Генетические и паратипические факторы, характеризующие эффективность использования корма у свиней породы Дюрок

Автор: Белоус А.А., Сермягин А.А., Костюнина О.В., Требунских Е.А., Зиновьева Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика племенной ценности
Статья в выпуске: 4 т.53, 2018 года.
Бесплатный доступ
Эффективность использования корма - важнейший экономически значимый показатель в свиноводстве. Для ее оценки используют значения среднесуточного потребления корма (daily feed intake, DFI) и конверсии корма как отношения количества потребленного корма к привесу живой массы за определенный период времени (feed conversion rate, FCR). Одна из групп факторов, способных оказать влияние на показатели эффективности использования корма, - кормовое поведение свиней. В настоящей работе впервые в России получены результаты, характеризующие особенности кормового поведения хрячков породы дюрок во взаимосвязи с показателями эффективности использования корма на автоматических кормовых станциях. Целью нашей работы было изучение влияния генетических и паратипических факторов на эффективность использования корма и кормовое поведение свиней ( Sus scrofa ). Исследования проводили на базе селекционно-генетического центра ООО «СГЦ» (п. Верхняя Хава, Воронежская обл.) с июля 2017 года по март 2018 года...
Свиньи, порода дюрок, конверсия корма, кормовое поведение, живая масса, среднесуточный прирост, наследуемость, изменчивость
Короткий адрес: https://sciup.org/142216571
IDR: 142216571 | УДК: 636.4:636.082:575.1 | DOI: 10.15389/agrobiology.2018.4.712rus
Study of genetic and environmental factors, characterizing the feed efficiency in Duroc pigs
Feed efficiency is the most important economically relevant factor in swine breeding. The values of daily feed intake (DFI) and feed conversion rate (the ratio of feed intake to the body weight gain for a certain period, FCR). A group of factors that can affect the feed efficiency is the feeding behavior. In this regard, it is relevant to study the genetic and environmental variability of a number of factors that affect the growth, feed efficiency, and ethological features of feed intake in Duroc pigs based on the automatic feeding station data records. The aim of our study was to select at test population the factors which can be associated with feed efficiency, including following traits: body weight (BW, kg), average daily gain (ADG, g), daily feed intake (DFI, g/day), time spent at the feeding station (TPD, min), the number of visits to the feeding station per day (NVD, times), feed intake per visit (FPV, g), feed rate (FR, g/min), and the time at the feeding station per visit (TPV, min)...
Текст научной статьи Генетические и паратипические факторы, характеризующие эффективность использования корма у свиней породы Дюрок
Эффективность использования корма — важнейшая составляющая экономической привлекательности производства свинины. Для оценки этого признака используют показатели среднесуточного потребления корма (daily feed intake, DFI) и конверсии корма как отношения потребленного корма к приросту живой массы за определенный период времени (feed conversion rate, FCR). Установлена положительная корреляция ( r от 0,32 до 0,70) между DFI и среднесуточными приростами (average daily gain, ADG) (1). Варьированием DFI может объясняться до 59 % изменчивости ADG (2). Вариабельность показателей эффективности использования корма обусловлена как внешними, так и внутренними факторами. К внешним факторам относятся состав и энергетическая питательность рационов, менеджмент и климатические условия. Внутренние факторы связаны с различными физиологическими процессами (регуляция аппетита, абсорбционная способность кишечника, усвояемость питательных веществ, терморегуляция, мышечная активность и т.д.), а также состоянием анаболического и катаболического обмена веществ (3).
Вместе с тем показатели эффективности использования корма с точки зрения популяционно-генетических исследований интересны в комплексе факторов кормового поведения свиней. Обладая информацией по закономерностям наследования и изменчивости этологических признаков, можно технологическими приемами регулировать функциональные потребности животных, исходя из породоспецифических особенностей. Разработка компьютеризированных систем откорма (кормовые станции, фидлоты) сделала возможным автоматический высокоточный ежесуточный индивидуальный учет не только потребления корма и прироста живой массы, но и признаков кормового поведения (4). Эта информация открывает новые возможности в изучении его особенностей у свиней во взаимосвязи с показателями эффективности использования корма.
