Генетическое разнообразие пород степного скота в России и их вклад в изменчивость аллелофонда крупного рогатого скота

Автор: Абдельманова А.С., Абрамов Г.О., Волкова В.В., Зиновьева Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Локальные породы уступают по продуктивности специализированным коммерческим породам скота, однако обладают хорошими адаптационными качествами. Степная зона России занимает обширные пространства, характеризуясь резко-континентальным климатом. Разведение высокопродуктивных коммерческих пород в условиях степной зоны требует значительных затрат, в то время как локальные породы менее требовательны к условиям содержания и кормления. Полногеномное генотипирование с применением SNP-маркеров помогает детально охарактеризовать генетическое разнообразие и структуру аллелофонда пород. Учитывая особенности таких пород, важно комплексно подходить к разработке программ сохранения их генетических ресурсов, включая использование молекулярно-генетического мониторинга для оценки структуры популяций и контроля инбридинга. В настоящей работе впервые приведены результаты анализа аллелофондов степных пород крупного рогатого скота разного происхождения (группы турано-монгольского, серого и красного скота) в сравнительном аспекте с улучшающими и предковыми породами. Описаны аллельные профили степных пород скота, а также профили улучшающих и предковых пород для исследуемых популяций, выявлены аллели, общие для степных пород скота. Целью исследований было изучение генетического разнообразия, филогенетических взаимосвязей и изменений аллелофонда локальных степных пород крупного рогатого скота с использованием микросателлитных маркеров. Объектом исследования были образцы биоматериала от животных трех пород крупного рогатого различного происхождения, традиционно разводимых в степной зоне: красная степная (REDSTEP, n = 161), калмыцкая (KALM, n = 38), серая украинская (GUKR, n = 150). Образцы от серой украинской породы были разделены на три субпопуляции в соответствии с происхождением, а именно архивные образцы от животных, разводимых в 2008 году в монастырском подсобном хозяйстве в Киевской области (GUKR, n = 42); образцы от животных, содержащихся в 2009-2010 годах в Новосибирской области (GUKR_SYB, n = 45); образцы от животных, содержащихся в 2023 году в Алтайском крае (GUKR_ALT, n = 63). В качестве групп сравнения в анализ были включены образцы от трех трансграничных пород, используемых для улучшения локального скота: голштинской (HOL, n = 34), симментальской (SIM, n = 32), герефордской (HRFD, n = 26). Также использовали образцы двух групп локального скота, разводимого в условиях степи на протяжении многих веков, — киргизского (KRGZ, n = 20) и монгольского (MNGL, n = 76). Породы генотипировали по 11 микросателлитным локусам: TGLA227, BM2113, TGLA53, ETH10, SPS115, TGLA122, INRA23, TGLA126, BM1818, ETH225, BM1824. Для оценки генетического разнообразия в каждой породе были рассчитаны наблюдаемая (HO) и несмещенная ожидаемая гетерозиготность (UHE), коэффициент инбридинга (UFIS), основанный на несмещенной ожидаемой гетерозиготности, и аллельное разнообразие, скорректированное на размер выборки (AR). Степень генетической дифференциации оценивали на основании попарных значений генетических дистанций Jost’s D. Матрицы попарных значений UFST и генетических дистанций Jost’s D использовали для построения дендрограмм по алгоритму «сеть соседей» в программе SplitsTree 4.14.5 software. Анализ структуры популяций проводили в программе Structure 2.3.4 с визуализацией результатов в программной среде R с использованием пакета BITE. Программа CLUMPAK использовалась для определения наиболее вероятного числа кластеров в исследованной выборке на основании значений DeltaK. Кроме того, при определении структуры популяции дополнительно учитывали полученные в CLUMPAK средние оценки сходства нескольких независимых запусков при одном значении K. На основании анализа РСА была выявлена дифференциация красной степной, голштинской и одной из популяций серой украинской породы от остальных групп. Анализ генетических дистанций ' выявил минимальные расстояния внутри группы турано-монгольского скота (Jost’s D = 0,042). Показана близость между традиционно разводимым в степи скотом и серой украинской (Jost’s D = 0,088), а также красной степной (Jost’s D = 0,124) породами. По результатам анализа генетического разнообразия установлены минимальные значения наблюдаемой гетерозиготности HO и аллельного разнообразия AR в серой украинской породе (HO = 0,593-0,736 и AR = 4,211-6,250), в то время как в группе турано-монгольского скота они оказались несколько выше (HO = 0,715-0,841 и AR = 7,436-8,000). Сопоставление аллельных профилей популяций выявило аллели (81 в локусе TGLA227, 115 в TGLA126), общие для всех степных пород скота. Также обнаружено совпадение аллельных профилей для пар пород: 5 из 11 локусов — для KALM и REDSTEP, 7 из 11 локусов — для KALM и GUKR, 9 из 11 — для REDSTEP и GUKR. Анализ структуры популяций выявил генетические компоненты, характерные для каждой из исследованных пород, а также предковые компоненты. Исследованные породы степного скота являются носителями уникальных форм изменчивости.
