Генофонд мтДНК представителей восточного варианта пахомовской культуры

Автор: Молодин В.И., Пилипенко А.С., Журавлев А.А., Трапезов Р.О., Ромащенко А.Г.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Эпоха палеометалла
Статья в выпуске: 4 (52), 2012 года.
Бесплатный доступ
В работе представлены результаты изучения серии образцов митохондриальной ДНК, полученных из костных материалов представителей восточного варианта пахомовской культуры из могильника Старый Сад в Барабинской лесостепи. Их сравнение с аналогичными данными по предшествующим популяциям региона позволил реконструировать процессы формирования генетического состава пахомовского населения с учетом археологического контекста. Выявлены компоненты генофонда, происхождение которых связано с автохтонным доандроновским населением Западно-Сибирской лесостепи и мигрантными андроновскими (и постандроновскими) группами. В статье приведена предварительная реконструкция этнокультурных взаимодействий в регионе в период миграции андроновского (федоровского) населения и формирования на его территории этнокультурных групп эпохи поздней бронзы, основанная на археологических и палеогенетических данных.
Археология, палеогенетика, древняя днк, митохондриальная днк, пахомовская культура, этнокультурные взаимодействия, андроновская (федоровская) культура, миграции
Короткий адрес: https://sciup.org/14522954
IDR: 14522954 | УДК: 572.1/4
Текст научной статьи Генофонд мтДНК представителей восточного варианта пахомовской культуры
Многолетние исследования археологических памятников эпохи бронзы в Обь-Иртышской лесостепи позволили разработать схему историко-культурного развития [Молодин, 1983, 1985], которая совершенствуется по мере появления новых источников [Молодин, 2010]. Сегодня картина существования и взаимодействия культур в эпоху поздней бронзы на означенной территории выглядит мозаичной, а в переходное от бронзы к железу время - еще более сложной.
Очевидно, что здесь доминирующее место занимали носители ирменской культуры, мощнейшего для своего времени образования. Они обитали на огромных пространствах Западно-Сибирской лесостепи практически от Ачинско-Мариинской котловины до левобережья Иртыша [Бобров, 1992; Молодин, 1985]. Носители ирменской культуры были автохтонным для региона населением, оказывавшим большое влияние на соседей и активно взаимодействовавшим с мигрантами. Северо-западную часть Обь-Иртышья занимали представители барабинского варианта суз-гунской культуры [Молодин, Чемякина, 1984] - пришлое с северо-запада население, испытывавшее влияние ирменцев [Молодин, Зах, 1985].
Еще в 1980-е гг один из авторов данной работы обратил внимание на отдельные погребальные комплексы на территории Барабы, выделяющиеся по инвентарю (прежде всего керамике) и погребальному обряду как от ирменских, так и от сузгунских [Мо-лодин, 1981]. Дальнейший анализ выявил антропологическую специфику людей, оставивших данные
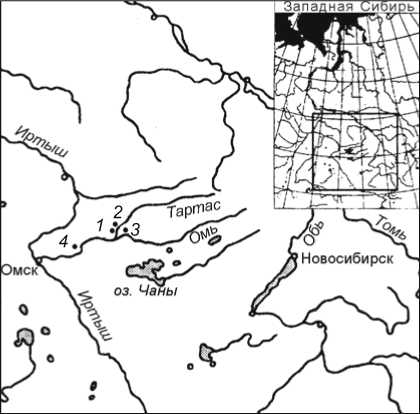
Рис. 1. Карта-схема расположения археологических памятников (некрополей) с проявлениями восточного варианта пахомовской культуры. 1 – Тартас-1; 2 – Старый Сад; 3 – Преображенка-3;
4 – Гришкина Заимка.
комплексы [Молодин, Чикишева, 1988]. Уже в момент открытия этих объектов была очевидна их связь с памятниками бегазы-дандыбаев-ской культуры в Центральном Казахстане, что подтверждало предположение М.Ф. Косарева и В.И. Матющенко о бегазы-дандыбаевском субстрате у носителей еловской культуры [Косарев, 1981, с. 28; Матющенко, 1974, с. 163]. Затем эти связи для более южных от Барабы районов Ку-лунды были прослежены в работе В.С. Удодо-ва [1994].
