The objects of the study were the herds of red-motley breed. Monitoring genetic structure, the level of polymorphism of the types of red-motley cattle was carried out over the period of 2005-2015 for five herds and in general for the Yenisei type. Total number of the surveyed was 1996 heads of the type. The following indicators were analyzed: the frequency of antigenic facts and alleles of simple systems, genetic (antigenic) similarity index r. Intra-herd dynamics was estimated by the periods, one of them was basic herd (initial data) and the other was the herd already changed during selection with an interval of birth of animals after 2-3 years, conditionally designated as the period 1, 2 and 3. It was found that in simple systems having a limited number of alleles, the changes occurring under the influence of artificial selection had been recorded. The cumulative effect of two factors reflecting the effect of the Holstein gene pool on the variability of the frequency characteristics of blood group antigens made 49.5 %. Thus, genetic distances reflect microevolutionary processes at different levels of biological organization occurring when the gene pool of local livestock changes under the influence of Holstein producers in the process of forming the Yenisei type of redmotley cattle. More sophisticated mathematical methods should be used for agricultural facilities, both at the breed level as a whole and for smaller taxonomic population units.
Текст научной статьи Генетические дистанции как показатель микроэволюционных процессов при формировании енисейского типа красно-пестрого скота
Введение . Внутрипородная селекция, имея постоянный и кумулятивный характер, является довольно медленной стратегией генетического улучшения. В связи с чем дает заинтересованным структурам время для адаптации системы производства к вводимым изменениям. Енисейский тип красно-пестрой породы признан новым селекционным достижением, на которое выдан патент 22 июня 2009 г. за № 4804, улучшателем для животных, отвечающим промышленной технологии и рекомендуемым для широкого использования в Сибирском регионе [1]. Работа по его выведению, начатая с 1999 г. в базовых хозяйствах, принесла закономерные плоды. Фенотипический прирост по удою у коров с 2008 по 2017 г. составил 1 119 кг, при содержании массовой доли жира в молоке – 0,08 %, белка – 0,01 %. Реализованный прирост генетического потенциала, рассчитанный по методике S. Smith в модификации В.М. Кузнецова, за период формирования типа составил +86 %. При этом по отдельным хозяйствам этот показатель отрицательный. В целом у молочного крупного рогатого скота показатели продуктивности, много лет служившие основной целью селекции, за прошедшие десятилетия во всем мире значительно увеличились. По данным информационного центра ВНИИплем, молочная продуктивность в России за период с 2010 по 2017 г. возросла на 37,3 %. В то же время поголовье коров в целом по
России снизилось на 28,9 %, а в Сибирском федеральном округе – на 20,25 % [2].
Известно, что отбор исключительно по выходу продукции приводит к ухудшению здоровья и репродуктивных функций, увеличивает метаболическое напряжение и уменьшает продолжительность хозяйственного использования [3].
Улучшение отечественного скота с использованием голштинской породы сужает генофонд молочного скота России, снижая генетическое разнообразие и число ценных, редких пород животных, не выгодных с экономической точки зрения. Ориентация на выдающихся производителей уменьшает их число, ускоряет темпы повышения гомозиготности маточных стад. А без необходимого генетического разнообразия популяции животных теряют свою эволюционную приспособленность и становятся неустойчивыми к воздействию патогенной микрофлоры и негативным влияниям окружающей среды [4, 5].
Потеря генетического разнообразия животных приводит к значительному экономическому ущербу [6, 7]. Еще в 2004 г. академиком Ю.П. Алтуховым постулированы три категории аргументов в пользу сохранения генофондов локальных пород: культурные, научные и экономико-биологические [8]. Вместе с тем экономически ценные свойства животноводческой продукции модифицируются под воздействием измен- чивых, периодически трансформирующихся нужд и ожиданий потребителей.
Все это указывает на потребность в высокой генетической изменчивости, потеряв которую, можно достичь так называемого «селекционного плато», при котором деятельность селекционеров становится малоэффективной [9, 10].
