Геномная оценка признаков воспроизводства свиней (Sus scrofa) крупной белой породы
 крупной белой породы")
Автор: Белоус А. А., Отраднов П. И., Ромазева М. Е.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетика, геномика, фенотип
Статья в выпуске: 6 т.60, 2025 года.
Бесплатный доступ
Размер помета - один из важнейших экономических признаков для свиноводства, поскольку он напрямую связан с эффективностью производства товарного и племенного поголовья. Важной характеристикой размера помета у свиней считается число отнятых поросят (ЧОП). Учет этого признака демонстрирует, в частности, сохранность молодняка от рождения до отъема и последующей постановки на выращивание. Существуют также признаки, характеризующие негативные аспекты воспроизводства: поросята, рожденные мертвыми (мертворожденные поросята, МР) и поросята, мумифицированные в период эмбрионального развития (мумифицированный плод, ММ). Впервые в России мы представили данные о генетической обусловленности количественных признаков воспроизводства свиней породы крупная белая. В условиях смоделированного отбора показано, что ранжирование согласно комплексной оценке племенной ценности особей дает иной результат, чем выявление животных, лучших по значениям признаков, с определением их вклада в потенциальный доход. Цель работы заключалась в селекционно-генетическом анализе характеристик воспроизводства у свиней крупной белой породы на основании геномных оценок племенной ценности и полногеномного анализа ассоциаций, а также прогноза потенциального дохода от использования оцениваемых особей в селекции. Объектом исследования были 709 свиней ( Sus scrofa ) крупной белой породы (126 свиноматок и 583 хряков) с записями о признаках ЧОП, МР и ММ. Источниками информации были записи первичного зоотехнического учета, включавшего наблюдаемые (фенотипические) значения признака и сведения о происхождении, а также результаты SNP-генотипирования образцов ткани животных. По каждому показателю были предложены модели для расчета геномных оценок племенной ценности (GEBV), которые показали, что наибольшую степень изменчивости демонстрировал признак ММ (R2 = 0,770), наименьшую - ЧОП (R2 = 0,479). Затем в соответствии с направлением селекции животных с контрастными консолидированными значениями показателей ранжировали от лучших к худшим. У отобранных лучших особей выполнили полногеномное исследование ассоциаций с SNP (однонуклеотидныt полиморфизмs) (GWAS). В общей сложности выявлено 702 SNP-маркера, взаимосвязанных с изучаемыми признаками. Интерес представляла аннотация общих для ряда признаков маркеров, по итогам которой идентифицировано 27 генов. Из них 10 ( PSMB1, CLDN3 , CLDN4 , FAM120B, ТCTE3 , WDR27 , MLXIPL , ELN , PARP14 , DTX3L ) рассматривались как перспективные (они имели взаимосвязь с изучаемыми признаками и ранее были обнаружены у свиней). Функциональная аннотация по биологическим путям (GO) была выполнена для 8 генов, взаимосвязанных с дифференциацией жировой ткани (GO:0045444), регуляцией кровяного давления (GO:0008217), развитием спинного мозга (GO:0021510), увеличением размера или массы скелетной мышечной ткани (GO:0048630), развитием скелетной мышечной ткани (GO:0007519), развитием нервной системы (GO:0007399), биосинтезом липидов (GO:0008610), развитием головного мозга (GO:0007420), формированием кожного барьера, ограничивающего проницаемость кожи (GO:0061436), формированием и структурой аортального клапана (GO:0003180) и развитием мужских половых органов (GO:0001673). Проведенное исследование углубляет представления о генетической структуре крупной белой породы и формировании геномной оценки свиней по показателям воспроизводства. С практической точки зрения комплексная индексная оценка животных может рассматриваться как метод повышения экономической эффективности производства свинины.
