Генетический анализ количественных признаков у инбредных линий озимой ржи ( Secale cereale L.) в диаллельных скрещиваниях
 в диаллельных скрещиваниях")
Автор: Гончаренко А.А., Крахмалев С.В., Макаров А.В., Ермаков С.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетические основы селекции
Статья в выпуске: 1 т.50, 2015 года.
Бесплатный доступ
Один из эффективных методов оценки инбредных линий озимой ржи - диаллельный анализ, позволяющий разложить генотипическую вариансу на общую (ОКС) и специфическую (СКС) комбинационную способности и количественно оценить вклад основных видов генных взаимодействий в экспрессию признаков. Целью настоящего исследований было изучение генетических особенностей инбредных линий посредством диаллельных скрещиваний. Испытывали 5 инбредных линий озимой ржи ( Secale cereale L.) и 10 межлинейных гибридов F 1, полученных по неполной диаллельной схеме (B. Griffing, 1956). Родительские линии и гибриды F 1 сравнивали в 2011-2012 годах в полевом опыте по схеме латинского прямоугольника (6*3*3) на делянках 8,8 м 2 в 3 повторениях при норме высева 500 зерен на 1 м 2. Сравнительную оценку генных эффектов проводили по 17 признакам. Определяли генетические параметры D, H 1, H 2, √H 1/D, Н 2/4Н 1 и h 2 по B.I. Hayman (1954), а также вклад эффектов ОКС и СКС в генотипическую вариансу. Показано, что по соотношению эффектов ОКС:СКС изучаемые признаки разделяются на две группы. Первую группу составили 5 признаков с низкой (h 2 = 0,10-0,22) наследуемостью (урожайность, число продуктивных стеблей на 1 м 2, число зерен в колосе, высота растений и содержание крахмала). Вклад эффектов ОКС в этой группе был относительно низким и варьировал от 7,0 до 36,4 %, а экспрессия признаков сильно зависела от эффектов доминирования и эпистаза. Вторую группу составили 12 признаков с относительно высокой наследуемостью (h 2 = 0,28-0,62), у которых аддитивная дисперсии превышала компоненту доминирования и варьировала от 50,3 до 82,0 %. Таковыми оказались зимостойкость, масса 1000 зерен, натура зерна, число падения, высота амилограммы, вязкость водного экстракта, температура клейстеризации крахмала, формоустойчивость подового хлеба, объем формового хлеба, содержание белка в зерне, поражение снежной плесенью и бурой ржавчиной. Шесть признаков из этой группы выделялись сильным взаимодействием генотип-среда. Сделано заключение, что селекционное улучшение инбредных линий по этим признакам целесообразно проводить по принципу кумулятивного накопления ценных генов с помощью рекуррентного отбора.
Озимая рожь, инбредные линии, количественные признаки, диаллельный анализ, общая и специфическая комбинационная способность, генетические параметры
Короткий адрес: https://sciup.org/142133571
IDR: 142133571 | УДК: 633.14:631.523.4:575.222.6 | DOI: 10.15389/agrobiology.2015.1.75rus
Genetic research of quantitative traits of inbred lines of winter rye ( Secale cereale L.) in diallel crossings
One of effective methods for estimation of inbred lines of winter rye is the diallel analysis allowing to spread out genotypic variance to general (GCA) and specific (SCA) combinational abilities and to estimate quantitatively the contribution of the basic gene interactions in expression of traits. The purpose of our research was studying genetic features of inbred lines of rye by means of diallel crossings. Tested 5 inbred lines of winter rye and 10 interline F 1 hybrids were obtained according to incomplete diallel scheme (B. Griffing, 1956). Parental lines and F 1 hybrids were compared in 2011 and 2012 in a field experiments under the scheme of a Latin square (6*3*3) on the plots of 8.8 m 2 in 3 replicates at a sowing rate of 500 seeds per 1 m 2. Comparative estimation of gene effects has been carried out for 17 traits. The genetic parameters D, H1, H2, √H1/D, Н2/4Н1 and h 2 according to B.I. Hayman (1954) were evaluated, and also the contribution of GCA and SCA effects into genotypic variance was assessed. It was shown that according to effects of GCA to SCA ratio the traits can be divided into two groups. The first group contained 5 traits with low heritability (h 2 = 0.10-0.22), exactly the productivity, number of productive stalks per 1 m 2, number of grains in an ear, height of plant and the content of starch. In this group the contribution of GCA effects was rather low and varied from 7.0 up to 36.4 %, and the expression of traits strongly depended on domination and epistasis. In the second group there were 12 traits with rather high heritability (h 2 = 0.28-0.62) for which an additive dispersion exceeded the component of domination and varied from 50.3 up to 82.0 %. These high-hereditable traits were the winter hardiness, weight of 1000 grains, a grain unit, number of falling, amylogramm peak, viscosity of water extract, temperature of pasting starch, fluidity of dough, loaf volume, the content of protein in grain, sensitivity to snow mould and brown rust. For six traits of this group a strong genotype-environment interaction was characteristic. The conclusion is made that cumulative accumulation of valuable genes by means of recurrent selection is advisable for improvement of inbred lines on these traits in the course of breeding
Текст научной статьи Генетический анализ количественных признаков у инбредных линий озимой ржи ( Secale cereale L.) в диаллельных скрещиваниях
Одно из приоритетных направлений в селекции озимой ржи — использование эффекта гетерозиса и получение гибридов F 1 на основе цитоплазматической мужской стерильности (ЦМС) (1). Созданные в Германии гибриды ржи в настоящее время занимают около 70 % от всех посевов под культурой и существенно потеснили популяционные сорта в большинстве стран Центральной Европы, возделывающих рожь. По сообщению H.H. Geiger и T. Miedaner (2), за период с 1982 по 2005 год среднегодовая прибавка урожая за счет селекции гибридных сортов составила 51 кг, популяционных — 30 кг. К тому же превосходство гибридов над сортами-популяциями проявляется не только по урожайности, но и по более короткой длине стебля, лучшей устойчивости к полеганию, по качеству зерна и другим признакам (3).