Применяемые методы мониторинга не могут в достаточной степени покрыть имеющиеся потребности контроля внутрипопуляционных изменений, протекающих под воздействием селекционного давления. Установлено, что у близких по происхождению пород больше сходных аллелей с большей частотой встречаемости. При сравнении генетически близких популяций (пород) нельзя пренебрегать влиянием имеющегося полиморфизма. Для точности анализа потребуются либо данные по большому числу белков, либо генов, последнее весьма трудоемко и дорогостояще [11].
Уже не первое десятилетие метод оценки генетической структуры популяций животных дополняется анализом особенностей исследуемых популяций по полиморфным белкам и группам крови животных. Белковый полиморфизм и эритроцитарные антигены стали первыми молекулярными маркерами, использованными в исследованиях домашних животных [12, 13].
Цель исследования: проанализировать мик-роэволюционные процессы при формировании енисейского типа красно-пестрого скота через структурную динамику генетических расстояний.
Задачи исследования:
-
1) проанализировать динамику аллельного состава в простых системах групп крови (J, L, M, Z);
-
2) рассчитать генетические расстояния, провести кластерный анализ;
-
3) продемонстрировать направленность мик-роэволюционных процессов с помощью дендрограмм и многомерного шкалирования.
Методы исследования: иммуногенетиче-ский – для определения антигенов групп крови, зоотехнический – для формирования базы данных Селэкс, а также методы математического моделирования, основные положения математической статистики, методов оптимизации, аппарата многомерного шкалирования (пакет STATISTICA). Внедрение компьютерных технологий в исследования, направленные на анализ селекционных воздействий, позволило в на- стоящее время использовать сложные математические методы применительно к сельскохозяйственным объектам как на уровне породы в целом, так и к меньшим таксономическим популяционным единицам [14].
Объект исследования: стада животных красно-пестрой породы (АО «Краснотуранский», АО «Тубинск», ЗАО «Назаровское», АО «Сол-гон», ОАО «Племзавод “Красный Маяк”»). Мониторинг генетической структуры, уровня поли-морфности типов красно-пестрого скота проводили за период 2005–2015 гг. по отдельным стадам и в целом по енисейскому типу. Проводились сравнения данных по популяции за 2 или 3 периода: базовое стадо (исходные данные) и уже измененное в ходе селекции стадо с интервалом рождения животных через 2 и 3 года, которые условно обозначили как период 1, 2 и 3 (табл. 1). Опираясь на теоретические механизмы изменения генофонда при скрещивании с голштинскими производителями, предполагаем, что популяция периода 1 по сравнению с периодом 2 будет иметь некоторые, возможно, несущественные отличия и значительные изменения по отношению к периоду 3. При этом характеристики популяции периодов 2 и 3 будут весьма близки. Общее обследованное поголовье по типу – 1996 гол. Для определения направленности микроэволюционных процессов в анализ дополнительно введены иммуногенети-ческие показатели о животных разных пород предшествующих генераций (1995 год рождения). Анализировали показатели: частота антигенных фактов и аллелей простых систем, индекс генетического (антигенного) сходства r [15].
Результаты исследования и их обсуждение. Изменения аллельного состава изучали в популяционных группах – стадах четырех хо-зяйств-оригинаторов по енисейскому типу и в стаде ОАО «Племзавод “Красный Маяк”». Проанализирована динамика концентрации аллелей в четырех системах групп крови J, L, M, Z. Учитывая, что данные системы простые и содержат ограниченное число аллелей (2–3), ожидаемая изменчивость частот меньше, в отличии от традиционно анализируемой полиаллельной «В»-системы.
«J»-система сбалансирована во всех популяциях, разводящих животных енисейского типа красно-пестрого скота (Тубинский, Назаровский, Солгонский). В то время как в популяции краснопестрого голштинского скота ОАО «Племзавод “Красный Маяк”» это отношение смещено в сторону негативного аллеля j. По периодам отмечается: существенное возрастание частоты в ОАО «Племзавод Красный Маяк» (+0,078), незначительные изменения в АО «Тубинск» (–0,10),
АО «Солгон» (–0,07) и ЗАО «Назаровское» (0,08) при нестабильности частоты и разнонаправлен-ности процесса в АО «Краснотуранский» (+0,20; –0,13; –0,05).