Степные породы, крупный рогатый скот, молекулярно-генетические маркеры, аллелофонд, популяция
Короткий адрес: https://sciup.org/142247686
IDR: 142247686 | УДК: 636.2:575.174.015.3:577.21 | DOI: 10.15389/agrobiology.2026.2.245rus
Genetic diversity of steppe breeds bred in Russia and their impact in variability of allele pool of cattle
Local breeds are inferior in productivity to specialized commercial cattle breeds, but possess good adaptability. The Russian steppe zone occupies vast areas, characterized by a sharply continental climate. Breeding highly productive commercial breeds in the steppe zone requires significant investment, while local breeds are less demanding in terms of housing and feeding conditions. Whole-genome genotyping using SNP markers helps to thoroughly characterize the genetic diversity and allele pool structure of breeds. Given the characteristics of such breeds, it is important to take a comprehensive approach to developing programs for preserving their genetic resources, including the use of molecular genetic monitoring to assess population structure and control inbreeding. This paper presents the first analysis of the allele pools of steppe cattle breeds of different origins (Turanian-Mongolian, Gray, and Red cattle groups) in a comparative study with improving and ancestral breeds. The allelic profiles of steppe cattle breeds, as well as the profiles of improving and ancestral breeds for the studied populations are described; alleles common to steppe cattle breeds are identified. The aim of the research was to investigate the genetic diversity, phylogenetic relationships and changes in the allele pool of local steppe cattle breeds using microsatellite markers. The object of the study were samples of three cattle breeds of different origins, traditionally bred in the steppe zone: Red Steppe (REDSTEP, n = 161), Kalmyk (KALM, n = 38), and Gray Ukrainian (GUKR, n = 150). Samples of the Gray Ukrainian breed were divided into three subpopulations according to their origin, namely, archival samples of animals bred in 2008 at a monastery subsidiary farm in the Kyiv region (GUKR, n = 42); Samples of animals kept in 2009-2010 in the Novosibirsk region (GUKR_SYB, n = 45); samples of animals kept in 2023 in the Altai region (GUKR_ALT, n = 63). Samples of three transboundary breeds used to improve local cattle were involved as comparison groups, these were Holstein (HOL, n = 34), Simmental (SIM, n = 32), Hereford (HRFD, n = 26) cattle. Samples of two groups of local cattle bred in steppe conditions for many centuries also used were Kyrgyz (KRGZ, n = 20) and Mongolian (MNGL, n = 76) cattle. The breeds were genotyped for 11 microsatellite loci TGLA227, BM2113, TGLA53, ETH10, SPS115, TGLA122, INRA23, TGLA126, BM1818, ETH225, and BM1824. To assess the genetic diversity in each breed, the observed (HO) and unbiased expected heterozygosity (UHE), the inbreeding coefficient (UFIS) based on the unbiased expected heterozygosity, and the allelic diversity adjusted for sample size (AR) were calculated. The degree of genetic differentiation was estimated based on pairwise Jost’s D genetic distances. Matrices of pairwise FST values and Jost’s D genetic distances were used to construct dendrograms using the neighbor-joining network algorithm in SplitsTree 4.14.5 software. Population structure analysis was performed in Structure 2.3.4 with results visualization in the R software environment using the BITE package. CLUMPAK was used to determine the most probable number of clusters in the studied sample based on DeltaK values. In addition, the average similarity estimates obtained in CLUMPAK for several independent runs with the same K value were additionally accounted when determining the population structure. Based on PCA analysis, differentiation of the Red Steppe, Holstein, and one of the populations of the Gray Ukrainian breed from the other groups was revealed. Analysis of the Jost's D genetic distances revealed minimal distances within the Turanian-Mongolian cattle group (Jost’s D = 0.042). The closeness between cattle traditionally bred in the steppe and the Grey Ukrainian (Jost’s D = 0.088) and Red Steppe (Jost’s D = 0.124) breeds was shown. Based on the genetic diversity data, the minimum values of the observed heterozygosity Ho and allelic diversity Ar were revealed in the Grey Ukrainian breed (HO = 0.593-0.736 and AR = 4.211-6.250), while in the Turanian-Mongolian cattle group they were slightly higher (HO = 0.715-0.841 and AR = 7.436-8.000). Comparison of the allelic profiles of the populations revealed alleles (81 in the TGLA227 locus, 115 in TGLA126) common to all steppe cattle breeds. A match in the allelic profiles for pairs of breeds was also found: 5 of 11 loci for KALM and REDSTEP, 7 of 11 loci for KALM and GUKR, and 9 of 11 for REDSTEP and GUKR. An analysis of the population structure revealed genetic components characteristic of each of the studied breeds, as well as ancestral components. The studied steppe cattle breeds are carriers of unique forms of variability. A strategy for further conservation of the genetic resources of these breeds may involve monitoring the genetic state of populations and preventing mass crossbreeding with transboundary breeds.
Текст научной статьи Генетическое разнообразие пород степного скота в России и их вклад в изменчивость аллелофонда крупного рогатого скота
Интенсификация и глобализация животноводства привели к вытеснению локальных аборигенных пород высокопродуктивными коммерческими (1). Породы крупного рогатого скота, разводимые в настоящее время в степной зоне Российской Федерации, характеризуются уникальным алле-лофондом, формировавшимся в течение долгого времени под влиянием суровых природно-климатических условий и преимущественно кочевого образа жизни народов, когда-то населявших Великую степь. Специфические комбинации генных вариантов у степных пород способствуют устойчивости к экстремальным температурным режимам, длительной засухе, а также обусловливают иммунитет к различным патогенам и инвазиям (2).