Особенно важные материалы были получены при раскопках могильника Старый Сад в Барабе* (рис. 1). Они позволили предположить существование в Западной Барабе в период поздней бронзы особого культурного образования [Молодин, Нескоров, 1992]. По обломку бронзовых удил, обнаруженному на могильнике Старый Сад (рис. 2, 5 ), исследованный памятник был отнесен к финальной стадии бронзовой эпохи – переходному от бронзы к железу времени. Следовательно, могильник Старый Сад представляет заключительную стадию па-хомовской культуры (но никак не начальную, как полагают некоторые исследователи [Костомаров, 2010, с. 22]).
Уже в 1980-х гг. было очевидно, что мы имеем дело с особым культурным образованием, хотя спе-
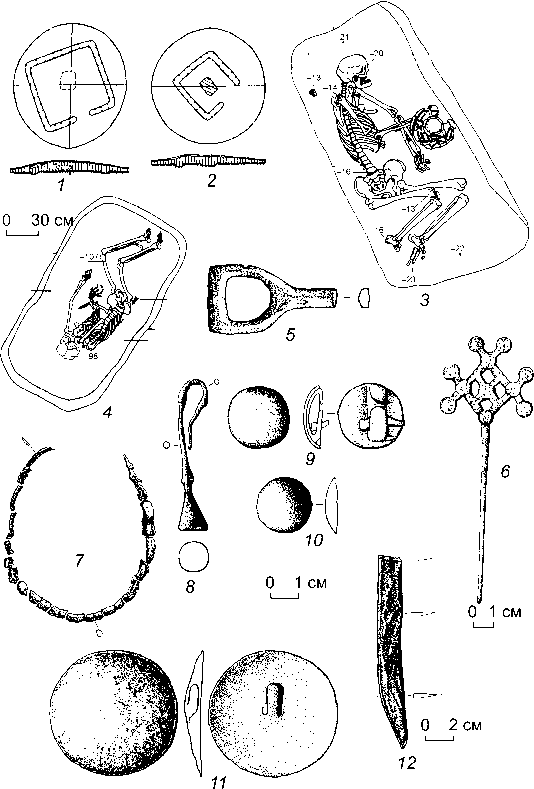
Рис. 2. Планы погребений ( 1–4 ) восточного варианта пахомовской культуры и найденный в них бронзовый инвентарь ( 5–12 ).
1, 2, 4–6 , 12 – Старый Сад; 3, 7–11 – Гришкина Заимка.
циальная работа по его выявлению не проводилась. В 1987 г. О.Н. Корочковой была выделена пахомов-ская культура [1987], ареал которой укладывался в пределы Тоболо-Иртышья [Корочкова, 2010]. На сегодняшний день ясно, что серия памятников в Бара-бинской лесостепи, о которых речь шла выше, представляет восточную периферию ареала пахомовской культуры (см. рис. 1). Со всей определенностью можно сказать, что эта культура многокомпонентна. По-видимому, она сформировалась на основе собственно пахомовцев (западных), у которых чрезвычайно ощутим южный бегазы-дандыбаевский, а в основе андроновский (федоровский) компонент (он ощутим и в материалах из барабинских памятников) (рис. 3, 2 – 4, 7, 10 ). Кроме того, налицо влияние автохтонной ирменской культуры (рис. 3, 5, 6, 8, 9, 11, 12 ), а также постоянное воздействие носителей сузгунской культуры с севера.

Рис. 3. Сосуды из могильников восточного варианта пахомовской культуры. 1-4, 6, 7, 10, 14 - Старый Сад; 5, 8, 11-13 - Гришкина Заимка; 9 - Преображенка-3.
На территории Барабы находится незначительная часть исследованных к настоящему моменту пахо-мовских памятников (см. рис. 1). Однако они (в первую очередь некрополь Старый Сад) содержат палеоантропологические материалы, которые в настоящее время являются, вероятно, наиболее представительными для пахомовского населения. В западной части ареала пахомовской культуры открыто и исследовано пока немного ее погребальных комплексов [Там же, с. 68-70].
Культурная и антропологическая специфика представителей восточного варианта пахомовской культуры, репрезентативность палеоантропологического материала делают в высшей степени актуальным изучение биологических характеристик носителей пахомовской культуры именно Барабинской лесостепи как традиционными методами физической антропологии, так и методами палеогенетики. В данной работе приводятся первые результаты исследования вариантов мтДНК представителей пахомовской культуры, погребенных в могильнике Старый Сад в Барабинской лесостепи, а также их анализа с точки зрения механизмов формирования генетического состава обитателей Западно-Cибирской лесостепи финала эпохи бронзы. Кроме того, излагаются представления о некоторых популяционно-генетических процессах, обусловивших формирование самобытных культур региона в означенный период.