«L»-система относительно стабильна, исключение – АО «Краснотуранский», где наблюдается увеличение частоты с 0,17 до 0,66 (табл.).
Частота аллелей простых систем у молочных коров маточного поголовья Красноярского края
|
Популяция, период |
Год рождения коров |
Обследовано, гол. |
Частота |
|||
|
J |
L |
M |
Z |
|||
|
АО «Тубинск», 1 |
2008 |
401 |
0,423 |
0,313 |
0,080 |
0,318 |
|
АО «Тубинск», 2 |
2010 |
258 |
0,319 |
0,448 |
0,093 |
0,518 |
|
АО «Краснотуранский», 1 |
2005 |
119 |
0,398 |
0,170 |
0,051 |
0,348 |
|
АО «Краснотуранский», 2 |
2008 |
437 |
0,414 |
0,460 |
0,092 |
0,556 |
|
АО «Краснотуранский», 3 |
2010 |
220 |
0,186 |
0,500 |
0,145 |
0,423 |
|
АО «Краснотуранский», 4 |
2011 |
132 |
0,094 |
0,656 |
0,156 |
0,375 |
|
АО «Солгон», 1 |
2008 |
272 |
0,491 |
0,303 |
0,033 |
0,432 |
|
АО «Солгон», 2 |
2010 |
265 |
0,436 |
0,364 |
0,080 |
0,246 |
|
ЗАО «Назаровское», 1 |
2008 |
359 |
0,520 |
0,254 |
0,039 |
0,388 |
|
ЗАО «Назаровское», 2 |
2010 |
70 |
0,600 |
0,286 |
0,157 |
0,586 |
|
ОАО «Племзавод “Красный Маяк”», 1 |
2008 |
175 |
0,057 |
0,303 |
0,120 |
0,503 |
|
ОАО «Племзавод “Красный Маяк”», 2 |
2012 |
422 |
0,014 |
0,690 |
0,075 |
0,332 |
|
ОАО «Племзавод “Красный Маяк”», 3 |
2013 |
200 |
0,135 |
0,349 |
0,146 |
0,495 |
«M»-система имеет невысокие частоты встречаемости у всех молочных животных при общей тенденции незначительного повышения частот.
«Z»-система более сбалансирована. В динамике прослеживаются групповые особенности изменения насыщенности, происходящие за счет генофонда используемых быков-производителей).
В целом изменения частот аллелей простых систем указывают на микроэволюционные изменения, протекающие под воздействием искусственного отбора.
Изменения в генофонде отражает индекс генетического сходства, рассчитанный с учетом всех иммуногенетических систем. Графическое представление соотношения индексов в виде кластеров отражает рисунок 1. На дендрограмме А помеси F1 (KrM F1), в генофонде которых присутствовала симментальская порода, находятся на наибольшем генетическом расстоянии от анализируемого кластера енисейского типа. Наименьшее расстояние – между животными 2 и 3 переходных периодов (рис. 1).
В популяционной группе АО «Краснотуран-ский» (дендрограмма Б) картина иная: животные позднего периода (сформированный енисейский тип) значительно отличаются от всего кластера 1, 2 и 3 периода.