Для эффективной селекции инбредных линий озимой ржи очень важно знать, какие виды генных взаимодействий участвуют в обеспечении наследственного полиморфизма признаков, каков характер их экспрессии, в какой коррелятивной связи они находятся, какова доля вклада конкретных генетических эффектов (аддитивных, доминантных и различного рода эпистатических) и в какой степени такие эффекты модифицируются факторами внешней среды. Для решения этой задачи в селекции широко применяется диаллельный анализ, позволяющий разложить генотипиче- скую вариансу признака на компоненты — общую (ОКС) и специфическую (СКС) комбинационную способность и количественно оценить вклад тех или иных генных взаимодействий в экспрессию признака.
Практически все селектируемые признаки у ржи полигенны по своей природе, а их фенотипическое проявление есть сумма эффектов аллельного и неаллельного взаимодействия по многим генам, корректируемых в значительной степени условиями внешней среды. Эффект гетерозиса по селектируемым признакам в системе диаллельных скрещиваний инбредных линий у ржи изучен весьма слабо. Имеются данные, что варианса ОКС по большинству признаков значительно выше, чем варианса СКС. H.H. Geiger (4) приводит сводку долевого участия вариансы СКС по ряду культур, из которой следует, что для урожайности ржи она составляет 21 %, что в 2,0-2,5 раза ниже соответствующего показателя у кукурузы и сахарной свеклы. Поэтому оценка линий озимой ржи по ОКС считается более важной, чем по СКС, а на практике эффекты СКС принимают в расчет только на заключительном этапе изучения экспериментальных гибридов (2).
Замечено (5), что фенотипическое выражение многих признаков у инбредных линий per se может служить основанием для прогнозирования их ОКС. Поэтому самоопыленные линии предлагается интенсивно отбирать на высокую собственную продуктивность и неоднократно тестировать на ОКС (1). Однако достоверная корреляция между продуктивностью инбредных линий и продуктивностью их гибридов проявляется не по всем признакам. Причиной тому служат различия в степени их наследуемости. Именно высоконаследуемые признаки дают заметный генетический сдвиг при непрямом отборе линий в процессе селекции (6). Если в генетическом контроле признака преобладают эффекты доминирования, то отбор должен проводиться не по продуктивности линий per se, а по продуктивности межлинейных гибридов F 1 (7).
У исследователей пока нет полной ясности относительно механизма генетической детерминации признака урожайности и связанных с ней структурных компонентов. То же самое можно сказать и в отношении признаков качества зерна у ржи. Весьма неоднозначны выводы о структуре наследственной вариансы селектируемых признаков и доли вклада конкретных генетических эффектов — аддитивных, доминирования, эпистаза. H.H. Geiger (5) определил, что для большинства признаков озимой ржи аддитивная варианса гораздо больше, чем все другие компоненты генетической вариансы, вместе взятые. Считается, что даже по урожаю зерна неаддитивные эффекты дают меньший вклад в генетическую вариансу, чем аддитивные, хотя у некоторых гибридов от скрещивания линий с высокой ОКС эффект доминирования может быть значительным. Линии могут различаться по адаптивности к условиям возделывания и оказывать существенное влияние на вариансу взаимодействия генотип—среда. На преобладающий вклад аддитивной вариансы указывают и другие исследователи (6, 8, 9).
Значительная часть неаддитивной дисперсии у ржи приходится на эпистатическое взаимодействие генов (10). В зависимости от генотипа и внешних условий эта компонента генотипической вариансы может достоверно увеличивать или уменьшать величину признака. В последнем случае селекционер вынужден искать способы минимизации негативных последствий неаллельного взаимодействия, с тем чтобы максимально использовать эффекты СКС (11).
Нашей задачей было изучение комбинационной способности ин- бредных линий озимой ржи в системе диаллельных скрещиваний и сравнительная оценка основных видов генных взаимодействий, влияющих на величину селекционно важных признаков.
Методика. Исходным материалом послужили 5 инбредных линий озимой ржи (Н-649, Н-1078, Н-1179, Н-451, Н-842) и 10 межлинейных гибридов F 1 , полученных по неполной диаллельной схеме (II метод по Гриффингу) (12). Исследуемые линии последовательно прошли многократный инцухт (S 14 -S 16 ) и были глубоко гомозиготными. Скрещивание линий выполняли в условиях Московской области в 2010 году в изоляционных домиках площадью 25 м2, где ЦМС-аналоги вышеперечисленных линий высевались рядом с фертильными формами для переопыления. Родительские линии и межлинейные гибриды F 1 изучали в полевом опыте, который закладывали в 2011-2012 годах по схеме латинского прямоугольника (6x3x3) на 8-рядковых делянках площадью 8,8 м2 в трех повторностях при норме высева 500 зерен на 1 м2.