Геномная оценка, признаки воспроизводства, оценка племенной ценности, полногеномное ассоциативное исследование, селекционный индекс, свиньи, крупная белая порода
Короткий адрес: https://sciup.org/142247708
IDR: 142247708 | УДК: 636.4:577.2 | DOI: 10.15389/agrobiology.2025.6.1005rus
Genomic evaluation of reproductive traits in large white pigs (Sus scrofa)
Litter size is one of the most important economic traits for the pig industry because it is directly related to the efficiency of production of marketable and breeding stock. An important characteristic of litter size in pigs is the number of weaned piglets (NFP). This characteristic shows, in particular, the safety of young pigs from birth to weaning and subsequent rearing. There are also traits characterizing negative aspects of reproduction: piglets born dead (stillborn piglets, MP) and piglets “mummified” during embryonic development (“mummified” fetus, MM). The material of our study was a total of 709 Large White pigs with records of CPC, MR and MM traits. For each trait, genomic estimates of breeding value (GEBV) were calculated, the best individuals were selected and a genome-wide association study (GWAS) was performed. A total of 702 SNP markers (single nucleotide polymorphisms) interrelated with the traits under study were identified, but of interest was the annotation of markers “common” to a number of traits, which resulted in the identification of 27 genes, of which ten promising genes were identified, having relationships with the traits under study and previously found in pigs (Sus scrofa): PSMB1, CLDN3, CLDN4, FAM120B, TCTE3, WDR27, MLXIPL, ELN, PARP14, DTX3L. Functional annotation by biological pathways (GO) was generated for 8 genes related to adipose tissue differentiation (GO:0045444), blood pressure regulation (GO:0008217), spinal cord development (GO:0021510), skeletal muscle tissue size or mass increase (GO:0048630), skeletal muscle tissue development (GO: 0007519), nervous system development (GO:0007399), lipid biosynthesis process (GO:0008610), brain development (GO:0007420), formation of a functional barrier in the skin that limits its permeability (GO:0061436), aortic valve structure formation and organization process (GO:0003180), and male genital development (GO:0001673). This study contributes to the understanding of genetic structure and formation of genomic evaluation on reproductive performance in large white breed pigs, which can have a positive impact on the efficiency of pork production and show economic benefits, as reflected by the method of comprehensive index evaluation of animals.
Текст научной статьи Геномная оценка признаков воспроизводства свиней (Sus scrofa) крупной белой породы
Развитие геномных технологий с использованием биоинформатиче-ского анализа данных для практического применения — одна из основных задач стратегии научно-технологического развития Российской Федерации (1). Одной из интенсивных отраслей продуктивного животноводства считается свиноводство из-за быстрого роста животных и смены поколений. Фенотипическое проявление экономически значимых показателей поголовья связано с генетикой количественных признаков, в которой одним из ключевых методов анализа выступает полногеномное ассоциативное исследование (genome wide association study, GWAS). По сравнению с традиционным картированием QTL (quantitative trait loci) анализ GWAS позволяет идентифицировать SNPs (single-nucleotide polymorphisms), изменчивость которых значимо связана с изменчивостью анализируемых признаков (2).
В свиноводстве репродуктивные признаки относятся к наиболее экономически значимым при оценке эффективности селекции (3). При разведении и селекции чистопородных свиноматок расчет оценок племенной ценности (Estimated Breeding Value, EBV) животных по репродуктивным признакам помогает оптимизировать программу разведения и предсказать генетическую предрасположенность к улучшению или ухудшению показателей в следующих поколениях (4).
Оценка генетических параметров, связанных с характеристиками воспроизводства, — неотъемлемый компонент программ разведения и селекции свиней (5-7). На первоначальном этапе оценки приемлем подход, учитывающий массивы наблюдаемых значений признаков, а также родословные особей (8, 9), но для системы отбора необходима разработка и апробация геномной оценки, предназначенной для расчета несмещенных показателей, то есть не искаженных влиянием средовых факторов и взаимосвязанных характеристик (10).