Известно, что эффективность использования корма и кормовое поведение неодинаковы у разных пород свиней (1, 5, 6). Если конверсия корма по породам относительно стабильна (отмечается ее поступательное снижение за последние 15 лет, обусловленное интенсивной селекцией по этому показателю и совершенствованием рационов), то значения показателей кормового поведения в различных исследованиях сильно варьируют как между, так и внутри пород (7-9). Установлено, что показатели конверсии корма и кормового поведения характеризуются умеренной и высокой степенью наследуемости (10-12), однако использование последних в селекции требует сведений об их взаимосвязях с признаками эффективности использования корма. Хотя проведенные исследования показали наличие корреляций между вышеназванными показателями, в большинстве случаев выявленные закономерности имели породо- и популяционно-специфический характер (1, 13, 14). Работы, выполненные ранее на российских популяциях свиней, в основном были направлены на выявление связи между конверсией корма и другими хозяйственно полезными признаками без учета их изменчивости (15, 16). Исследования, проведенные на других видах сельскохозяйственных животных, показывают перспективность включения конверсии корма в характеристику мясных и откормочных качеств поголовья (17, 18).
В настоящем исследовании впервые в России получены результаты, характеризующие особенности кормового поведения хрячков породы дюрок во взаимосвязи с показателями эффективности использования корма на автоматических кормовых станциях.
Нашей целью было изучение влияния генетических и паратипиче- ских факторов на эффективность использования корма и кормовое поведение свиней российской репродукции.
Методика. Исследования проводили с июля 2017 года по март 2018 года (селекционно-генетический центр ООО «СГЦ», п. Верхняя Хава, Воронежская обл.). Учеты выполняли с использованием автоматических станций тестового откорма GENSTAR («Cooperl», Франция). Животные получали полнорационный комбикорм ПК-56-1, изготовленный согласно ГОСТ 21055-96 («Комбикорма полнорационные для беконного откорма свиней») по рецептурам СК-6, СК-7 и СК-52 (ОАО «Верхнехавский элеватор», Россия). Содержание основных питательных компонентов в рационе варьировало за период наблюдений в пределах, допускаемых техническими требованиями стандарта, и составляло для обменной энергии 12,61-13,59 МДж/кг, для массовой доли сырого протеина — 14,16-16,77 %, клетчатки — 3,83-4,49 %, лизина — 0,81-1,14 %, метионина и цистина — 0,51-0,73 %, триптофана — 0,15-0,21 %.
Выборка включала 71 хрячка породы дюрок отечественной репродукции. Все животные имели индивидуальный идентификатор (электронный чип). Выращивание животных на станциях осуществляли группами по 10-15 гол. (партиями) в возрасте от 70 сут (живая масса 29-33 кг). Продолжительность контрольного выращивания варьировала и завершалась в возрасте от 138 до 174 сут. В течение всего периода контрольного выращивания оценивали живую массу (BW, кг), среднесуточное потребление корма (DFI, г/сут) и показатели кормового поведения, в том числе общее время посещения кормовой станции в сутки (TPD, мин/сут), число посещений кормовой станции в сутки (NVD, ед.), среднее потребление корма за одно посещение (FPV, г), скорость поедания корма (FR, г/мин; FR = DFI/TPD), среднюю продолжительность одного посещения (TPV, мин; TPV = TPD/NVD). Аббревиатуры терминов соответствуют общепринятым сокращениям показателей конверсии корма, роста и кормового поведения (5).
Для оценки конверсии корма (FCR, кг/кг) использовали три способа. Первый предусматривал расчет показателя на основе начальной и конечной живой массы при контрольном выращивании и суммы потребленного корма:
FCRi =
2 i=i CRi= n (wn-W i ) ,
где FCR1 — конверсия корма за весь период выращивания, E i=i CR i=n — сумма потребленного корма за n наблюдений, начиная с i = 1; (W n - W1) — прирост живой массы за период выращивания, W n — живая масса в конце периода, W1 — живая масса в начале периода.