По каждой делянке учитывали урожайность (т/га), зимостойкость, число продуктивных стеблей на 1 м2, число зерен в колосе, высоту растений, массу 1000 зерен, натуру зерна, число падения, высот амилограммы, вязкость водного экстракта, температур клейстеризации крахмала, формо-устойчивость теста (H/D), объем формового хлеба, содержание белка и крахмала в зерне, устойчивость к поражению снежной плесенью и бурой ржавчиной. Общую и специфическую комбинационную способность линий определяли по B. Griffing (12), генетический анализ диаллельного комплекса выполняли по B.I. Hayman (13): определяли генетические параметры D, H 1 , H2, V H 1 /D, Н2/4Н | и h2, а также вычисляли долю вклада эффектов ОКС и СКС в генотипическую вариансу признака.
Для статистической обработки данных использовали компьютерную программу Agros (2.13).
Результаты. Полевое испытание проводили в разные по погодным условиям годы. В июне и июле 2011 года преобладала жаркая и сухая погода, которая ускорила прохождение фаз колошения, цветения и налива зерна, посевы не полегли, уборка наступила на 10 сут раньше многолетнего срока. В 2012 году растения перезимовали хорошо, густота сформировавшегося стеблестоя была высокой, но обильные осадки в июне (150 % от многолетней нормы) привели к раннему (в фазу цветения) полеганию посевов, что неблагоприятно отразилось на урожайности и качестве зерна.
Статистическая обработка исходных данных выявила достоверные различия между родительскими линиями и гибридами F 1 по всем изучаемым признакам. Дисперсионный анализ комбинационной способности показал, что в наследственную вариансу признаков достоверный вклад вносили как ОКС, так и СКС. Однако доля вклада этих компонентов сильно варьировала в зависимости от признака.
Взяв за основу относительный вклад ОКС и СКС в генотипическую вариансу, мы разделили все изученные признаки на две группы. В первую отнесли 5 признаков с низкой (h2 = 0,10-0,22) наследуемостью (урожайность, число продуктивных стеблей на 1 м2, число зерен в колосе, высота растений и содержание крахмала), у которых по ОКС вклад эффектов был ниже, чем по СКС, и в среднем за годы исследований варьировал в пределах 7,0-36,4 %. Во вторую группу объединили 12 признаков с относительно высокой (h2 = 0,28-0,62) наследуемостью, у которых аддитивная компонента дисперсии превышала компоненту доминирования и изменялась от 50,3 до 82,0 %. Таковыми оказались зимостойкость, масса 1000 зерен, натура зерна, число падения, высота амилограммы, вязкость водного экстракта, температура клейстеризации крахмала, формоустойчи-вость подового хлеба, объем формового хлеба, содержание белка в зерне, поражение снежной плесенью и бурой ржавчиной (табл. 1).
-
1. Группы количественных признаков по величине вклада общей и специфической комбинационной способности в их генотипическую вариансу у озимой ржи ( Secale cereale L.) по годам наблюдения (Московская обл.)
Признак |
2011 год
| 2012 год |
Среднее
1-я
группа
(ОКС < СКС)
Урожайность
4,4/94,5
9,5/87,9
7,0/91,2
Число продуктивных стеблей на 1 м2
43,9/56,1
20,6/68,4
32,3/62,3
Число зерен в колосе
43,0/50,4
29,8/63,1
36,4/56,8
Высота растений
20,1/79,3
23,8/74,9
22,0/77,1
Содержание крахмала
2-я
41,1/56,0
группа
17,3/79,8 (ОКС > СКС)
29,2/67,9
Зимостойкость
73,5/25,6
82,1/8,6
77,8/17,1
Масса 1000 зерен
54,8/43,1
66,7/31,1
60,8/37,1
Вязкость водного экстракта
66,5/33,1
82,2/15,1
74,4/24,1
Число падения
45,5/54,4
69,3/29,6
57,4/42,0
Высота амилограммы
77,1/22,9
86,9/12,2
82,0/17,6
Температура клейстеризации крахмала
53,4/38,7
85,1/9,7
69,3/24,2
Натура зерна
47,2/52,6
53,4/42,8
50,3/47,7
Формоустойчивость теста
29,4/58,8
83,2/16,6
56,3/37,7
Объемный выход хлеба
74,4/23,2
60,8/37,4
67,6/30,3
Содержание белка
55,7/42,4
58,6/37,9
57,2/40,2
Устойчивость к снежной плесени
82,5/15,2
48,4/47,3
65,5/31,3
Устойчивость к бурой ржавчине
67,8/20,6
74,2/18,0
71,0/19,3
Примечание. Перед косой чертой — доля (%) вариансы общей комбинационной способности (ОКС), после черты — специфической комбинационной способности (СКС).