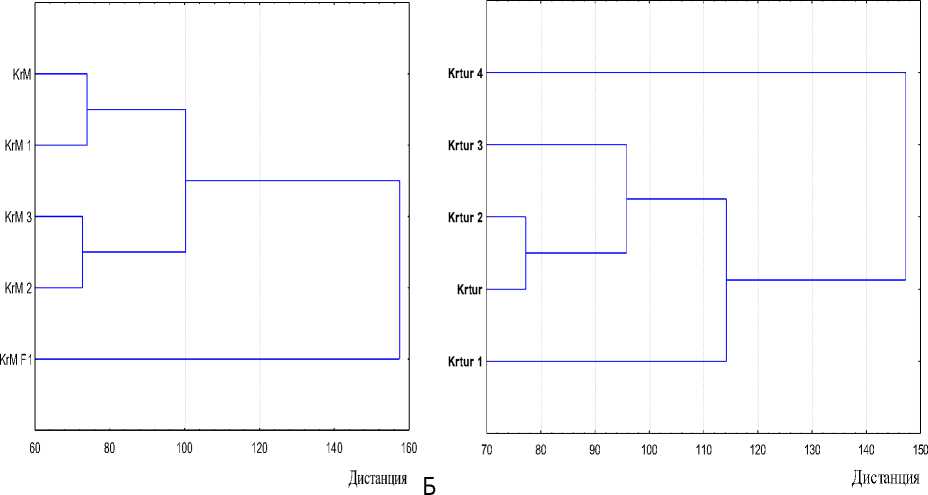
Рис. 1. Сравнительная характеристика процессов преобразования генофонда красно-пестрой породы: А – ОАО «Племзавод “Красный Маяк”» (KrM F 1 , КrМ, КrМ1, КrМ2, КrМ3 , );
Б – АО «Краснотуранский» (Krtur, Krtur1, Krtur2, Krtur3)
Обе дендрограммы указывают на последовательный процесс изменения, происходящий в генофонде, причем на момент анализа в АО «Краснотуранский» этот процесс уже завершен, тип сформирован.
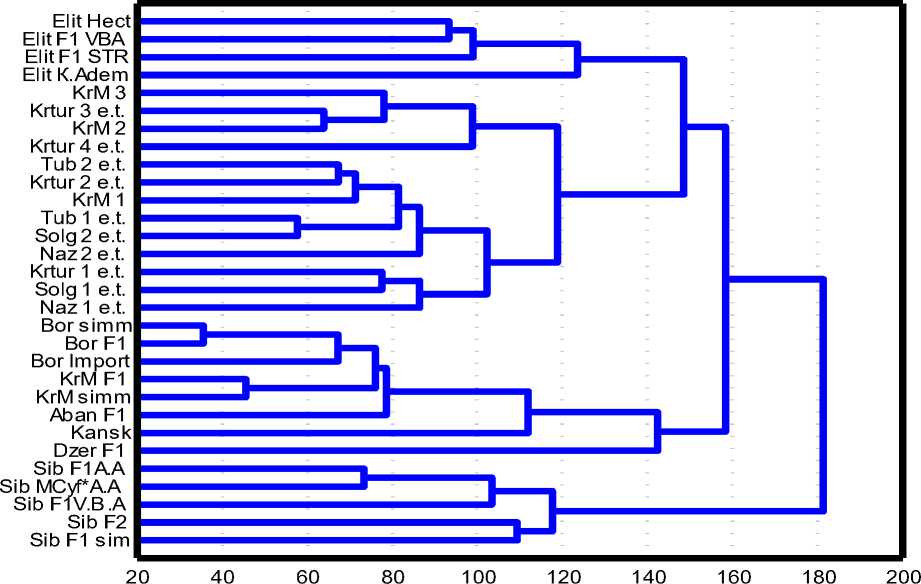
Для того чтобы рассмотреть процессы с уровня межпородных различий, анализировали: исходный симментальский, интродуцируемый красно-пестрый голштинский, гибриды первого поколения и популяции, работавшие по формированию нового енисейского типа краснопестрого скота. Кроме того, с целью определения относительной локализации на дендрограмме и ее масштабирования в анализ включены ряд популяций черно-пестрого корня. Четко виден единый кластер енисейского типа красно-пестрого скота, подтверждающий его генетическую обособленность (рис. 2).
Интересно, что среди кластера енисейского типа присутствует популяционная группа ОАО «Племзавод “Красный Маяк”» 3. Это можно объяснить, во-первых, генетической близостью к енисейскому типу, работавшими быками-производителями, имеющими высокое родство, а во-вторых, недостаточным количеством сравнительных данных по красно-пестрому молочному скоту, не отнесенному к енисейскому типу. Эти хозяйства не являются репродукторами и не проводят иммуногенетический анализ животных.