В настоящей работе впервые в России показана генетическая обусловленность количественных признаков воспроизводства у свиней породы крупная белая. С помощью смоделированного отбора мы выявили, что результаты оценки особей по наблюдаемым значениям признаков с учетом вклада в потенциальный доход противоречат данным ранжирования животных на основании комплексной оценки племенной ценности.
Цель работы заключалась в селекционно-генетическом анализе характеристик воспроизводства у свиней крупной белой породы на основании геномных оценок племенной ценности, полногеномного анализа ассоциаций и прогноза потенциального дохода от использования оцениваемых особей в селекции.
Методика. Работу проводили на поголовье свиней (Sus scrofa) крупной белой породы (126 свиноматок и 583 хряка). Использовали два источника информации: массив записей первичного зоотехнического учета, включавший наблюдаемые (фенотипические) значения признака и сведения о происхождении животных, а также массив результатов SNP-генотипирования. Фенотипические данные генотипированных особей были взяты из базы данных программы Herdsman для ведения зоотехнического учета в хозяйстве (ООО Селекционно-гибридный центр «Топ Ген», п. Верхняя Хава, Воронежская обл.) . Данные датировались 2020-2024 годами.
Учитывали число мертворожденных поросят (МР) и мумифицированных плодов (ММ), а также признак, характеризующий сохранность потомства — число отнятых поросят (ЧОП). Подобное разделение было необходимо потому, что на популяционном уровне селекция ведется на снижение значений МР и ММ и на увеличение ЧОП.
Выделение ДНК из образцов ткани особей (ушной выщип) проводили с помощью наборов для выделения геномной ДНК серии ДНК-Экстран (ЗАО «Синтол», Россия) в соответствии с протоколом фирмы-производителя. Концентрацию двухцепочечной ДНК определяли с помощью флуориметра Qubit 2.0 («Invitrogen/Life Technologies», США). Для определения качества ДНК измеряли соотношение OD260/OD280 на cпектрофото-метре NanoDrop8000 («Thermo Fisher Scientific, Inc.», США). Для анализа использовали ДНК с OD260/OD280 = 1,6-1,8. Кроме того, качество ДНК оценивали методом гель-электрофореза в 1 % агарозном геле.
Полногеномное генотипирование проводили с использованием высокоплотных ДНК-чипов PorcineHD Genotyping BeadChip (платформа GeneSeek Genomic Profiler, «Neogen», США, , содержащих 66763 SNP.
После структурирования базы данных был проведен расчет GEBV (genomic estimates of breeding value) по методологии GBLUP (genomic BLUP) (11, 12). В общем виде модель BLUP имела вид:
у = Xb + Zu + e, [1] где y — вектор наблюдаемых значений признака, X — матрица распределения выборки по уровням фиксированных эффектов, Z — матрица распределения выборки по уровням рандомизированного эффекта, b — вектор оценок фиксированных эффектов, u — вектор оценок рандомизированных эффектов, e — вектор остатков.
В матричном виде нахождение решений моделей имело вид:
I UI = I
I X'X
X'Z
Z'X Z ' Z + G- 1 Z
I - 1*IX'yi ,
где G - 1 — обратная матрица геномного сходства исследуемых животных, ^e 1 ^
Л = -f = — --коэффициент, определяющий степень влияния на признак
^ а ^ 2
негенетических эффектов, h2 = ^^2 — коэффициент наследуемости.
Модель была ориентирована на оценку агрегатного генетического эффекта каждой особи. Это рандомизированный эффект, который рассчитывается с учетом поправки (нивелирования) наблюдаемых значений относительно статистически значимых эффектов среды и взаимосвязанных характеристики и матрицы геномного сходства особей по результатам SNP-генотипирования. Основными метриками качества модели BLUP-оценки в нашем исследовании выступали коэффициенты детерминации (R2), которые выражали долю изменчивости оцениваемого признака, обусловленную вариабельностью поддающихся учету эффектов среды и взаимосвязанных характеристик, а также статистическая достоверность влияния таких эффектов. Особенностью оценок племенной ценности хряков по исследованным признакам было то, что эти оценки основывались не на реальных данных о собственной продуктивности, а на матрице геномного сходства.