Полногеномное генотипирование с применением SNP-маркеров помогает детально охарактеризовать генетическое разнообразие и структуру аллелофонда пород. Например, при анализе российских молочных пород черно-пестрого скота выявлена значительная интрогрессия голштинской породы, влияющая на уникальность генетического фонда местных животных (3). В связи с тем, что многие локальные породы скота нашей страны, в том числе степные, находятся в условиях малочисленности и/или изоляции, в последние десятилетия возрос интерес к сохранению аллелофонда локальных пород. Учитывая особенности таких пород, важно комплексно подходить к разработке программ сохранения их генетических ресурсов, включая использование молекулярно-генетического мониторинга для оценки структуры популяций и контроля инбридинга (4, 5).
Евразийская степь, также известная как Великая степь, представляет собой огромные территории с разным климатом, от умеренного до резкоконтинентального, которые протянулись от Восточной Европы через Причерноморье и Казахстан до Дальнего Востока и характеризуются безлесными пространствами, покрытыми травянистой растительностью. Великая степь играла ключевую роль в истории человечества как колыбель кочевых цивилизаций. Степная зона России занимает обширные пространства на юге европейской части и на юге Западной Сибири. Она характеризуется резко-континентальным климатом с жарким и засушливым летом и холодной, почти бесснежной зимой, что создает экстремальные условия для обитающих здесь животных (6). Коммерческие породы, такие как голштинская и симментальская, завезенные в степные зоны России, обеспечивают высокую продуктивность, однако требуют больше затрат и внимания в содержании и в условиях степей могут испытывать стресс, что снижает продуктивность и жизнеспособность (7).
В свою очередь, следует отметить существенное различие климатических зон степей в России. Климат Алтая и южной части страны различается, что служит важным фактором для адаптации пород. Южная часть России (Приазовье, Причерноморье, Северный Кавказ) характеризуется более теплым и мягким климатом с жарким длинным летом и мягкой зимой, что обеспечивает более богатую кормовую базу, в отличие от Алтайского региона. В связи с этими климатическими особенностями породы крупного рогатого скота (КРС) демонстрируют разную степень адаптации.
Исследуемые породы можно разделить на две группы: европейский тауринный скот (герефордская, серая украинская, красная степная, голштинская, симментальская) и турано-монгольский скот (монгольская, киргизская, калмыцкая). Многие породы турано-монгольского скота проявляют большую выносливость в результате адаптации к суровому климату азиатских степей. Так, животные калмыцкой породы без ущерба для здоровья переносят продолжительные морозы (до -40 °С и ниже) и холодные ветры зимой, и жару до 45 °С летом. У скота этой породы прекрасно выражен физиологический гомеостаз (способность организма сохранять внутреннюю среду при различных изменениях внешней среды) (7).
Генетические исследования турано-монгольского скота с использованием разных типов маркеров, проводившиеся на протяжении последних 15 лет, подтвердили общность происхождения степных пород (9, 10). Совместное изучение европейских и турано-монгольских пород показало, что между ними сохраняется генетическое разделение, отражающее особенности их происхождения и филогенеза. Однако были выявлены и следы генетического обмена (11, 12). Например, у калмыцкой породы в генофонде присутствуют аллели, присущие европейским породам. Возможно, это связано с прилитием крови шортгорнской породы еще в период существования СССР (5).
На большей части южных территорий страны наиболее распространена красная степная порода и ее помеси разной кровности. Им отдается предпочтение, благодаря приспособленности к засушливому климату степной зоны, устойчивости к заболеваниям, хорошей воспроизводительной способности и продолжительному хозяйственному использованию. При разведении помесного скота изменение генофонда затронуло и красную степную породу, в которой все чаще стали встречаться аллели, свойственные голштинской породе (13).
Серая украинская порода принадлежит к ветви так называемого серого степного скота (также именуемого подольским по названию региона возможного происхождения — Подолье), куда, помимо нее, относят ряд серых пород скота Балканского и Итальянского полуостровов (14). Показано, что, несмотря на схожесть фенотипов (серая шерсть, палевая пигментация теленка при рождении, большие рога), генетически серая украинская отличается от южно-европейских пород (15).
В настоящей работе впервые приведены результаты анализа аллело-фондов степных пород крупного рогатого скота разного происхождения (группы турано-монгольского, серого и красного скота) в сравнительном аспекте с улучшающими и предковыми породами. Описаны аллельные профили степных пород скота, а также профили улучшающих и предковых пород для исследуемых популяций, выявлены аллели, общие для степных пород скота.
Нашей целью было изучение генетического разнообразия, филогенетических взаимосвязей и изменений аллелофонда локальных степных пород крупного рогатого скота с использованием микросателлитных маркеров.
Методика. Работу проводили в течение 2024-2025 годов. Материалом служили образцы (ушной выщип, кровь, сперма) восьми пород КРС, сохраняемые в биоресурсной коллекции «Банк генетического материала домашних и диких видов животных и птицы» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста.
Объектом исследования были три породы крупного рогатого различного происхождения, традиционно разводимого в степной зоне: красная степная (REDSTEP, n = 161), калмыцкая (KALM, n = 38), серая украинская (GUKR, n = 150).