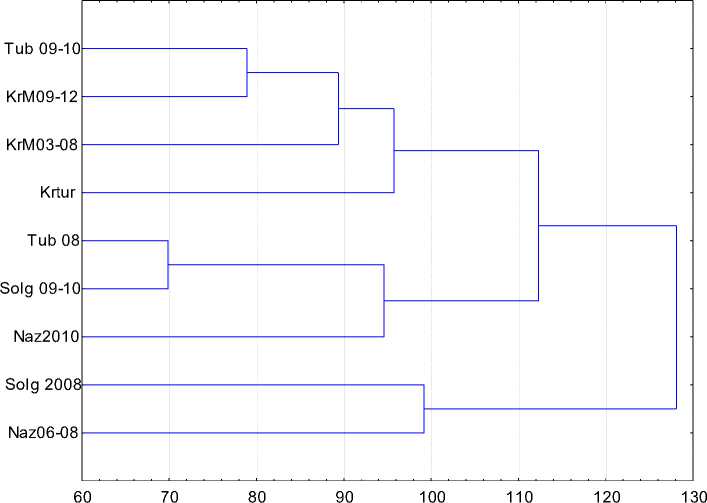
Рассматривая отдельно процесс формирования нового типа по годам (рис. 3, 4), следует отметить происходящие изменения в генофондах соответствующих популяций. На дендрограмме одни и те же популяции, но за разные периоды времени, объединяются в разные кластеры: в 2008 г. наблюдается сходство популяций ЗАО «Назаровское» и АО «Солгон», и через два года это сходство сохраняется, но АО «Сол-гон» по генофонду становится более близким с АО «Тубинск» (2008 г.), т. е. идет как бы вслед, демонстрируя результат от воздействия на маточные стада наследственности голштинских производителей.
Дистанция
Рис. 2. Обобщенная дендрограмма, отражающая межпородные генетические расстояния (e.t. – енисейский тип)
Linkage Distance
Рис. 3. Дендрограмма генетических расстояний, отражающих процесс формирования енисейского типа по годам
Activ e
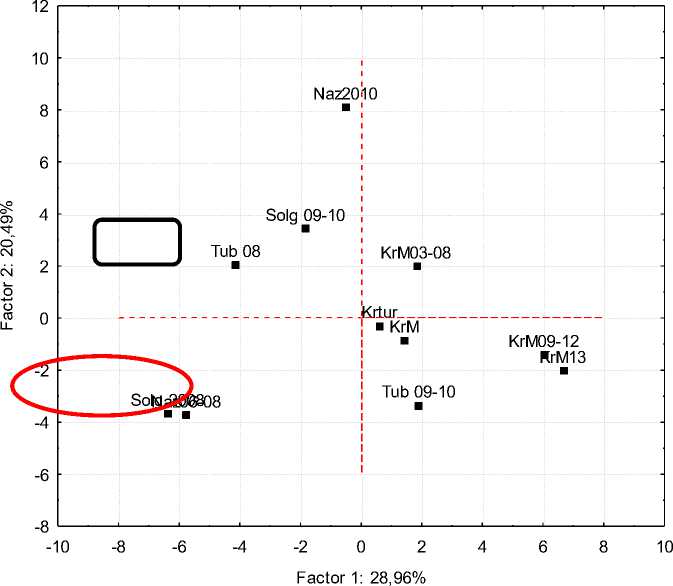
Рис. 4. Проекция многомерного пространства по 54 антигенам на плоскость, отражающая микроэволюционные процессы
Двухмерная проекция иллюстрирует динамику происходящих процессов более объемно. Закономерно предположить, что на рисунке 4 проявилось воздействие фактора 1 – принадлежность (типичность) к породе (в данном случае как принадлежность к новому типу), возрастающее во всех популяциях. В АО «Тубинск» с –4 в 2008 г. до +2 в 2009–2010 гг. В ОАО «Племзавод “Красный Маяк”» с 1,7 в 2003–2008 гг. до 6,78 в 2013 г. В АО «Солгон» та же тенденция: с –6 в 2008 г. до –1,7 в 2009–2010 гг. Общая направленность– увеличение влияния данного фактора (на проекции – слева направо).