Материалы и методы
Палеоантропологические материалы. Выборка образцов костных тканей носителей пахомовской культуры для проведения исследования получена из палеоантропологической коллекции ИАЭТ СО РАН, сформированной из материалов раскопок могильника Старый Сад. Для проведения палеогенетических исследований были отобраны длинные кости конечностей (бедренные, большие берцовые или плечевые кости) 11 индивидов, скелеты которых характеризуются наилучшей макроскопической сохранностью.
Предварительная обработка палеоантропологического материала и экстракция ДНК. Были использованы методы, описанные в работах [Pilipenko et al., 2010; Пилипенко, Молодин, Ромащенко, 2011]. Поверхность образцов обрабатывали 7 % раствором гипохлорита натрия для разрушения возможных загрязнений современной ДНК, затем механически удаляли слой толщиной ~1–3 мм и облучали образец ультрафиолетом не менее 1 ч с каждой стороны. Из компактного вещества длинных костей конечностей высверливали костный порошок. Для выделения ДНК костный порошок в течение 48 ч выдерживали в лизирующем 5М гуанидинизотиоционатном буфере [Pilipenko et al., 2010]. Выделение ДНК проводили методом фенол-хлороформной экстракции с последующим осаждением изопропанолом.
Анализ последовательности мтДНК. Амплификацию ГВС I мтДНК проводили двумя разными методами: амплификацию четырех коротких перекрывающихся фрагментов – с помощью однораундовой ПЦР [Haak et al., 2005], амплификацию одного длинного фрагмента – с помощью «вложенной» ПЦР (включала два раунда реакции) [Пилипенко и др., 2010]. Продукты амплификации фрагментов ГВС I клонировали в бактериальном векторе с помощью набора pGEM-T Easy Vector System Kit (Promega, США) и секвениро-вали 6–15 клонов для каждого фрагмента.
Определение последовательности нуклеотидов проводили с использованием набора реактивов ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, США) в соответствии с рекомендацими производителя. Продукты секвенирующей реакции анализировали на приборе
ABI Prism 3100 Genetic Analyzer (Applied Biosystems, США) в Центре коллективного пользования автоматического секвенирования ДНК СО РАН (Новосибирск). Полученные последовательности сравнивали c уточненной Кембриджской референсной последовательностью мтДНК [Andrews et al., 1999]. Филогенетическую интерпретацию последовательностей проводили в соответствии с существующей классификацией структурных вариантов мтДНК [van Oven, Kayser, 2009]. Для проведения филогеографического анализа использовали собственную базу данных по структуре ГВС I мтДНК, включающую более 20 тыс. образцов из современных популяций Евразии, которая была сформирована по литературным источникам.
Меры против контаминации . Все работы с древним материалом были выполнены в лабораторных помещениях, предназначенных для работы с древней ДНК: они оборудованы независимой системой приточной вентиляции с функцией фильтрации воздуха, создающей градиент его давления от более чистых к менее чистым помещениям, а также системой облучения ультрафиолетом, ламинарными шкафами второго класса защиты и др. Персоналом лаборатории использовались комплекты спецодежды для чистых помещений, проводилась частая смена стерильных перчаток. Все рабочие поверхности и приборы регулярно обрабатывались раствором гипохлорита натрия (5 %) и облучались ультрафиолетом. Полную процедуру экстракции и амплификации параллельно проходили древние образцы и контроли чистоты системы (без добавления палеоматериала) для выявления возможного загрязнения используемых реактивов и оборудования. Для всех сотрудников палеогенетической лаборатории, имеющих доступ в чистые помещения, а также для специалистов-антропологов, занимавшихся анализом палеоантропологических остатков и отбором материалов для генетического исследования, с целью выявления возможной контаминации материалов определена последовательность ГВС I мтДНК.
Результаты и обсуждение
До стоверные результаты по определению последовательности ГВС I мтДНК были получены для 8 из 11 образцов. Высокий показатель успешного получения образцов ДНК, пригодных для анализа, обусловлен как относительно благоприятными для сохранности ДНК в костных остатках климатическими условиями Центральной Барабы (и месторасположением могильника Старый Сад), так и строгим отбором включенных в выборку образцов по степени сохранности костного материала.