Второй способ был основан на оценке по средним значениям живой массы, ее среднесуточных приростов и конверсии корма, полученным в течение 10-суточных периодов:
ёж = e( ^S^s^^, [2]
(wi+io-wi+i )
где FCR2 — показатель конверсии корма по 10-суточным периодам для животного, E i^o CR^ (t+1_i+io) — суммарное потребление корма животным за 10-суточный период, W i +10 — живая масса животного на 10-е сут каждого 10-суточного периода, W i +1 — живая масса животного на 1-е сут каждого 10-суточного периода, i — шаг периода продолжительностью 10 сут при t > 1, t — число 10-суточных периодов.
В третьем случае использовали параметры суточной оценки про- дуктивности свиней с выравниванием показателя конверсии корма по отрицательным значениям (коррекция на среднюю величину положительных вариант) с учетом ежедневных колебаний живой массы и объема потребленного корма:
FC^ = Я=1 (^)л, [3]
где FCR3 — суточный показатель конверсии корма для животного, CR i — суммарное суточное потребление корма, W i + 1 — живая масса животного на момент наблюдения, W i — живая масса животного при предшествующем наблюдении, i — номер наблюдения.
Расчет значений среднесуточных приростов (ADG1, ADG2, ADG3) проводили для FCR1, FCR2, FCR3. Для оценки интенсивности роста хрячков определяли возраст достижения живой массы 100 кг (AGE100, сут).
Первичную информацию по контрольному выращиванию хрячков собирали в электронные файлы, на основе которых формировали базу данных с элементами логического контроля входной информации: даты, повторы, отсутствие одного из оцениваемых параметров. Первично массив из 99867 учетных записей был подвергнут контролю на соответствие закону нормального распределения анализируемых показателей (индивидуальных измерений). Выявленные выбросы в суточных данных каждого животного, превышавшие порог ±10,0 % отклонения от предыдущего значения, исключали из дальнейшей обработки. В анализ брали только особей, у которых число тестовых значений (по сумме учтенных) было не ниже 60,0 %. Конечная выборка составила 60 хрячков (потомки 13 отцов и 37 матерей) со средним показателем качества сбора и учета данных 91,2 % (лимиты от 63,3 до 99,5 %). База данных включала 4138 среднесуточных значений для каждого изучаемого параметра. При оценке конверсии корма для анализа динамики изменчивости комплекса показателей сформировали выборку по семи 10-суточным периодам от начала до завершения выращивания, в которую вошли 413 наблюдений.
Для оценки эффектов генетической и средовой природы было выбрано уравнение, характеризующееся наименьшим значением вариансы неучтенных факторов (вариансы ошибки) по модели дисперсионного анализа для фиксированных эффектов без взаимодействия факторов. Далее методом наименьших квадратов (least square means, LSM) c использованием программы STATISTICA 10 («StatSoft, Inc.», США) рассчитывали средние значения оценок:
y = µ + Feeding Station i + Batch k + Age l (1) + Sire j + e ikl j , [4] где y — показатели, характеризующие конверсию корма (DFI, TPD, NVD, FPV, FR, TPV, FCR, BW, ADG); µ — средняя популяционная константа; Feeding Station i — эффект кормовой станции ( i = 1…3); Batch k — эффект номера партии контрольного выращивания ( k = 1…4); Age l — эффект начального возраста контрольного выращивания ( l = 1…18) либо при аналогичной модели Age l (1) — эффект возраста животного за весь период контрольного выращивания ( l (1) = 1…102), Sire j — генетический эффект хряка-производителя (отцов оцениваемых потомков) ( j = 1…13), e ikl j — случайная ошибка (нераспределенная варианса).
Значения генетических и паратипических варианс особей для получения селекционно-генетических параметров рассчитывали по аналогичной модели посредством привлечения информации по родителям хрячков для построения аддитивной матрицы родства согласно подходу ограниченного максимального правдоподобия (REML, n = 110 гол., включая
60 гол., имеющих показатели собственной продуктивности):
y = µ + Test-day e + Feeding Station i + Batch k + Age lb 1 + Animal j + e eikl j , [5] где Test-day e — фиксированный эффект наблюдения в течение всего периода проведения эксперимента ( e = 1…226); Age lb 1 — начальный возраст контрольного выращивания, b 1 — коэффициент линейной регрессии, Animal j — рандомизированный эффект животного, имеющий нормальное распределение со средней, равной 0, и вариансой, равной A σ a2, где A — аддитивная матрица родства ( j = 1…110), e eikl j — случайная ошибка (нераспределенная варианса).