Предыдущие исследования (16) показали значительную генетическую дифференциацию серой украинской породы в зависимости от региона и времени разведения, поэтому в настоящей работе образцы серой украинской породы были разделены на три субпопуляции в соответствии с происхождением, а именно архивные образцы от животных, разводимых в 2008 году в монастырском подсобном хозяйстве в Киевской области (GUKR, n = 42); образцы от животных, содержавшихся в 2009-2010 годах в Новосибирской области (GUKR_SYB, n = 45); образцы от животных, содержавшихся в 2023 году в Алтайском крае (GUKR_ALT, n = 63).
В качестве групп сравнения в анализ были включены образцы от трех трансграничных пород, используемых для улучшения локального скота, — голштинской (HOL, n = 34), симментальской (SIM, n = 32), гере-фордской (HRFD, n = 26). Также использовали образцы двух групп локального скота, разводимых в условиях степи на протяжении многих веков и, вероятно, родственных современной калмыцкой породе, — киргизского (KRGZ, n = 20) и монгольского (MNGL, n = 76) скота.
ДНК из образцов выделяли с использованием коммерческих наборов «ДНК-Экстран» (ЗАО «Синтол», Россия) согласно протоколу фирмы-изготовителя. Качество ДНК оценивали посредством измерения концентрации на флуориметре Qubit 3.0 («Thermo Fisher Scientific», США) (не менее 1 нг/мкл) и установления соотношения А260/А280 на спектрофотометре NanоDrop8000 («Thermo Fisher Scientific», США) (в интервале 1,7-2,0). Породы были генотипированы по 11 микросателлитным локусам, рекомендованным Международным обществом генетики животных (International Society for Animal Genetics, ISAG) (17): TGLA227, BM2113, TGLA53, ETH10, SPS115, TGLA122, INRA23, TGLA126, BM1818, ETH225, BM1824. Амплификацию и анализ длин фрагментов проводили, как описано ранее (18). ПЦР проводили в конечном объеме 10 мкл в ПЦР буфере с 200 мМ dNTP, 1,0 мМ MgCl2, 0,5 мМ смеси праймеров (последовательности приведены на , 1 ед. Taq-полимеразы (ООО «Диалат», Россия) и 1 мкл геномной ДНК. Амплификацию выполняли при следующем температурно-временном режиме: 4 мин при 95 °С (начальная денатурация); 20 с при 95 °С, 30 с при 63 °С, 1 мин при 72 (35 циклов) °С; 10 мин при 72 °С (завершающая элонгация). Анализ длин фрагментов осуществляли с использованием генетического анализатора ABI 3130xl («Applied Biosystems, Inc.», США). Размеры аллелей определяли с помощью программного обеспечения GeneMapper v.4.0 («Applied Biosystems, Inc.», США).
Общее число аллелей и приватных аллелей рассчитывали с использованием программного обеспечения GenAIEx 6.503 (19), средние значения параметров генетического разнообразия для каждой группы, стандартные ошибки и 95 % доверительный интервал определяли с помощью R-пакета diveRsity (20). Для оценки генетического разнообразия в каждой породе были рассчитаны наблюдаемая (Ho) и несмещенная ожидаемая гетерозиготность (uHe) (21), коэффициент инбридинга (uFis) (22), основанный на несмещенной ожидаемой гетерозиготности, и аллельное разнообразие, скорректированное на размер выборки (Ar) (23). Степень генетической дифференциации изучаемых пород оценивали на основании попарных значений генетических дистанций Jost’s D (24). Матрицы попарных значений Fst и генетических дистанций Jost’s D использовали для построения дендрограмм по алгоритму «сеть соседей» в программе SplitsTree 4.14.5 software (25). Анализ структуры популяций проводили в программе Structure 2.3.4 (26) с визуализацией результатов в программной среде R с использованием пакета BITE (27). Программа CLUMPAK (28, 29) использовалась для определения наиболее вероятного числа кластеров в исследованной выборке на основании значений DeltaK по методам, предложенным G. Evanno с соавт. (30) и J.K. Pritchard с соавт. (26). Кроме того, при определении структуры популяции дополнительно учитывали полученные в CLUMPAK средние оценки сходства нескольких независимых запусков при одном значении K.
Результаты. В таблице 1 приведена сводная информация о статусе пород, объеме выборок и основаниях для включения в настоящее исследование.