Все исследованные образцы мтДНК различаются по последовательности ГВС I (см. таблицу ). Структу-
Структура и филогенетическое положение образцов мтДНК представителей населения восточного варианта пахомовской культуры из могильника Старый Сад в Барабинской лесостепи
|
Образец |
Погребальный комплекс |
Гаплотип ГВС I мтДНК * |
Гаплогруппа мтДНК |
|
Sts1 |
Курган 65, погр. 2 |
223-298-325-327-362 |
C |
|
Sts2 |
Курган 67, погр. 2 |
051-183AC-189-256 |
U2e |
|
Sts4 |
Курган 83, погр. 1 |
126-294-296 |
T |
|
Sts6 |
Курган 6, погр. 1, скелет 1 |
093-223-227AC-290-311-319 |
A10 |
|
Sts7 |
Курган 27, погр. 1 |
223-298-327 |
C |
|
Sts9 |
Курган 59, погр. 1 |
256-270 |
U5a |
|
Sts10 |
Курган 62, погр. 1, скелет 1 |
126-163-186-189-294 |
T1 |
|
Sts11 |
Курган 49, погр. 1 |
148-223-227AC-290-311-319 |
A10 |
*Нумерация позиций в мтДНК соответствует Кембиджской референсной последовательности мтДНК [Andrews et al., 1999] без 16000 (т.е. позиция 223 в данной таблице соответствует позиции 16223 в референсной последовательности). Буквами после номера позиции обозначены замены оснований только в случае трансверсии.
ра гаплотипов позволяет однозначно определить принадлежность вариантов мтДНК к пяти гаплогруппам: западно-евразийским U2e, U5a, T и восточно-евразийским С и А10. Смешанная структура генофонда, сочетающая группы мтДНК, которые типичны для населения западной и восточной частей Евразии, характерна для всех исследованных нами к настоящему времени древних популяций западно-сибирского лесостепного пояса [Пилипенко, 2010].
Наличие в нашем распоряжении обширной выборки образцов мтДНК представителей населения Бара-бинской лесостепи и сопредельных территорий различных периодов эпохи бронзы позволяет проводить их сравнительный анализ. Поэтому полученные данные по генофонду мтДНК пахомовского населения с учетом археологического контекста можно использовать для выяснения механизмов формирования генетического состава этой группы.
По имеющимся сведениям в составе генофонда мтДНК населения восточной периферии ареала пахо-мовской культуры выделяются две группы вариантов. Первая группа, представленная линиями западно-евразийских гаплогрупп U2e, U5a и восточно-евразийских A10 и С, сближает рассматриваемую популяцию с автохтонными группами населения региона эпохи неолита и доандроновской бронзы – носителями усть-тартасской, одиновской и кротовской культур. Так, гаплогруппы U2e и U5a составляли основу западноевразийского компонента генофонда мтДНК перечисленных групп населения Барабы эпохи раннего металла, ранней бронзы и начала периода развитой бронзы, а гаплогруппа С в наибольшей степени представлена в восточно-евразийском компоненте их генофонда [Там же].
Следует подчеркнуть, что близо сть указанных компонентов генофонда пахомовского населения, представленного в антропологических материалах могильника Старый Сад, к ранним популяциям Бара-бы проявляется не только на уровне гаплогрупп или подгрупп мтДНК. Большинство конкретных структурных вариантов, выявленных в пахомовской выборке, присутствует и в генофонде ранних групп. Это вариант западно-евразийской гаплогруппы U2e с гаплотипом 051-183АС-189-256, который прослежен во всех исследованных нами группах населения Барабы от эпохи неолита до начала развитой бронзы, а также вариант гаплогруппы U5a с гаплотипом 256-270, обнаруженный в генофонде усть-тартасско-го населения Барабы эпохи раннего металла ([Там же], неопубликованные данные авторов). На уровне конкретных гаплотипов сложнее интерпретировать варианты гаплогруппы C в генофонде носителей па-хомовской культуры. Корневой гаплотип 223-298-327 является типичным компонентом генофонда ранних групп населения Барабы, особенно усть-тартасского ([Там же], неопубликованные данные авторов). Однако, по данным филогеографии, этот вариант широко представлен в Сибири в настоящее время, как, вероятно, и в эпоху бронзы. Следовательно, использовать его в качестве маркера генетической преемственности между разновременными группами на локальной территории затруднительно. Гаплотип 223-298-325327-362, по нашим данным, появляется в генофонде населения Барабы только в период миграции андро-новского населения на фоне общего увеличения разнообразия линий гаплогруппы С. Филогенетически предшествующий ему вариант гаплогруппы C с гаплотипом 223-298-325-327 зафиксирован в генофонде носителей кротовской культуры Барабы (могильник Сопка-2/4Б) [Там же]. Таким образом, этот вариант пока не может однозначно рассматриваться в качестве генетического маркера связи с доандроновским населением Барабы.