Оценку племенной ценности признаков проводили, основываясь на подходе, принятом в рамках методологии GBLUP, согласно следующим ма-
тематическим моделям:
МР = µ + Возраст0 + b1Д + a + e,[3]
ММ = µ + Возраст0 + b1КК + b2М + a + e,[4]
ЧОП = µ + ДОП + a + e,[5]
где в качестве фиксированных категориальных переменных учитывались: возраст 0 — возраст постановки на откорм, ДОП — дата отъема поросят; в качестве компонентов ковариации учитывались: КК — коэффициент конверсии корма (кг/кг), МР — число мертворожденных поросят (шт.), ММ — число мумифицированных зародышей (шт.), ЧОП — число отнятых поросят (гол.), Д — число поросят женского пола (гол.), М — число поросят мужского пола (гол.); μ — популяционная константа; a — рандомизированный категориальный эффект «животное» (представляющий собой оценку племенной ценности), e — остаток математической модели.
Модели разрабатывали с помощью библиотеки lme4 языка программирования R. Применение этой библиотеки обосновано возможностью
проверки влияния как категориальных, так и ковариационных (взаимосвязанных) эффектов на изменчивость признака в рамках одной процедуры. Стоит отметить, что проверка влияния эффекта «животное» с использованием такой библиотеки принципиально недостижимо, если по каждой особи имеется только одна запись. Поэтому при разработке уравнений учитывались только средовые факторы и компоненты ковариации по данным первичного зоотехнического учета.
Оценки племенной ценности рассчитывали согласно разработанным уравнениям в программах семейства BLUPF90, в частности применяли re-numf90 и blupf90+ (I. Misztal, D. Lourenco and collaborators, University of Georgia, США).
Для формирования уравнения селекционного индекса использовали методику, разработанную L.N. Hazel (1943) и адаптированную в ФГБНУ ФИЦ ВИЖ им. академика Л.К. Эрнста (13).
В общем виде уравнения могут быть представлены как li = q 1Xi + q2X2 + ... qmXm = X^EBVjQj = EBV'Q, [6] где q1-qm — весовые коэффициенты j-го признака, j ϵ (1, m), m — число признаков, включенных в уравнение индекса, X1-Xm — оценки племенной ценности i-го животного, i e (1, n), n — число животных в выборке, EBV' — вектор-строка оценок племенной ценности i-го животного, Q — вектор-столбец весовых коэффициентов уравнения селекционного индекса.
Весовые коэффициенты определяли как сумму членов вектора-столбца Q'p
Q' j = P - 1 *G j *W j , [7] где P - 1 — обратная матрица фенотипических коварианс, G j — вектор-столбец генетических (геномных) коварианс j- го признака ( вектор-столбец матрицы аддитивной генетической ковариации G , соответствующий j -му признаку), w j — экономический вес признака (стоимостной эквивалент единицы измерения признака).
Расчеты проводили с помощью разработанного в ФГБНУ ФИЦ ВИЖ им. академика Л.К. Эрнста специализированного программного обеспечения (14).
В качестве экономической поправки использовали модельную (полученную от хозяйства-оригинатора исследованной выборки) стоимость реализации (продажи) одного поросенка, равную 40000 руб. Выбор именно этого значения в качестве базового был обоснован в первую очередь тем, что рассматриваемые признаки так или иначе связаны с числом поросят, которые в дальнейшем могут быть использованы для реализации в товарные хозяйства.