1. Характеристика исследуемых популяций крупного рогатого скота степных пород (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы)
|
Обозначение |
Число образцов |
Порода |
Статус породы |
Группа скота по происхождению |
Причина включения в анализ |
|
KALM |
38 |
Калмыцкая |
Локальная |
Турано-монгольский скот |
Исследуемая порода |
|
GUKR |
42 |
Серая украинская Локальная |
Серый скот |
Исследуемая порода |
|
|
GUKR_SYB |
45 |
Серая украинская Локальная |
Серый скот |
Исследуемая порода |
|
|
GUKR_ALT |
63 |
Серая украинская Локальная |
Серый скот |
Исследуемая порода |
|
|
REDSTEP |
161 |
Красная степная |
Локальная |
Красный скот |
Исследуемая порода |
|
HOL |
34 |
Голштинская |
Трансграничная |
Черно-пестрый скот |
Улучшающая порода |
|
SIM |
32 |
Симментальская |
Трансграничная |
Палево-пестрый скот Улучшающая порода |
|
|
HRFD |
26 |
Герефордская |
Трансграничная |
Красный скот |
Улучшающая порода |
|
KRGZ |
20 |
Киргизский скот |
Локальная |
Турано-монгольский скот |
Традиционно разводится в условиях степи |
|
MNGL |
76 |
Монгольский скот |
Локальная |
Турано-монгольский скот |
Традиционно разводится в условиях степи |
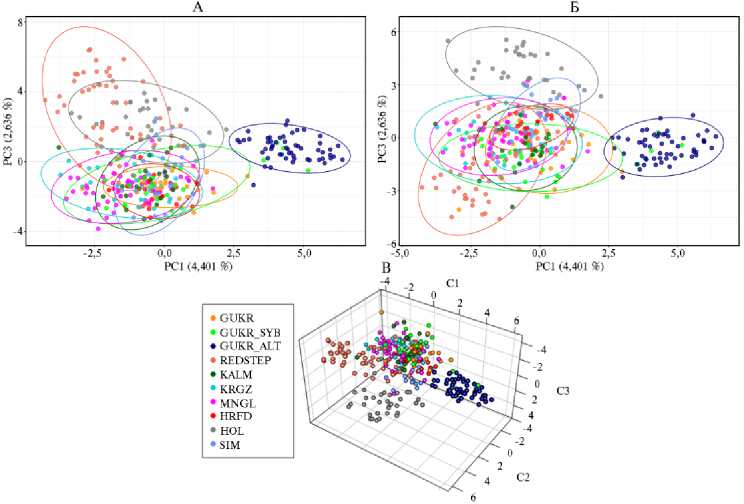
Рис. 1. Анализ генетической дифференциации исследуемых популяций крупного рогатого скота степных пород на основании анализа главных компонент: А — пространство 1 и 2 главных компонент, Б — пространство 1 и 3 главных компонент, В — 3D-проекция локализации образцов в пространстве первых трех компонент; GUKR — серая украинская, GUKR_SYB — серая украинская (г. Новосибирск), GUKR_ALT — серая украинская (Алтай), REDSTEP — красная степная, KALM — калмыцкая, KRGZ — киргизский скот, MNGL — монгольский скот, HRFD — герефордская, HOL — голштинская, SIM — симментальская (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы).
На основании генотипов по 11 микросателлитным локусам была проведена оценка филогенетических взаимосвязей и дифференциации исследуемых пород (рис. 1).
В исследуемой выборке выделились четыре основных кластера: один был образован преимущественно образцами красной степной породы, второй — голштинской породы, третий — серой украинской породы, разводимой в Алтайском крае, в четвертый, наиболее крупный кластер вошли образцы от остальных исследуемых пород (см. рис. 1).
Первая компонента, отвечающая за 4,401 % генетической изменчивости, отделила алтайскую популяцию серой украинской породы от остальных. Вторая компонента, описывающая 3,298 % генетической изменчивости, дифференцировала голштинскую и красную степную породы от основной группы. Формирование перекрывающегося массива значительным числом образцов разных пород может объясняться следующими причинами. Популяции турано-монгольского скота (KALM, KRGZ, MNGL) имеют общее происхождение, а также могли на определенном этапе развития улучшаться герефордской породой для повышения мясных качеств, архивные образцы серой украинской породы (GUKR, GUKR_SYB) могли улучшаться другими породами (симментальской и/или герефордской).
На основании 11 микросателлитных локусов были определены генетические дистанции посредством расчета значений Jost’s D (табл. 2).
2. Генетические дистанции между изученными популяциями крупного рогатого скота степных пород на основании 11 STR-локусов (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы)
Алтайская популяция серой украинской породы (GUKR_ALT) показала наибольшую генетическую удаленность от других групп (Jost’s D = 0,2130,384) (см. табл. 2). Интересно отметить, что для киргизского скота и монастырской популяции серой украинской породы (GUKR) выявлены минимальные дистанции (Jost’s D = 0,088). Наименьшие генетические дистанции выявлены между калмыцкой породой, киргизским (Jost’s D = 0,042) и монгольским (Jost’s D = 0,041) скотом, что подтверждает общность происхождения этих групп скота. Наиболее удалены от калмыцкой породы голштинская (Jost’s D = 0,216) и алтайская популяция серой украинской породы (Jost’s D = 0,252). Красная степная порода была наиболее близка к киргизскому (Jost’s D = 0,130) и монгольскому (Jost’s D = 0,124) скоту.
Для наилучшего понимания взаимных связей между исследуемыми популяциями генетические дистанции Jost’s D были визуализированы в форме дендрограммы, построенной по алгоритму Neighbour-Net (рис. 2).
Как видно из дендрограммы (см. рис. 2), исследуемые популяции образовали породоспецифичные ветви, причем калмыцкая порода, монгольский и киргизский скот оказались локализованы вблизи друг от друга, что свидетельствует в пользу общих исторических корней. Интересно отме- тить, что степные породы (архивная серая украинская и красная степная) располагались недалеко от группы пород, традиционно разводимых в условиях степи.