С помощью программы STATISTICA 10 («StatSoft, Inc.», США) были проведены расчеты оценок значений по признакам методом наименьших квадратов. Достоверность влияния организованных групп факторов, включенных в модель, определяли согласно методу MANOVA. При применении описательной статистики (показатели вариации), а также декомпозиции фенотипической изменчивости использовали общепризнанные подходы (19), при анализе варианс и расчете корреляций по модели для ряда взаимосвязанных признаков — программы семейства BLUPF90 (20).
Результаты. Средний возраст начала тестового откорма с использованием кормовых станций составил 74,2±1,0 сут (Cv = 10,6 %). Хрячки хорошо адаптировались, что позволило получать высокие среднесуточные приросты живой массы (950±19 г) (рис. 1). Динамика ее прироста по 10-суточным периодам была равномерной. Возраст достижения живой массы 100 кг составлял в среднем 149,9±1,0 сут (Cv = 5,0 %). Фенотипическая изменчивость оставалась в биологических пределах, достигая максимума вариации во II и III декадах откорма — соответственно 15,9 и 16,1 %, а минимума — в первый (13,7 %) и последний (12,0 %) периоды.
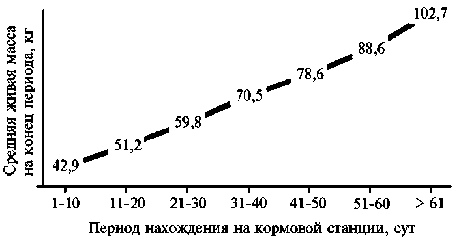
Рис. 1. Динамика прироста живой массы у свиней ( Sus scrofa ) породы дюрок по периодам выращивания на кормовых станциях (ООО «СГЦ», п. Верхняя Хава, Воронежская обл., июль 2017 года—март 2018 года).
Среди показателей кормового поведения наименьшая вариабельность была зарегистрирована для времени нахождения на кормовой станции (TPD, 85,3 мин/сут, Cv = 13,7 %) (табл. 1). Промежуточное положение отмечали для скорости поедания корма (FR, 27,6 г/мин) — Cv = 17,7 %. Для средней продолжительности одного посещения (TPV, 19,6 мин) и количества потребляемого корма за одно посещение (FPV, 499,1 г) фенотипическая вариация находилась в пределах 21,6-21,8 %. Наибольшей вариабельностью (Cv = 27,4 %) характеризовалось число посещений кормовой станции в сутки (NVD, 5,2 ед.).
Для расчета конверсии корма (FCR) мы применили три различных подхода. Колебания живой массы свиней, обусловленные влиянием тех- нологических, паратипических факторов, состоянием здоровья животных, могут быть значительными. При первом варианте расчета (FCR1) как высокие среднесуточные приросты, так и снижения живой массы маскиру- ются. Второй способ расчета (FCR2) ориентирован на определение конверсии корма по декадным периодам, что позволяет сгладить колебания среднесуточного изменения живой массы и более надежно оценить показатель. Третий подход (FCR3) предполагает использование в расчетах суточных данных по потреблению корма и приросту живой массы у свиней с учетом положительных значений динамики изменчивости для конверсии корма.
Параметры конверсии корма, рассчитанные тремя способами, характеризовались разной изменчивостью (табл. 1). Так, в вариантах FCR2 и FCR3 величина Сv составила соответственно 23,2 и 19,2 % при средних показателях 3,08 и 2,52 кг/кг. Для FCR1 была выявлена низкая степень вариации показателя (12,3 %) при его близком среднем значении — 2,55 кг/кг.