По фактору 2 (влияние производителя) ситуация иная: в АО «Солгон» – повышение с –2 в 2008 г. до 3,5 в 2009–2010 гг., а в АО «Тубинск» и ОАО «Племзавод “Красный Маяк”» – снижение доли влияния данного фактора, т. е. динамика разнородна и несистемна. Наложение на проекции популяций коров АО «Солгон» 2008 г. и ЗАО «Назаровское» 2005–2008 гг. указывает на близость их генофондов в тот период, обусловленную влиянием родственных быков. Кумулятивный эффект двух факторов, отражающих воздействие голштинского генофонда на изменчивость частотных характеристик антигенов групп крови, составил 49,5 %.
Выводы. Использование математических методов применительно к сельскохозяйственным объектам позволило выявить особенности микроэволюционных процессов при формировании енисейского типа красно-пестрого скота.
Генетические дистанции отражают микро-эволюционные процессы, происходящие при изменении генофонда местного скота под воздействием голштинских производителей в процессе формирования енисейского типа краснопестрого скота. Установлено, что в простых системах, имеющих ограниченное число аллелей, фиксируют изменения, протекающие под воздействием искусственного отбора. На уровне породы и популяционном уровне выявлена экологогенетическая дивергенция, обусловленная локализацией каждой отдельной популяции. Двухмерная проекция демонстрирует влияние двух факторов: принадлежность к породе, енисейскому типу (28,96 %) и влияние генотипов быков, работавших на маточном поголовье (20,49 %).
Список литературы Генетические дистанции как показатель микроэволюционных процессов при формировании енисейского типа красно-пестрого скота
- Голубков А.И., Лефлер Т.Ф. Создание внутрипородного типа "Енисейский" красно-пестрой породы // Вестник КрасГАУ. 2016. № 1. С. 173-180.
- Голубков А.И. Реализация генетического потенциала молочной продуктивности у коров енисейского типа красно-пестрой породы за 2008-2017 годы // Агросибирь. 2018. № 147. С. 20-22.
- Rischkowsky B. et al. Состояние всемирных генетических ресурсов животных в сфере продовольствия и сельского хозяйства. Рим; М., 2010. С. 428.
- Деева В.С., Сухова Н.О. Группы крови крупного рогатого скота и их селекционное значение. Новосибирск, 2002. 172 с
- Тихонов В.Н., Бобович В.Е. Мониторинг микроэволюции и породообразования свиней на основе молекулярно-иммуногенетического анализа // Сельскохозяйственная биология. 2004. № 2. С. 10-27.
- Новиков А.А., Семак М.С. Значение, правила проведения и состояние генетической экспертизы племенной продукции в Российской Федерации // Теоретические и прикладные аспекты современной науки. 2014. № 5-1. С. 199-203.
- Камалдинов Е.В. Генофонд пород крупного рогатого скота и свиней Западной Сибири: автореф. дис.. д-ра биол. наук. Новосибирск, 2013.
- Алтухов Ю.П. Генетические процессы в популяциях. М., 2003. 372 с.
- Улимбашев М.Б., Кулинцев В.В., Селионова М.И. и др. Рациональное использование генофонда ценных пород животных с целью сохранения биологического разнообразия // Юг России: экология, развитие. 2018. № 2.
- Паронян И.А. Возможности сохранения и совершенствования генофонда пород крупного рогатого скота отечественной селекции // Достижения науки и техники АПК. 2018. № 5. С.
- Попов Н.А., Иванов В.А. Пути разведения крупного рогатого скота малочисленных пород с использованием аллелей групп крови. Дубровицы: Изд-во Всерос. ин-т животноводства, 1999. 70 с.
- Misztal I. Challenges of application of marker assisted selection - a review // Anim. Sci. Pap. and Rep., 2006, 24(1):5-10.
- Герасимова Л.А., Еремина И.Ю. Имуногенетический анализ базового генофонда скота в ОАО "Племзавод Красный Маяк" // Вестник КрасГАУ. 2015. № 3. С. 106-110.
- Кузнецов В.М. Методы Нея для анализа генетических различий между популяциями // Проблемы биологии продуктивных животных. 2020. № 1. С. 91-110.
- Животовский Л.А., Машуров А.М. Методические рекомендации по использованию статистического анализа иммуногенетических данных в селекции животных // ВНИИ животноводства. Дубровицы, 1987. 24 с.