Мертворожденные поросята и мумифицированные зародыши учитываются как недополученный доход (на основании данных о доходе, рассчитанном для числа отнятых поросят). Так, для показателей МР и ММ весовые коэффициенты индекса корректировались таким образом, чтобы итоговый вес был отрицательным, то есть формировал негативное давление отбора на оценочные значения племенной ценности этих показателей. Для показателя ЧОП, напротив, был сформирован положительный весовой коэффициент:
I = - 1114,68 ∗ MР - 824,40 ∗ ММ + 4,56 ∗ ЧОП. [8]
Для оценки вклада прогнозируемой племенной ценности особей в гипотетический доход, получаемый от продажи молодняка, рассчитывали гипотетический доход от продажи молодняка с учетом потерь, связанных с мертворождениями и мумификацией (на основе фенотипических значений)
(Д ф ); вклад племенной ценности особи в доход (Д EBV ); вклад племенной ценности относительно наблюдаемых значений (EBV) по следующим формулам:
Дф = 40000 ∗ ЧОП - 40000 ∗ (МР + ММ),[9]
ДEBV = 40000 ∗ EBVЧОП - 40000 ∗ (EBVМР +EBVММ),[10]
EBV% = 100 ∗ (ДEBV/Дф).[11]
Полногеномный ассоциативный анализ (GWAS) или GWAS для исследования множественных признаков в одной популяции (MS-GWAS) выполняли для 422 лучших животных, отобранных по геномным оценкам племенной ценности (GEBV) каждого из исследованных признаков. Лучшими особями по анализируемым признакам считались особи с самыми низкими (отрицательными) оценками геномной племенной ценности по МР и ММ, по признаку ЧОП, напротив, отбирали особей с самыми высокими (положительными) оценками. В результате сформировали три выборки, для каждой из которых проводили полногеномный поиск ассоциаций по признаку, используемому в качестве критерия отбора.
Чтобы минимизировать ошибки и повысить достоверность GWAS анализа, процедура включала несколько этапов контроля качества данных, результатов. Во-первых, при контроле качества образцов (особей) (call rate) исключались образцы с низким процентом успешного прочтения генотипов, обычно ниже 95-98 %, что указывало на плохое качество данных для конкретного индивидуума. Также проводили проверку на близкородственные связи (исключались образцы с высокой степенью родства, или коэффициентом инбридинга IBD > 0,185) и выполняли популяционную стратификацию (исключались выбросы, которые не соответствуют основной популяции, чтобы избежать ложных ассоциаций из-за различий в генетическом фоне). Во-вторых, осуществляли контроль качества SNP. Определяли частоту минорного аллеля (MAF) (исключались SNP с низкой частотой минорного аллеля, обычно MAF < 0,01 или < 0,05, поскольку их вариабельность может быть недостаточной для включения в статистическую модель); отклонение от равновесия Харди-Вайнберга (HWE) (исключались SNP, которые значительно отклонялись от равновесия Харди-Вайнберга, обычно p-value < 1½10 - 6 для контроля ложноположительных результатов). Также исключали SNP с пропусками, которые могли быть связаны с техническими ошибками генотипирования. В-третьих, после расчета статистик GWAS, чтобы минимизировать число ложноположительных результатов, проводили коррекцию с учетом множественного тестирования. Для этого использовали критерий Бонферрони, формирующий два порога значимости ассоциаций (15, 16). Первый — полногеномный уровень значимости (genomewide significance threshold, порог значимости, скорректированный на число тестируемых SNP). Второй — суггестивный порог (suggestive threshold), менее строгий порог для выявления потенциально интересных SNP. Порог рассчитывали по формуле:
-
т = »Ъ [12]
где p — базовый p-критерий, используемый для расчета поправки, N SNP — число SNP, прошедших фильтрацию.
Из-за половой дифференциации выборки, в исследование включили только SNP-маркеры, расположенные на аутосомах.
Для поиска генов, локализованных в области идентифицированных
SNP, использовали геномный ресурс Sscrofa11.1 базы данных NCBI . Сборку генома загружали в специально разработанный программный код для автоматизации процесса структурной аннотации.