О селекционной ценности родительских форм можно судить по их ОКС. В наших опытах изучаемые линии имели как положительные, так и отрицательные оценки эффектов ОКС (табл. 2). Относительно высокую ОКС по урожайности и некоторым другим признакам показали линии Н-649, Н-1179, Н-451. В селекционных программах они могут служить ценными компонентами для синтеза высокоурожайных гибридов с высоким качеством зерна. Дифференциация между линиями позволяет заключить, что каждая из них содержит специфический комплекс генов, по-разному влияющих на комбинационную способность. Большой контраст между инбредными линиями и гибридами F 1 по урожайности дополнительно подтверждает, что генетическая дисперсия этого признака большей частью обусловлена внутрилокусным доминированием и неаллельным взаимодействием генов.
-
2. Эффекты общей комбинационной способности (g j ) у инбредных линий озимой ржи ( Secale cereale L.) по группам количественных признаков (Московская обл., среднее за 2011-2012 годы)
Признак
| Н-649
| Н-1078 |
| Н-1179 |
Н-451 |
| Н-842 |
r
1-я группа (ОКС < СКС)
Урожайность:
ц/га
22,9
21,0
34,5
28,7
28,9
0,10-0,55
g j
4,6
- 0,8
2,1
- 1,9
- 4,0
Число продуктивных стеблей на 1 м2:
шт.
287
362
443
399
512
0,73-0,79
g j
- 33,0
13,4
38,5
- 31,6
12,8
Число зерен в колосе:
шт.
30,3
35,4
29,9
37,6
34,2
0,74-0,78
g j
0,3
- 0,6
- 3,4
2,3
1,4
Высота растений:
см
91,7
89,0
117,5
90,7
94,2
0,91-0,98*
g j
- 0,88
- 4,3
6,5
- 0,65
- 0,72
Содержание крахмала:
%
56,7
57,2
55,5
57,4
56,4
0,89-0,95*
g j
0,28 2-я г р у
0,15 - 0,67
п п а (ОКС > СКС)
0,59
- 0,35
Зимостойкость:
%
90,6
77,9
94,8
73,5
94,3
0,95-0,98*
g j
0,9
- 3,4
4,6
- 5,4
3,4
Масса 1000 зерен:
-
3. Сравнительная оценка параметров генетической изменчивости количественных признаков с различным соотношением общей и специфической комбинационной способности у озимой ржи ( Secale cereale L.) по годам наблюдения (Московская обл.)
|
г |
26,6 |
16,8 |
26,2 |
19,2 |
16,8 |
0,96-0,99* |
|
g i |
2,4 |
- 1,8 |
1,7 |
- 0,3 |
- 2,0 |
|
|
Вязкость водного экстракта: |
||||||
|
сП |
8,0 |
5,5 |
7,1 |
3,4 |
3,9 |
0,98-0,99* |
|
g i |
1,19 |
- 0,18 |
0,82 |
- 1,03 |
- 0,86 |
|
|
Число падения: |
||||||
|
с |
103 |
174 |
235 |
313 |
161 |
0,95-0,99* |
|
g i |
- 24,4 |
- 12,5 |
9,1 |
41,8 |
- 14,1 |
|
|
Высота амилограммы: |
||||||
|
ЕА |
169 |
132 |
224 |
594 |
114 |
0,91-0,98* |
|
g i |
5,2 |
- 83,3 |
- 9,2 |
137,1 |
- 49,8 |
|
Примечание. ОКС — общая комбинационная способность, СКС — специфическая комбинационная способность, g i — величина эффекта ОКС i -й линии.
* Достоверно на 5 % уровне значимости.
|
Признак |
Отношение ОКС:СКС |
Генетические параметры |
||||
|
D |
1 H |
H1/D |
1 H-IH- | |
h2 |
||
|
2011 |
год |
|||||
|
Урожайность |
0,1 |
9,3 |
2268* |
15,6 |
0,247 |
0,10 |
|
Число продуктивных стеблей на 1 м2 |
0,5 |
4492* |
67274* |
3,9 |
0,217 |
0,18 |
|
Число зерен в колосе |
0,6 |
22,7 |
148,9* |
2,6 |
0,212 |
0,22 |
|
Содержание крахмала |
0,4 |
1,9* |
23,2* |
3,5 |
0,241 |
0,10 |
|
Высота растений |
0,3 |
206* |
1092* |
2,3 |
0,240 |
0,10 |
|
Высота амилограммы |
4,7 |
48615* |
86184* |
1,3 |
0,198 |
0,62 |
|
Зимостойкость |
4,5 |
281,9* |
321,9* |
1,1 |
0,210 |
0,49 |
|
Вязкость водного экстракта |
3,1 |
4,9* |
13,2* |
1,6 |
0,240 |
0,49 |
|
Масса 1000 зерен |
1,6 |
12,2* |
46,9* |
2,0 |
0,243 |
0,54 |
|
Число падения |
1,4 |
5478* |
12957* |
1,5 |
0,207 |
0,29 |
|
2012 |
год |
|||||
|
Урожайность |
0,1 |
41,7 |
1331* |
5,6 |
0,238 |
0,10 |
|
Число продуктивных стеблей на 1 м2 |
0,5 |
13887* |
44926* |
1,8 |
0,178 |
0,04 |
|
Число зерен в колосе |
0,6 |
15,3 |
246,7* |
4,0 |
0,233 |
0,05 |
|
Содержание крахмала |
0,4 |
2,1* |
9,5* |
2,1 |
0,245 |
0,22 |
|
Высота растений |
0,3 |
104* |
899* |
2,9 |
0,240 |
0,10 |
|
Высота амилограммы |
4,7 |
28052* |
14855* |
0,7 |
0,178 |
0,61 |
|
Зимостойкость |
4,5 |
17,9* |
7,0 |
0,6 |
0,190 |
0,69 |
|
Вязкость водного экстракта |
3,1 |
2,7* |
1,8 |
0,8 |
0,250 |
0,54 |
|
Масса 1000 зерен |
1,6 |
36,1* |
43,4* |
1,1 |
0,206 |
0,31 |
|
Число падения |
1,4 |
7282* |
7996* |
1,1 |
0,213 |
0,37 |
Примечание. D, Hb Н2 и h2 — по B.I. Hayman (13). * Достоверно на 5 % уровне значимости.