-
1. Показатели эффективности использования корма и кормового поведения в выборке свиней ( Sus scrofa ) породы дюрок (ООО «СГЦ», п. Верхняя Хава, Воронежская обл., июль 2017 года—март 2018 года)
-
2. Разложение компонентов фенотипической изменчивости (%) по комплексу признаков конверсии корма в долях генетических и средовых факторов для выборки свиней ( Sus scrofa ) породы дюрок (ООО «СГЦ», п. Верхняя Хава, Воронежская обл., июль 2017 года—март 2018 года)
|
Показатель |
1 M ±SEM |
] σ 1 |
Cv , % |
|
AGE 100 , сут |
149,9±1,0 |
7,5 |
5,0 |
|
ADG 1 , г |
950±19 |
150 |
15,8 |
|
ADG 2 , г |
892±21 |
162 |
18,2 |
|
ADG 3 , г |
986±24 |
187 |
19,0 |
|
DFI, г/сут. |
2309,1±36,5 |
282,7 |
12,2 |
|
FCR1, кг/кг |
2,55±0,04 |
0,31 |
12,3 |
|
FCR2, кг/кг |
3,08±0,09 |
0,72 |
23,2 |
|
FCR3, кг/кг |
2,52±0,06 |
0,48 |
19,2 |
|
TPD, мин/сут |
85,3±1,5 |
11,7 |
13,7 |
|
NVD, ед. |
5,2±0,2 |
1,4 |
27,4 |
|
FPV, г |
499,1±13,9 |
107,6 |
21,6 |
|
FR, г/мин |
27,6±0,6 |
4,9 |
17,7 |
|
TPV, мин |
19,6±0,6 |
4,3 |
21,8 |
П р и м е ч а н и е. Аббревиатуры соответствуют общепринятым сокращениям для обозначения показателей (5).
Компонент изменчивости
|
Показатель |
отец потомка (sire) |
кормовая станция (feeding station) |
номер партии (batch) |
возраст постановки (age) |
e |
R2 |
F |
Показатели роста
*, **, *** Доля влияния компонента изменчивости на показатель статистически значима соответственно при р < 0,05, р < 0,01 и р < 0,001.
сокодостоверные результаты ( F = 4,57; p < 0,001), которые объясняют до 84,4 % общей изменчивости по фенотипу в модели. Иными словами, линейная зависимость предсказанных (ожидаемых) результатов для конверсии корма по первому варианту расчета в сравнении с наблюдаемыми была лимитирована строго выбранными компонентами дисперсии при минимальной ошибке вариансы.
Однако паратипические факторы, такие как кормовая станция и номер партии контрольного выращивания, имели минимальное влияние (0,0-7,1 %) во всех вариантах FCR. Исключение составил возраст постановки на тестовый откорм: высокая обусловленность этого фактора, по всей видимости, связана с начальной живой массой животных в группах на станциях откорма. Для FCR3 по сравнению с FCR2 и FCR1 было показано значимое влияние генотипа отца, по доле превосходящее сопоставляемые показатели соответственно на 10,9 процентного пункта (п.п.) и 3,7 п.п. Можно утверждать, что накапливаемая база данных позволит привлечь в организованную группу факторов сезонные колебания (влияние года, месяца и дня контроля показателей).
Среднесуточные приросты по ADG (1…3) и величина живой массы хрячков на откорме по силе влияния были на 6,5-9,0 % детерминированы аддитивно-генетической группой при R2 = 74,5-84,2 % ( F = 2,464,49; p < 0,01…0,001).
3. Популяционно-генетические константы для показателей эффективности использования корма и кормового поведения в выборке свиней ( Sus scrofa ) породы дюрок (ООО «СГЦ», п. Верхняя Хава, Воронежская обл., июль 2017 года—март 2018 года)
Наследуемость показателя конверсии корма (FCR3) была низкой (h2 = 0,019), что, по нашему мнению, связано с высокой долей суточной (средовой) изменчивости признака в изученной выборке (табл. 3). Были получены умеренные коэффициенты наследуемости для FR — 0,458, TPV — 0,368, FPV — 0,258, NVD — 0,218. Низкое значение отмечали для TPD — 0,134. Высокую долю генетической составляющей выявили для показателя живой массы (h2 = 0,744), что в целом было характерно для признаков мясной продуктивности у специализированной породы дюрок.