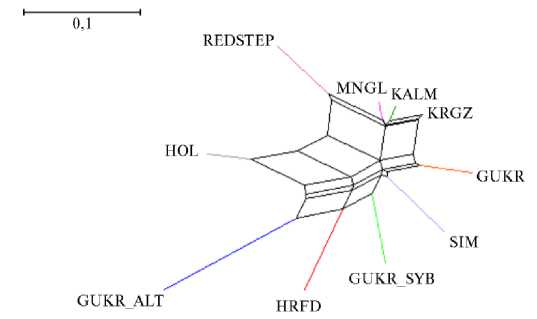
Рис. 2. Дендрограмма взаимных связей между исследуемыми популяциями крупного рогатого скота степных пород, построенная на основании генетических дистанций по Jost’s D с использованием алгоритма Neighbour-Net: GUKR — серая украинская, GUKR_SYB — серая украинская (г. Новосибирск), GUKR_ALT — серая украинская (Алтай), REDSTEP — красная степная, KALM — калмыцкая, KRGZ — киргизский скот, MNGL — монгольский скот, HRFD — ге-рефордская, HOL — голштинская, SIM — симментальская (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы).
Анализ генетического разнообразия (табл. 3) показал, что значения наблюдаемой гетерозиготности и аллельного разнообразия сильно варьировали в популяциях серой украинской породы разных лет и локаций разведения (H o = 0,593-0,736 и A r = 4,211-6,250).
-
3. Характеристика генетического разнообразия исследуемых популяций крупного рогатого скота степных пород по STR-маркерам (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы)
-
4. Аллельное разнообразие (число аллелей) в исследованных популяциях крупного рогатого скота степных пород по STR-маркерам (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы)
|
Популяция n |
H o ( M ±SEM) |
u H e ( M ±SEM) |
A r ( M ±SEM) |
U F IS (ДИ 95 %) |
|
|
GUKR |
43 |
0,736±0,034 |
0,736±0,031 |
6,250±0,500 |
- 0,006 ( - 0,086; 0,074) |
|
GUKR_SYB |
44 |
0,657±0,043 |
0,696±0,027 |
5,778±0,355 |
0,052 ( - 0,053; 0,157) |
|
GUKR_ALT |
63 |
0,593±0,045 |
0,577±0,040 |
4,211±0,308 |
- 0,029 ( - 0,084; 0,026) |
|
REDSTEP |
161 |
0,718±0,041 |
0,731±0,034 |
5,995±0,424 |
0,016 ( - 0,048; 0,080) |
|
KALM |
38 |
0,715±0,046 |
0,764±0,032 |
7,436±0,745 |
0,063 ( - 0,027; 0,153) |
|
KRGZ |
20 |
0,841±0,021 |
0,778±0,024 |
8,000±0,661 |
- 0,085 ( - 0,137; -0,033) |
|
MNGL |
76 |
0,731±0,024 |
0,767±0,022 |
7,552±0,794 |
0,043 ( - 0,015; 0,101) |
|
HRFD |
26 |
0,668±0,064 |
0,653±0,053 |
4,990±0,452 |
- 0,005 (-0,078; 0,068) |
|
HOL |
34 |
0,690±0,040 |
0,682±0,036 |
5,127±0,457 |
- 0,016 ( - 0,098; 0,066) |
|
SIM |
32 |
0,744±0,036 |
0,679±0,033 |
5,711±0,535 |
- 0,102 ( - 0,167; - 0,037) |
|
Примечание. |
GUKR — серая |
украинская, GUKR_ |
SYB — серая украинская (г. Новосибирск) |
||
GUKR_ALT — серая украинская (Алтай), REDSTEP — красная степная, KALM — калмыцкая, KRGZ — киргизский скот, MNGL — монгольский скот, HRFD — герефордская, HOL — голштинская, SIM — симментальская, Ho — наблюдаемая гетерозиготность, Ar — аллельное разнообразие, uHe — несмещенная ожидаемая гетерозиготность, uFis — коэффициент инбридинга (доверительный интервал 95 %).
При этом в других степных породах (KALM и REDSTEP) наблюдаемая гетерозиготность была несколько выше, чем в алтайской и сибирской популяциях серой украинской породы. В группах локального скота (KRGZ и MNGL), традиционно разводимого в условиях степи с минимальным селекционным вмешательством человека, значения наблюдаемой гетерозиготности и аллельного разнообразия превосходили аналогичные значения во всех остальных породах, кроме симментальской.
Чтобы уточнить изменения, затронувшие алеллофонд изучаемых групп скота, мы провели анализ аллельного разнообразия для каждого локуса (табл. 4).
Всего в исследуемой выборке идентифицировали 137 аллелей, наиболее полиморфными оказались локусы TGLA122, TGLA53 и TGLA227 (соответственно 22, 19 и 17 аллелей), наименьший полиморфизм был выявлен в локусах BM1824 и ETH10 (по 8 аллелей в каждом) (см. табл. 4).
В изучаемых популяциях степного скота наибольшее число аллелей и приватных аллелей обнаружили у калмыцкой породы (соответственно 89 и 4), у серой украинской породы наибольшее число аллелей найдено в монастырской популяции GUKR (78 и 2 аллеля), у красной степной породы всего выявили 86 аллелей, но приватных аллелей обнаружено не было.
С целью сравнения частот встречаемости аллелей мы составили аллельные профили для исследуемых степных пород скота (рис. 3). В каждом локусе были определены аллели с максимальной частотой встречаемости. Они были приняты как характерные (наиболее часто встречающиеся) для популяции.