Важным критерием служит величина коэффициента корреляции (r) между количественной характеристикой признака у гомозиготных линий per se и эффектами ОКС. В наших опытах он был высокодостоверным по всем изученным признакам, кроме урожайности, числа продуктивных стеблей на 1 м2 и числа зерен в колосе (см. табл. 2). Это создает трудности для селекции, так как по этим признакам невозможно раннее прогнозирование ОКС у инбредных линий и требуется интенсивное тестирование полученных на их основе гибридов Fi. Причина здесь в следующем: корреляция между собственной продуктивностью линий и их преимуществом в скрещиваниях зависит от того, какая часть дисперсии обусловлена аддитивными генами (14). В наших опытах доля вклада ОКС в вариансу признака урожайности оказалась низкой — в среднем 7,0 %. Следовательно, при использовании этого признака в качестве косвенного критерия для отбора вероятность получения высокогетерозисных гибридов на базе лучших по урожайности линий будет небольшой. В этом отношении рожь сходна с кукурузой, у которой продуктивность линий per se также не является надежным показателем их ОКС (15). Однако пренебрегать высокой урожайностью инбредных линий не следует, поскольку она имеет важное значение для их семеноводства.
Анализ параметров генетической изменчивости показал (табл. 3), что потенциал признаков продуктивности и качества зерна у ржи зависит от эффектов генных взаимодействий трех типов: аддитивного (когда эффекты суммируются по многим локусам), доминирования (внутрилокус-ное взаимодействие генов) и эпистаза (неаллельное взаимодействие генов). Однако их долевой вклад в дисперсию признаков сильно варьировал по годам и определялся тесным взаимодействием испытуемых генотипов с лимитирующими экологическими факторами (осадки, полегание), которые изменяли спектр эффективных генов, детерминирующих среднюю величину и генотипическую дисперсию признаков.
Отмеченное выше разделение изученных признаков на группы позволяет дифференцированно подойти к оценке их генетических систем и обоснованию методов селекционного улучшения.
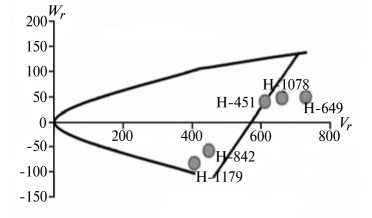
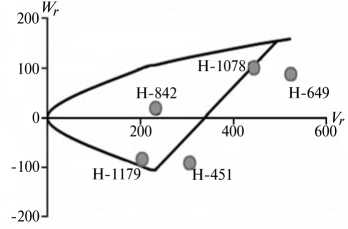
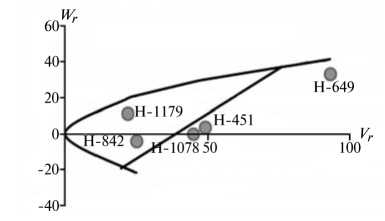
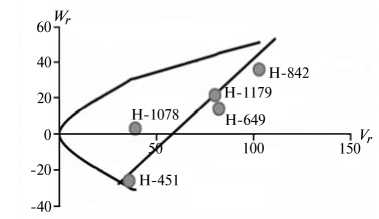
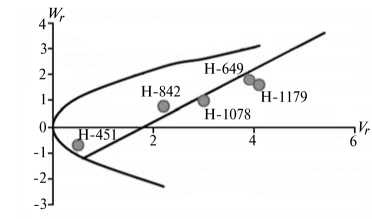
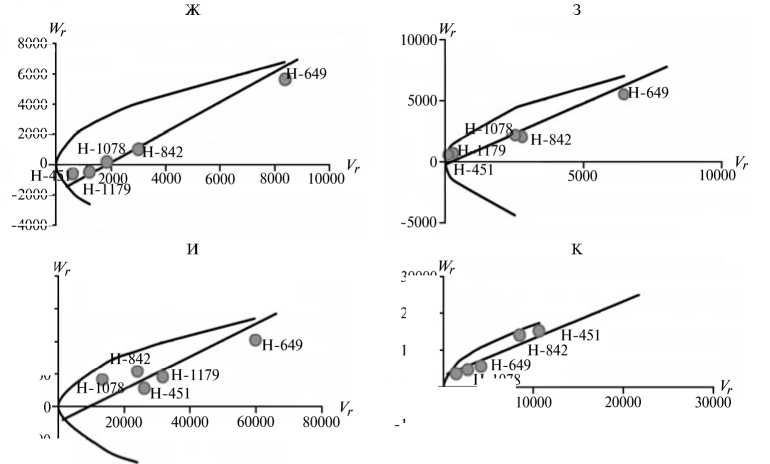
Отличительным свойством признаков из 1-й группы была сильная депрессия при инцухте, низкий вклад аддитивных эффектов генов в наследуемость, сравнительно высокий (за исключением крахмала) гипотетический гетерозис при межлинейных скрещиваниях (34,4-174,6 %) и точка пересечения линией регрессии оси W r , которая лежит ниже начала координат (рис. 1).