Взаимосвязи между кормовым поведением, конверсией корма и живой массой колебались по направлению и были умеренными по величине. У свиней живую массу в наибольшей мере предопределяли показатели DFI ( r g = 0,565) и FR ( r g = 0,588). Обратную генетическую зависимость обнаружили для TPV ( r g = - 0,516), то есть отбор животных с долгим пребыванием на станции в течение одного посещения не позволит проводить эффективную селекцию по живой массе. Отбор по FCR3 будет более результативен при таких параметрах кормового поведения животных, как меньшая продолжительность пребывания на кормовой станции в сутки (TPD, r g = 0,585), высокое потребление корма за посещение (FPV,
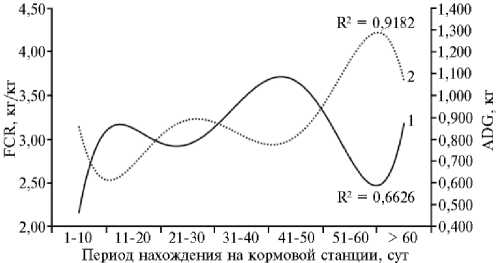
Рис. 2. Зависимость показателей конверсии корма ( FCR 2 , 1) и среднесуточных приростов живой массы (ADG 2 , 2) в выборке свиней ( Sus scrofa ) породы дюрок по контрольным декадам выращивания (ООО «СГЦ», п. Верхняя Хава, Воронежская обл., июль 2017 года—март 2018 года).
Анализ колебаний учетных показателей
rg = -0,274) и высокая скорость поедания корма (FR, rg = -0,368), меньшее число посещений кормовой станции в сутки (NVD, rg = 0,368). В этой связи для программы разведения свиней породы дюрок наиболее предпочтительна разработка селекционного ин- декса, в котором учитывается комплекс двух факторов — кормового поведения и конверсии корма.
по 10-суточным циклам
(рис. 2) выявил поступательную динамику роста: соответственно от 851±51 г для ADG2 и 2,19±0,13 кг/кг для FCR2 в начале откорма (1-10-е сут) до 1072±41 г и 3,31±0,18 кг/кг в конце тестирования (> 61 сут). Тренд по показателю конверсии корма (полином 5-го порядка) имел форму синусои- дальной кривой, динамика которой описывалась с точностью по периодам R2 = 66,3 %. Аналогичное распределение было получено для тренда пока- зателя ADG2 (полином 5-го порядка), который имел обратную («зеркальную») зависимость с FCR2 (rp = -0,592). Следовательно, высокие приросты живой массы обеспечивались лучшими результатами по конверсии корма.
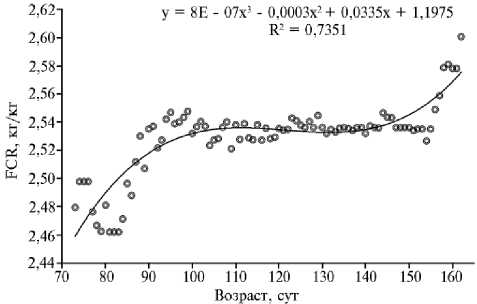
Рис. 3. Динамика показателей LSM (least square means) оценок конверсии корма (FCR 3 ) в зависимости от возраста тестового откорма свиней ( Sus scrofa ) породы дюрок (ООО «СГЦ», п. Верхняя Хава, Воронежская обл., июль 2017 года—март 2018 года).
С целью сглаживания эффектов среды (кормовой станции и номера тестируемой партии хрячков) были получены оценки по методу наименьших квадратов (LSM). Результаты (рис. 3) описаны полиномом 3-го порядка (в соответствии с уравнением модели точность прогноза R2 = 73,5 %). С даты постановки на откорм и до 100 сут наблюдалось плавное повышение конверсии корма за счет высокой интенсивности роста и раз- вития животных. Далее, в период 100-140 сут, величина конверсии стабилизировалась и формировались так называемое плато с минимальной вариацией (2,52-2,54 кг/кг). Начиная с возраста 140 сут, происходило резкое возрастание кривой тренда, что указывало на снижение эффективности откорма свиней за счет депонирования жировых отложений. Интенсив- ность роста и относительный прирост замедлялись, сигнализируя о завершении физиологической и биологической фазы роста животных.