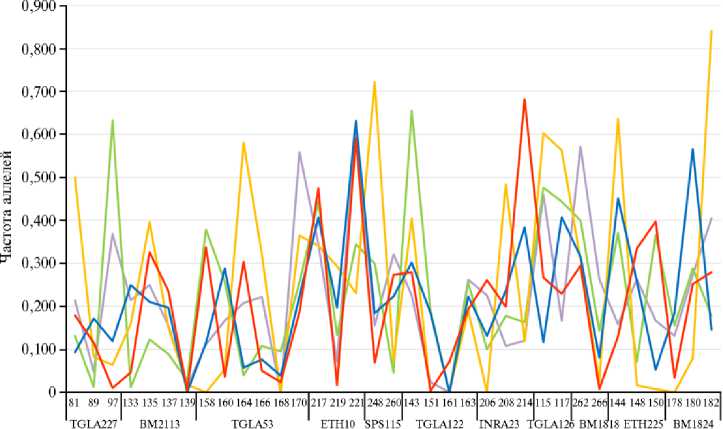
GUKR —GUKR_SYB —GUKR_ALT —KALM —REDSTEP
Рис. 3. Аллельный профиль степных пород скота разного происхождения: GUKR — серая украинская, GUKR_SYB — серая украинская (г. Новосибирск), GUKR_ALT — серая украинская (Алтай), KALM — калмыцкая, REDSTEP — красная степная (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы).
Наиболее сильно породы различались по локусу TGLA53, в котором было идентифицировано 5 аллелей (п.н.), различающихся в исследуемых популяциях скота. В локусе SPS115 выявили всего 2 аллеля, причем аллель 248 был общим для 9 популяций из 10, только в GUKR_ALT был обнаружен аллель 260 в этом локусе. В локусе ETH10 аллель 219 наиболее часто встречался у степных пород. Интересно отметить, что в локусе TGLA227 аллель 81 характеризовался наибольшими частотами встречаемости и был общим в породах степного скота. Также в локусе TGLA126 высокая частота встречаемости аллеля 115 была общей характеристикой для скота, традиционно разводимого в условиях степной зоны (красная степная и калмыцкая породы, киргизский и монгольский скот), кроме серой украинской породы. Для всех популяций серой украинской породы в этом локусе наиболее часто встречался аллель 117, общий также для европейских пород (HRFD, SIM, HOL).
Аллельные профили KALM и REDSTEP совпадали по 5 локусам из 11 (TGLA227, ETH10, SPS115, TGLA122, TGLA126). Профили KALM и серой украинской (учитывались аллели, выявленные в любой из исследуемых популяций) совпадали по 7 локусам из 11 (TGLA227, ETH10, SPS115, TGLA122, INRA23, BM1818, ETH225). Профили REDSTEP и GUKR совпадали по 9 из 11 локусов, причем 6 (TGLA227, TGLA53, ETH10, SPS115, TGLA122, BM1824) из них выявлены в сибирской популяции, 2 — в алтайской (BM2113, TGLA122) и 3 — в популяции из монастырского подсобного хозяйства (SPS115, BM1818, ETH225), что может быть связано с использованием общих улучшающих пород для этих групп.
Мы оценили генетическую структуру исследуемых популяций с использованием алгоритмов кластерного анализа (рис. 4).
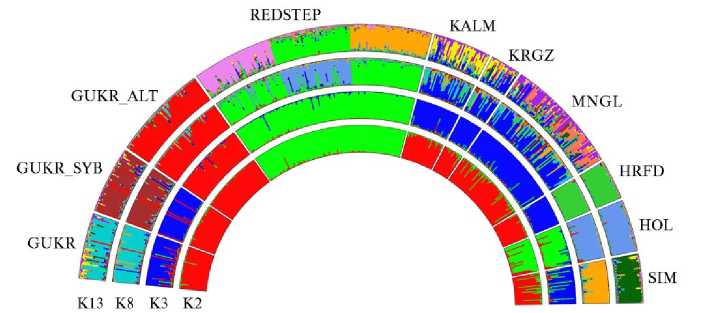
Рис. 4. Анализ генетической структуры популяций крупного рогатого скота степных пород: GUKR — серая украинская, GUKR_SYB — серая украинская (г. Новосибирск), GUKR_ALT — серая украинская (Алтай), REDSTEP — красная степная, KALM — калмыцкая, KRGZ — киргизский скот, MNGL — монгольский скот, HRFD — герефордская, HOL — голштинская, SIM — симментальская (ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста, 2024-2025 годы).
При числе кластеров К = 2 REDSTEP и HOL демонстрировали общие геномные компоненты, отличающие их от остальной выборки. При увеличении числа кластеров до К = 3 (оптимальное число К, согласно алгоритму G. Evanno с соавт.) (30), GUKR_ALT дифференцировалась в отдельный кластер. Голштинская и красная степная породы сохраняли общность компонентов, образуя единый кластер. В третий кластер вошли архивные популяции серой украинской породы, группы скота, традиционно разводимого в условиях степи (KALM, KRGZ, MNGL) и породы, используемые как улучшающие (HRFD, SIM).
При увеличении числа кластеров до К = 8, соответствующего числу пород, каждая из популяций серой украинской породы формировала отдельные группы, что могло быть следствием изменений аллелофонда современной и архивных популяций в результате селекционной работы и/или географической изоляции. Трансграничные породы также формировали породоспецифические кластеры, в REDSTEP выделилась группа особей, несущих компоненты, характерные для голштинского скота. Группа турано-монгольского скота демонстрировала преимущественно общие генетические компоненты.