Экспрессия признаков этой группы сильно зависит от эффектов доминирования в локусах и от взаимодействия эпистатических генов. Достоверный эффект эпистаза проявился в оба года исследований по урожайности, высоте растений, числу зерен в колосе и содержанию крахмала. Надо полагать, он существенно влиял на компоненту доминирования, увеличивая потенциал неаддитивной генетической дисперсии, отражением которой стала высокая варианса по СКС.
По двум признакам 1-й группы (урожайность и высота растений) система генетического контроля оказалась относительно стабильной, коэффициенты корреляции r между суммой W r + Vr (коварианса + варианса) и средней величиной признака у родителей X p имели по годам стабильно отрицательный знак, что указывает на высокую экспрессию этих признаков под контролем доминантных генов. Однако по числу продуктивных стеблей на 1 м2 и числу зерен в колосе коэффициенты r за 2011 год были недостоверными, что свидетельствует о нестабильности генетической системы вследствие влияния погодных условий. Аналогичная особенность отмечена и по содержанию крахмала в 2012 году. Она проявилась в сильном взаимодействии генотип—среда, когда доминантные гены имели различную экспрессивность по причине различной устойчивости линий к полеганию. Поэтому селекционную оценку инбредных линий по указанным двум признакам целесообразно проводить через испытание в различных экологических условиях.
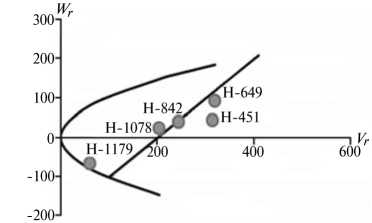
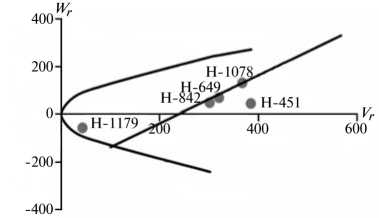
По сравнению с 1-й группой 2-я группа оказалась многочисленнее. Это вполне соответствует выводу H.H. Geiger (5) о том, что для большинства признаков озимой ржи аддитивная варианса гораздо больше, чем все другие компоненты генетической вариансы, вместе взятые. Характерная особенность признаков этой группы заключается в сравнительно высоком вкладе аддитивных генетических эффектов в наследуемость признака, слабой инбредной депрессии при инцухте и относительно низком (за исключением числа падения, высоты амилограммы и вязкости водного экстракта) гипотетическом гетерозисе при межлинейных скрещиваниях. Ни по одному из этих признаков стабильных эффектов эпистаза мы не выявили.
А
Б
В
Г
Д
Е
Ж
И
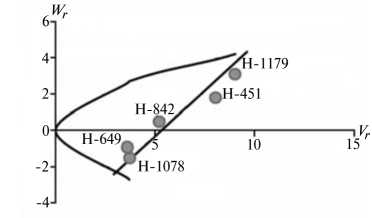
Рис. 1. Графики регрессии W r / V r (коварианса/варианса) у линий озимой ржи ( Secale cere-ale L.) Н-649, Н-1078, Н-1179, Н-451, Н-842 по признакам, для которых варианса общей комбинационной способности не превышает вариансу специфической комбинационной способности (1-я группа): А, Б — урожайность, В, Г — число продуктивных стеблей на 1 м2, Д, Е — число зерен в колосе, Ж, 3 — высота растений, И, К — содержание крахмала; слева и справа — соответственно 2011 и 2012 год (Московская обл.).