Индивидуальные различия в эффективности использования корма и кормовом поведении, обусловленные генетическими факторами, позво- ляют выявить ценные молекулярные биомаркеры для прогнозирования указанных признаков, а также их применения в селекции свиней (8, 9, 21). Кроме того, наблюдения за кормовым поведением могут использоваться в качестве инструмента в автоматизированных системах мониторинга для оценки состояния здоровья животных и лучшего выявления заболеваний, с целью контроля менеджмента (8, 22, 23). Полученные нами данные найдут применение при разработке программ геномной селекции, с развитием которых в настоящее время связывают генетический прогресс в племенном животноводстве (24, 25).
Таким образом, при откорме свиней на автоматических станциях влияние генетических факторов обусловливало 15,4 % общей изменчивости по показателю конверсии корма FCR3, при этом для параметров кормового поведения аддитивная вариация составляла 28,2 % (общее время нахождения на кормовой станции в сутки, TPD) и 30,8 % (число посещений кормовой станции в сутки, NVD). Анализ генетических корреляций между признаками показывает перспективность использования параметров кормового поведения для повышения надежности оценки конверсии корма. Динамика конверсии корма в связи с возрастом животных имеет тенденцию к увеличению, несмотря на степень колебания среднесуточных приростов по периодам откорма. Использование суточной вариации конверсии корма (FCR3) наравне с традиционной оценкой показателя (FCR1) позволит эффективнее отбирать терминальных хрячков за счет привлечения таких селекционных параметров, как кормовое поведение.
Список литературы Генетические и паратипические факторы, характеризующие эффективность использования корма у свиней породы Дюрок
- Do D.N., Strathe A.B., Jensen J., Mark T., Kadarmideen H.N. Genetic parameters for different measures of feed efficiency and related traits in boars of three pig breeds. J. Anim. Sci., 2013, 91(9): 4069-4079 ( ) DOI: 10.2527/jas.2012-6197
- Young R.J., Lawrence A.B. Feeding behaviour of pigs in groups monitored by a computerized feeding system. Anim. Prod., 1994, 58(1): 145-152 ( ) DOI: 10.1017/S0003356100007182
- Maselyne J., Saeys W., Van Nuffel A. Review: Quantifying animal feeding behaviour with a focus on pigs. Physiol. Behav., 2015, 138: 37-51 ( ) DOI: 10.1016/j.physbeh.2014.09.012
- Hyun Y., Ellis M. Effect of group size and feeder type on growth performance and feeding patterns in finishing pigs. J. Anim. Sci., 2002, 80(3): 568-574 ( ) DOI: 10.2527/2002.803568x
- Baumung R., Gerhard G., William A., Söelkner J. Feed intake behaviour of different pig breeds during performance testing on station. Arch. Tierz., Dummerstorf., 2006, 49(1): 77-88.
- Lewis C.R.G., McGlone J.J. Modelling feeding behaviour, rate of feed passage and daily feeding cycles, as possible causes of fatigued pigs. Animal, 2008, 2(4): 600-605 ( ) DOI: 10.1017/S1751731108001766
- Rohrer G.A., Brown-Brandl T., Rempel L.A., Schneider J.F., Holl J. Genetic analysis of behavior traits in swine production. Livest. Sci., 2013, 157(1): 28-37 ( ) DOI: 10.1016/j.livsci.2013.07.002
- Brown-Brandl T., Rohrer G., Eigenberg R. Analysis of feeding behavior of group housed growing-finishing pigs. Comput. Electron. Agr., 2013, 96: 246-252 ( ) DOI: 10.1016/j.compag.2013.06.002
- Reyer H., Shirali M., Ponsuksili S., Murani E., Varley P.F., Jensen J., Wimmers K. Exploring the genetics of feed efficiency and feeding behaviour traits in a pig line highly selected for performance characteristics. Mol. Genet. Genomics, 2017, 292(5): 1001-1011 ( ) DOI: 10.1007/s00438-017-1325-1
- Herd R., Arthur P. Physiological basis for residual feed intake. J. Anim. Sci., 2009, 87(Suppl. 14): E64-E71 ( ) DOI: 10.2527/jas.2008-1345
- Hall A.D., Hill W.G., Bampton P.R., Webb A.J. Genetic and phenotypic parameter estimates for feeding pattern and performance test traits in pigs. Anim. Sci., 1999, 68: 43-48.