Особенно значимым признаком опосредованной генетической преемственности между представителями пахомовской культуры и доандроновским автохтонным населением региона является присутствие в выборке вариантов гаплогруппы А10, которая, по-видимому, сформировалась в генофондах населения лесостепной полосы Западной Сибири на местной основе и прошла длительную эволюцию на данной территории*. Таким образом, по большинству исследованных линий мтДНК пахомовское население, останки которого обнаружены в могильнике Старый Сад, близко к популяциям Центральной Барабы доан-дроновского периода эпохи бронзы, т.е. значительная его часть происходит от автохтонного доандроновско-го населения Западной Сибири.
Следует подчеркнуть, что на основании полученных нами данных преждевременно говорить об участии конкретных групп доандроновского населения (например, носителей кротовской культуры) в формировании генофонда барабинских пахомовцев. Нам пока не известен состав вариантов мтДНК в генофондах представителей других культур эпохи развитой и поздней бронзы лесостепной зоны Западной Сибири, кроме кротовской. Не исключено, что варианты мтДНК, сближающие пахомовскую и кротовскую (а также одиновскую) группы населения Барабы, были характерны и для других доандроновских популяций Западной Сибири. Чтобы прояснить этот вопрос, необходимо провести масштабное исследование генофонда ранних групп населения, а также носителей культур эпохи поздней бронзы (ирменской, карасук-ской и др.) различных территорий Западной Сибири.
Вторым компонентом в исследованной серии образцов являются два варианта гаплогруппы T. Линии гаплогруппы T, по нашему мнению, могут рассматриваться как генетические маркеры миграции андро-новского (федоровского) населения в Западную Сибирь. В выборке образцов мтДНК от доандроновской популяции варианты гаплогруппы T не представлены. Вместе с тем линии этой гаплогруппы характерны для западно-евразийского компонента генофонда мтДНК андроновского населения региона [Молодин и др., 2011].
Таким образом, серия образцов мтДНК восточных пахомовцев является, как минимум, двухкомпонентной и отражает участие двух генетических субстратов в формировании андроноидного населения Западной
Сибири - автохтонного доандроновского и пришлого андроновского. Возможно, андроновский (мигрант-ный) компонент в определенной степени является реликтовым, составлявшим доминант в популяции носителей бегазы-дандыбаевской культуры, и генетически и культурно продолжает линию развития ан-дроновского (федоровского) населения центральной части его ареала.
В целом наши данные соответствуют общепринятым представлениям о формировании андроноидных культур Западной Сибири [Косарев, 1981; Корочкова, 2010, 2011]. Очевидно, что на уровне материальной и духовной культуры этот процесс был многоступенчатым (многостадийным), а его характер и интенсивность в разных частях лесостепного пояса Западной Сибири имели особенности, что и обусловило появление разнообразных по своим чертам т.н. андроно-идных культурных групп.
Изменения в генетическом составе населения в некоторой степени коррелируют с трансформациями материальной культуры, возникавшими в ходе масштабного этнокультурного взаимодействия. В формировании структуры генофонда андроноидного населения Западной Сибири можно условно выделить как минимум два этапа. Первый этап связан с массовым проникновением в регион представителей андронов-ской (федоровской) этнокультурной общности и их вступлением во взаимодействие с автохтонными группами населения. Ему, по-видимому, соответствуют памятники собственно андроновской культуры, а также комплексы, оставленные автохтонным (позднекротов-ским для Барабы) населением, уже испытывавшим, возможно, опосредованное влияние со стороны мигрантов с запада, близких к андроновским (федоровским), и памятники, отражающие процессы взаимного культурного влияния мигрантов и аборигенов, т.е. содержащие комплексы с признаками синкретизма культур [Молодин, 2011]. Ярким примером является могильник Тартас-1, отражающий процессы взаимодействия пришлого андроновского (федоровского) и аборигенного позднекротовского населения в Центральной Барабе [Там же]. На популяционно-генетическом уровне в этот период происходил обмен генетическим материалом (на разных территориях степень его интенсивности была различной) между группами мигрантов и аборигенов, т.е. генофонд местного населения региона обогащался новыми генетическими компонентами. При этом генетическое влияние автохтонного населения на мигрантов, по крайней мере по женской линии, которую отражает генофонд мтДНК, возможно, было очень значительным, что прослежено по материалам могильника Тартас-1 [Пилипенко, 2010; Молодин и др., 2010]. В материальной культуре постепенно усиливалась роль андроновских традиций [Молодин и др., 2010, 2011]. Этот этап формиро- вания андроноидного населения, по-видимому, был насыщен несколькими миграционными событиями, обусловившими появление в регионе различных генетических элементов из андроновской среды, которая не была генетически однородной. Результаты этнокультурного взаимодействия фиксируются не только археологическими материалами, но и данными физической антропологии: в андроновских сериях Западной Сибири выделяются компоненты, происхождение которых связано с влиянием аборигенного населения [Чикишева, Поздняков, 2003].