К
A
Б
8000л
80000л
60000-
40000-
6000-
4000-
2000-
-2000-
-4000-1
30000л
20000-
10000-
О-
-10000-
-20000-1
-20000J
-40000J
Н-1078
Н-1179 ,
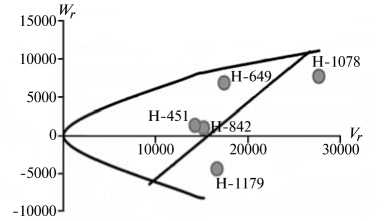
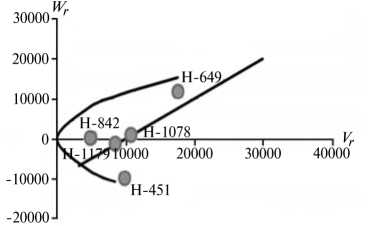
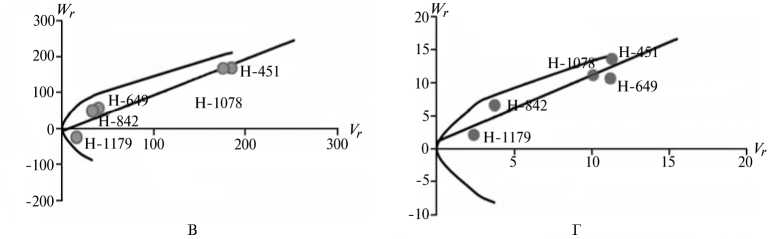
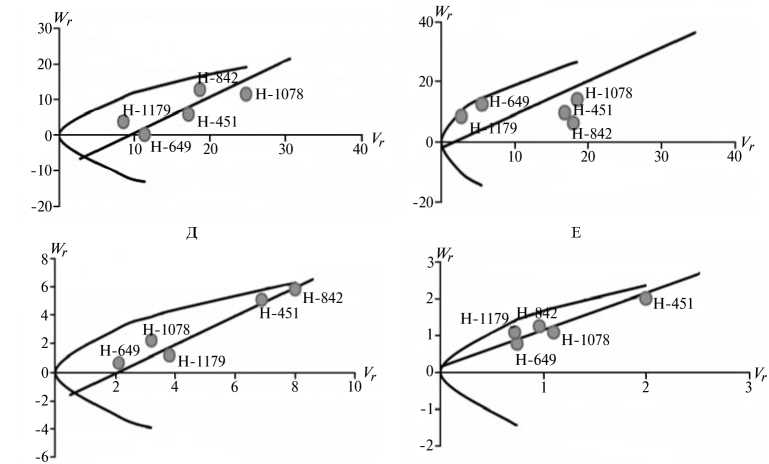
Рис. 2. Графики регрессии W r / V r (коварианса/варианса) у линий озимой ржи ( Secale cere-ale L.) Н-649, Н-1078, Н-1179, Н-451, Н-842 по признакам, для которых варианса общей комбинационной способности превышает вариансу специфической комбинационной способности (2-я группа): A, Б — зимостойкость, В, Г — масса 1000 зерен, Д, Е — вязкость водного экстракта, Ж, 3 — число падения, И, К — высота амилограммы; слева и справа — соответственно 2011 и 2012 год (Московская обл.).
20000-1
Преобладающую роль в их детерминации играют аддитивные эффекты генов, причем доминантные гены положительно увеличивали практически все признаки качества зерна, кроме содержания белка (рис. 2).
По шести признакам из этой группы (зимостойкость, масса 1000 зерен, натура зерна, число падения, вязкость водного экстракта, содержание белка) погодные условия в годы исследований мало влияли на систему генетического контроля. Она оказалась относительно стабильной, так как коэффициенты корреляции r между суммой W r + Vr и X p были достоверными и имели стабильный по годам знак, то есть направленность доминирования не изменилась. Однако по шести другим признакам (высота амилограммы, температура клейстеризации крахмала, отношение H/D, объем формового хлеба, а также поражение снежной плесенью и бурой ржавчиной) проявилось сильное взаимодействие генотип-среда. На экологическую неустойчивость генетических систем для этих признаков указывали коэффициенты r между X p и Wr + Vr , которые в разные годы были недостоверными, а также генетические параметры и точки расположения линий на графике Хеймана, которые по годам значительно различались. Это свидетельствует о лабильности указанных генетических систем под влиянием лимитирующих факторов среды. Можно полагать, что генетическая информация по этим признакам реализуется в таком взаимодействии со средой, при котором обе причины, их определяющие, становятся трудноотделимы друг от друга. В силу этого экспрессия указанных признаков в разные годы может определяться как доминантными, так и рецессивными генами, что усложняет работу по их селекционному улучшению.
Таким образом, для более эффективного улучшения низконасле-дуемых признаков (1-я группа) необходимо максимально задействовать методы гетерозисной селекции, отдавая приоритет отбору инбредных линий с высокой специфической комбинационной способностью (СКС) по урожайности и низкой СКС по высоте растений. Селекционное улучшение инбредных линий по высоконаследуемым признакам (2-я группа) целесообразно проводить по принципу кумулятивного накопления в них ценных генов методом рекуррентного отбора. Особенно важно придерживаться этого принципа при селекции инбредных линий на зимостойкость, так как гипотетический гетерозис по этому признаку в межлинейных скрещиваниях проявляется слабо. Стратегия селекции линий по экологически неустойчивым признакам, склонным к переопределению своей генетической формулы под влиянием средовых факторов (высота амилограммы, температура клейстеризации крахмала, формоустойчивость теста, объем хлеба, поражение болезнями), должна базироваться на раннем прогнозировании общей комбинационной способности родительских форм по степени фенотипической выраженности у них этих признаков.
Л И Т Е Р А Т У Р А
-
1. Geiger H.H. Hybrid breeding in rye. Proc. of the EUCARPIA Rye Meeting. Svalov, Sweden, 1985: 237-265.
-
2. Geiger H.H., Miedaner T. Rye breeding. In: Handbook Cereals /M.J. Carena (ed.). Springer Science + Business Media, LLC, 2009: 157-182.
-
3. Karpenstein-Machan M., Maschka R. Progress in rye breeding. Vortr. Pflan-zenzuchtung, 1996, 35: 7-13.
-
4. Geiger H.H. Wege, Fortschritte und Aussichten der Hybridzuchtung. In: Pflanzenproduction in Wandel. VCH Verlag, Weinheim, 1990: 41-72.
-
5. Geiger H.H. Breeding methods in diploid rye ( Secale cereale L.). Tag.-Ber. Akad. Land-wirtsch.-Wiss. DDR, Berlin, 1982, 198: 305-332.