- Chen C., Misztal I., Tsuruta S., Herring W., Holl J., Culbertson M. Influence of heritable social status on daily gain and feeding pattern in pigs. J. Anim. Breed. Genet., 2010, 127(2): 107-112 ( ) DOI: 10.1111/j.1439-0388.2009.00828.x
- Morgan C.A., Emmans G.C., Tolkamp B.J., Kyriazakis I. Analysis of the feeding behavior of pigs using different models. Physiol. Behav., 2000, 68(3): 395-403 ( ) DOI: 10.1016/S0031-9384(99)00195-X
- Rauw W.M., Soler J., Tibau J., Reixach J., Gomez Raya L. Feeding time and feeding rate and its relationship with feed intake, feed efficiency, growth rate, and rate of fat deposition in growing Duroc barrows. J. Anim. Sci., 2006, 84(12): 3404-3409 ( ) DOI: 10.2527/jas.2006-209
- Тагиров X.X., Асаев Э.Р. Оценка мясности подсвинков по выходу питательных веществ и биоконверсии корма в мясную продукцию. Известия Оренбургского государственного аграрного университета, 2007, 1: 118-120.
- Ларина О.В., Аристов А.В., Кудинова Н.А. Мясная продуктивность и конверсия кормов при откорме свиней разного генотипа. Вестник Рязанского государственного агротехнологического университета им. П.А. Костычева, 2017, 2: 26-29.
- Гальперн И.Л., Джолова М.Н. Метод селекции на повышение конверсии корма при одновременном увеличении яичной продуктивности и продолжительности использования несушек. Генетика и разведение животных, 2015, 1: 30-34.
- Левахин B.И., Ажмулдинов Е.А., Титов М.Г., Ласыгина Ю.А., Рябов Н.И. Комплексная оценка мясной продуктивности бычков различных генотипов по эффективности конверсии корма. Вестник Курской государственной сельскохозяйственной академии, 2015, 7: 145-146.
- Кузнецов В.М. Основы научных исследований в животноводстве. Киров, 2006.
- Misztal I., Tsuruta S., Strabel T., Auvray B., Druet T., Lee D.H. BLUPF90 and related programs (BGF90). Proc. 7th World Congress on genetics applied to livestock production. Montpellier, Communication No. 28-27, 2002, 28: 21-22.
- Ding R., Yang M., Wang X., Quan J., Zhuang Z., Zhou S., Li S., Xu Z., Zheng E., Cai G., Liu D., Huang W., Yang J., Wu Z. Genetic architecture of feeding behavior and feed efficiency in a Duroc pig population. Frontiers in Genetics, 2018, 9: 220 ( ) DOI: 10.3389/fgene.2018.00220
- Weary D., Huzzey J., Von Keyserlingk M. Board-invited review: using behavior to predict and identify ill health in animals. J. Anim. Sci., 2009, 87(2): 770-777 ( ) DOI: 10.2527/jas.2008-1297
- Cross A.J., Keel B.N., Brown-Brandl T.M., Cassady J.P., Rohrer G.A. Genome-wide association of changes in swine feeding behaviour due to heat stress. Genet. Sel. Evol., 2018, 50: 11 ( ) DOI: 10.1186/s12711-018-0382-1
- Сермягин А.А., Гладырь Е.А., Харитонов С.Н., Ермилов А.Н., Стрекозов Н.И., Брем Г., Зиновьева Н.А. Полногеномный анализ ассоциаций по хозяйственно полезным признакам скота в российской популяции голштинской породы. Сельскохозяйственная биология, 2016, 51(2): 182-193 ( ) DOI: 10.15389/agrobiology.2016.2.182rus
- Зиновьева Н.А., Сермягин А.А., Костюнина О.В. Геномная селекция -новая стратегия генетического совершенствования свиней. Животноводство России, 2018, тематический выпуск «Свиноводство»: 53-55.