Дальнейшее увеличение числа кластеров до К = 13 (оптимальное число К, согласно алгоритму J.K. Pritchard с соавт.) (26), показало, что в популяциях серой украинской породы, несмотря на значительную генетическую дифференциацию друг от друга, присутствовали животные, несущие генетические компоненты, общие для всех трех популяций этой породы. Архивные популяции (GUKR и GUKR_SYB) демонстрировали определенные генетические различия между собой. Однако в обеих этих группах были выявлены особи, которые несли генетические компоненты, доминирующие в современной алтайской популяции. Это позволяет сделать вывод о том, что популяция GUKR_ALT была сформирована на основе ограниченного генетического материала и прошла через «эффект бутылочного горлышка». Последующая изоляция, а также нетипичные для породы условия окружающей среды, вероятно, привели к распространению и закреплению в качестве основных генетических компонентов определенных производителей.
Следует отметить, что у некоторых особей в популяции GUKR_ALT сохранились генетические компоненты, свойственные архивной популяции из исконных мест разведения этой породы (GUKR).
В REDSTEP были выявлены три группы особей, отличающиеся друг от друга и несущих генетические компоненты, отделяющие их от других исследованных пород. Это может быть связано с формированием генетического пула этой породы на основе ограниченного числа производителей. В калмыцкой породе мы обнаружили как генетические компоненты, характерные для киргизского и монгольского скота, так и компоненты, отличающие ее от остальных пород турано-монгольского корня.
Результаты проведенных исследований согласуются с данными E. Solodneva с соавт. (31), которые показали наличие у калмыцкой породы скота общих предков с представителями как подольских пород, так и пород турано-монгольского происхождения. Генетическое разнообразие исследованных пород степного скота широко варьировало в зависимости от породы и было в среднем несколько выше (A r = 4,211-8,000, H o = 0,593-0,841) по сравнению с аналогичными показателями для трансграничных пород (A r = 4,990-5,711, H o = 0,668-0,744), что согласуется с данными других авторов (A r = 6,21-7,51, H o = 0,77-0,79 против A r = 4,90-5,32, H o = 0,64-0,73 соответственно для степных и трансграничных пород (12, 31). Интересно отметить, что в серой украинской породе аллельное разнообразие варьировало от 4,41 до 7,66 в популяциях соответственно Аскания-Нова и Алтая (32), в то время как, по нашим данными, алтайская популяция показала меньшее аллельное разнообразие по сравнению с образцами из подсобного хозяйства монастыря близ Киева (4,211 и 6,250 соответственно для алтайской и монастырской популяций). Значения показателей генетического разнообразия для красной степной породы также незначительно отличались от таковых в других работах: A r = 5,995 и H o = 0,718 против A r = 5,278, H o = 0,692 (33); H o = 0,746 (34).
Значительное генетическое разнообразие в изученных степных породах скота свидетельствует об их древнем происхождении и длительной адаптации к разнообразным экологическим условиям обширного ареала обитания. Преобладание естественного отбора при традиционном способе разведения локальных пород позволило сохранить богатый генофонд. Стоит отметить, что, несмотря на распространение исследуемых пород крупного рогатого скота в южных областях нашей страны, до настоящего времени значительная часть работ по их изучению посвящена анализу фенотипических показателей (35-38). Однако в последнее время в связи с возросшей доступностью коммерческих наборов для тестирования и оборудования стали появляться работы на основании анализа микросателлитных маркеров (34, 39). Тем не менее они часто носят характер локальных исследований поголовья в одном или нескольких хозяйствах и не предполагают рутинного мониторинга генофонда в течение нескольких лет или поколений животных.
Таким образом, мы выявили внутри серой украинской породы разнообразную генетическую структуру популяций, разводимых в разных географических регионах. Были идентифицированы изменения аллельного профиля в этих популяциях за несколько поколений. В обеих популяциях, разводимых в условиях Сибири, обнаружены особи, в разной степени сохранившие генетические компоненты, общие с предковой популяцией. Сравнительный анализ аллелофондов разных групп турано-монгольского скота показал, что, несмотря на проведение селекционной работы с калмыцкой породой, она сохранила значительную долю общих генетических компонентов с другими представителями турано-монгольско корня (киргизский и монгольский скот). Определены уникальные аллельные профили для каждой исследованной породы. Выявлены общие аллели для пород скота, традиционно разводимых в условиях степной зоны. Кластерный анализ позволил установить отсутствие значимой интрогрессии генетических компонентов трансграничных пород, которые наиболее часто используются как улучшающие, в аллелофонд трех локальных степных пород, разводимых в России. Изученные породы крупного рогатого скота показали уникальную генетическую основу, что делает их востребованными при разведении в условиях степной зоны, позволяя конкурировать со специализированными коммерческими породами и обеспечивая потребности местного населения в продуктах животноводства, в том числе при традиционном экстенсивном способе разведения. Стратегия дальнейшего сохранения генетических ресурсов этих пород может заключаться в предотвращении массового скрещивания с трансграничными породами, мониторинге генетического состояния популяций для своевременного выявления и предотвращения инбридинга, ведения качественного учета поголовья и обоснованного подбора родительских пар.