Второй этап формирования поздних андроноид-ных групп, в частности пахомовского населения, характеризуется взаимодействием смешанных групп, образовавшихся в регионе на первом этапе генетических контактов мигрантов и аборигенов, с культурными образованиями бегазы-дандыбаевского типа (с уже видоизмененными, но отчетливо узнаваемыми андроновскими признаками) и с населением, генетически близким (или идентичным) к автохтонному, которое существовало в лесостепи до прихода андро-новских групп, испытавшим в какой-то степени анд-роновское влияние. С генетической точки зрения это означает сохранение «баланса» пришлого андронов-ского по происхождению и автохтонного компонентов в генофонде населения, что подтверждают первые результаты исследования серии образцов мтДНК носителей восточного варианта пахомовской культуры из могильника Старый Сад.
Разумеется, отсутствие палеогенетических данных по носителям бегазы-дандыбаевской, собственно пахомовской, ирменской и других культур эпохи поздней бронзы региона существенно затрудняет интерпретацию полученных нами результатов. Однако анализ археологических материалов памятников восточного ареала пахомовской культуры (и ряда памятников ее метрополии [Корочкова, 2010]) выявил в них позднебронзовый компонент культур «карасукского типа» (по определению Н.Л. Членовой [1981, с. 17]), включающий как собственные, так и реликтовые ан-дроноидные элементы.
Полученные данные позволяют предположить, что в генофонде носителей культур эпохи поздней бронзы усилилась роль элементов, связанных с автохтонными доандроновскими группами. В исследованной серии среди двух обозначенных выше компонентов – аборигенного и «андроновского» – несколько превалируют аборигенные для Западно-Сибирской лесостепи варианты мтДНК. Доминирование автохтонных компонентов в женской части генофонда популяций ранее было прослежено по материалам памятника Тартас-1, отражающим непосредственные контакты аборигенного и пришлого населения.
Формирование каждой культуры эпохи поздней бронзы в регионе в реальности, безусловно, было бо- лее сложным и многофакторным процессом, чем в предложенной нами модели. Каждая из вступавших во взаимодействие групп характеризовалась различными направлениями генетических контактов не только с северными популяциями, но и с другими соседними культурными группами, в т.ч. андроноидными, представлявшими южное направление (бегазы-дандыба-евцы), что фиксируется по археологическим данным [Молодин, Нескоров, 1992]. Работа по реконструкции этих генетических взаимодействий осложняется ввиду сходства структуры генофонда культурных групп, испытавших влияние андроновцев или их потомков.
Необходимо отметить, что полученные нами па-леогенетические результаты (как по носителям пахо-мовской культуры, так и по представителям предшествующих этнокультурных групп) включают только данные по составу линий мтДНК, которая наследуется по материнскому типу и, следовательно, в первую очередь отражает процессы формирования генетического состава женской части рассматриваемых популяций. Следует иметь в виду, что эти данные пока фрагментарны и характеризуют локальные группы населения. Для создания полной и объективной картины происходивших процессов и экспериментальной проверки высказанных в настоящей работе предположений необходимо проведение масштабных исследований с привлечением палеоантропологических серий всех основных групп – участников рассматриваемых процессов, а также изучение материалов по другим генетическим маркерам, прежде всего маркерам Y-хромосомы, отражающим генетическую историю мужской части популяций. Только с получением достаточного по объему и составу маркеров генетического материала возможно проведение более детального сопоставления накопленных обширных археологических данных по формированию культур эпохи поздней бронзы (в т.ч. пахомовской) с соответствующими популяционно-генетическими. Представленная статья является одним из первых примеров совместных поисков генетиков и археологов в этом направлении.