-
6. Wilde P., Menzel J., Schmiedchen B. Estimation of general and specific combining
ability variances and their implications on hybrid rye breeding. Plant Breeding and Seed Science, 2003, 47(1/2): 89-98.
-
7. Geiger H.H. Z u chtung. In: Roggen. Anbau — Verarbeitung — Markt /W. Seibel, W. Steller (eds.). Behr’s Verlag, Hamburg, 1988: 25-43.
-
8. Kolasinska I., Wegrzyn S. Combining ability for selected characters in winter rye. Proc. of the EUCARPIA Rye Meeting. Radzikow, Poland, 2001: 91-96.
-
9. Smialowski Ò., Wegrzyn S. The genetic and statistical analysis of the heritability of important traits in winter rye ( Secale cereale L.). Biuletyn IHAR, 2003, 230: 205-214.
-
10. Smialowski Ò., Wegrzyn S. The influence of environments on the epistatic effects of genes controlling some traits in winter rye. Proc. of the EUCARPIA Rye Meeting, Radzikow, Poland, 2001: 105-117.
-
11. Geiger H.H. Epistasis and heterosis. Proc. Second Int. Conf. on Quantitative Genetics (31 May-5 June 1987) /B.S. Weir (ed.). Raleigh. NC. Sinauer Assoc. Inc., Sunderland MA, USA, 1987: 395-399.
-
12. Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems. Austral. J. Biol. Sci., 1956, 9: 463-493.
-
13. Hayman B.I. The theory and analysis of diallel crosses. Genetics, 1954, 39: 789-809.
-
14. Ôîëêîíåð Ä.Ñ. Ââåäåíèå â ãåíåòèêó êîëè÷åñòâåííûõ ïðèçíàêîâ. Ì., 1985.
-
15. Gama E.E.G., Hallauer A.R. Relation between inbred and hybrid traits in maize. Crop Sci., 1977, 17: 703-706.
ГНУ Московский НИИ сельского хозяйства Поступила в редакцию
«Немчиновка» Россельхозакадемии, 5 марта 2013 года
143026 Ðîññèÿ, Ìîñêîâñêàÿ îáë., Îäèíöîâñêèé ð-í, ãï Íîâîèâàíîâñêîå,
Sel’skokhozyaistvennaya biologiya [ Agricultural Biology ], 2015, V. 50, № 1, pp. 75-84
GENETIC RESEARCH OF QUANTITATIVE TRAITS OF INBRED LINES OF WINTER RYE ( Secale cereale L.) IN DIALLEL CROSSINGS
A.A. Goncharenko, S.V. Krahmalev, A.V. Makarov, S.A. Yermakov
Список литературы Генетический анализ количественных признаков у инбредных линий озимой ржи ( Secale cereale L.) в диаллельных скрещиваниях
- Geiger H.H. Hybrid breeding in rye. Proc. of the EUCARPIA Rye Meeting. Svalov, Sweden, 1985: 237-265.
- Geiger H.H., Miedaner T. Rye breeding. In: Handbook Cereals/M.J. Carena (ed.). Springer Science + Business Media, LLC, 2009: 157-182.
- Karpenstein-Machan M., Maschka R. Progress in rye breeding. Vortr. Pflanzenzuchtung, 1996, 35: 7-13.
- Geiger H.H. Wege, Fortschritte und Aussichten der Hybridzuchtung. In: Pflanzenproduction in Wandel. VCH Verlag, Weinheim, 1990: 41-72.
- Geiger H.H. Breeding methods in diploid rye (Secale cereale L.). Tag.-Ber. Akad. Landwirtsch.-Wiss. DDR, Berlin, 1982, 198: 305-332.
- Wilde P., Menzel J., Schmiedchen B. Estimation of general and specific combining ability variances and their implications on hybrid rye breeding. Plant Breeding and Seed Science, 2003, 47(1/2): 89-98.
- Geiger H.H. Züchtung. In: Roggen. Anbau -Verarbeitung -Markt/W. Seibel, W. Steller (eds.). Behr’s Verlag, Hamburg, 1988: 25-43.
- Kolasinska I., Wegrzyn S. Combining ability for selected characters in winter rye. Proc. of the EUCARPIA Rye Meeting. Radzikow, Poland, 2001: 91-96.
- Smialowski Т., Wegrzyn S. The genetic and statistical analysis of the heritability of important traits in winter rye (Secale cereale L.). Biuletyn IHAR, 2003, 230: 205-214.
- Smialowski Т., Wegrzyn S. The influence of environments on the epistatic effects of genes controlling some traits in winter rye. Proc. of the EUCARPIA Rye Meeting, Radzikow, Poland, 2001: 105-117.
- Geiger H.H. Epistasis and heterosis. Proc. Second Int. Conf. on Quantitative Genetics (31 May-5 June 1987)/B.S. Weir (ed.). Raleigh. NC. Sinauer Assoc. Inc., Sunderland MA, USA, 1987: 395-399.
- Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems. Austral. J. Biol. Sci., 1956, 9: 463-493.
- Hayman B.I. The theory and analysis of diallel crosses. Genetics, 1954, 39: 789-809.
- Фолконер Д.С. Введение в генетику количественных признаков. М., 1985.
- Gama E.E.G., Hallauer A.R. Relation between inbred and hybrid traits in maize. Crop Sci., 1977, 17: 703-706.