Поиск генов-кандидатов осуществляли в диапазоне ±200 тыс. п.н. от непосредственной позиции ассоциированного маркера, что связано с размером окна неравновесного сцепления у европейских пород свиней (17).
Функциональную аннотацию и анализ обогащения генов выполняли с привлечением источников литературы, веб-базы данных DAVID и базы данных Pig QTL .
Результаты. Крупная белая порода свиней известна повсеместно благодаря многоплодию, высокой выживаемости поросят и неприхотливости при содержании. На 2023 год доля этой породы из всех, используемых на территории Российской Федерации, составила 18,5 % (18). Воспроизводительные качества основных свиноматок в Воронежской области по многоплодию составляют 14,5, по числу поросят при отъеме — 14,3 гол. При этом размер помета (общее число родившихся поросят) существенное влияет на экономический успех свиноводства. Показатель складывается из числа поросят, родившихся живыми и мертвыми, включая число мумифицированных.
Показатели воспроизводства в значительной мере зависят от технологических процессов внутри хозяйства (19, 20), что требует составления статистических моделей, учитывающих в генерализированном виде влияние условий среды и взаимосвязанных характеристик в качестве компонентов изменчивости изучаемого признака. Для создания таких уравнений, мы проанализировали изменчивость изученных признаков, обусловленную как факторами категориального характера, так и взаимосвязанными величинами (табл. 1).
1. Влияние анализируемых факторов на изменчивость признаков воспроизводства у свиней ( Sus scrofa ) крупной белой породы в разработанных моделях ( n = 709, ООО «СГЦ ТопГен», данные датируются 2020-2024 годами)
|
Фактор в модели |
Модель оценки признака |
||
|
МР, гол. |
ММ, шт. |
ЧОП, гол. |
|
|
Категориальные переменн |
ые (дисперсия признака, ед. изм.2) |
||
|
Возраст 0 |
0,07*** |
0,003* |
– |
|
ДОП |
– |
– |
18,551*** |
|
Коварианты (коэффициенты линейной регрессии) |
|||
|
КК, кг/кг |
– |
- 0,013* |
– |
|
М, гол. |
– |
0,106*** |
– |
|
Д, гол. |
0,409** |
– |
– |
|
Остаточная изменчивость, ед. изм.2 |
0,815 |
0,043 |
20,217 |
|
Коэффициент детерминации R2 |
0,590 |
0,770 |
0,479 |
Примечание. ДОП — дата отъема поросят, КК — коэффициент конверсии корма, Д — число поросят женского пола, М — число поросят мужского пола, МР — число мертворожденных поросят, ММ — число мумифицированных зародышей, ЧОП — число отнятых поросят.
*, **, *** Влияние фактора на изменчивость признака в модели достоверно соответственно при p ≤ 0,05, p ≤ 0,01, p ≤ 0,001. Прочерки означают, что достоверного влияния фактора не обнаружили.
В наибольшей степени предложенные модели описывали изменчивость признака ММ (R2 = 0,770), в наименьшей — ЧОП (R2 = 0,479). На среднем уровне также находилась обусловленность признака МР (R2 = 0,590).
На основании составленных статистических моделей была проведена оценка племенной ценности животных исследованной выборки. Оценка племенной ценности позволяет ранжировать особей от лучших к худшим в соответствии с поставленной селекционной целью. В результате формирования выборки из особей, считающихся лучшими для проведения полногеномного ассоциативного исследования, получили субвыборку особей с контрастными значениями признаков, консолидированными в рамках направления селекции (21). Выявляемые в подобных группах ассоциации с большей вероятностью будут иметь селекционное значение, чем при применении классического подхода, охватывающего весь массив данных (22, 23).
Популяционная характеристика оценок признаков в сформированных выборках представлена в таблице 2.
2. Оценки племенной ценности свиней ( Sus scrofa ) крупной белой породы в субвыборках, сформированных для MS-GWAS (ООО «СГЦ ТопГен», данные датируются 2020-2024 годами)
|
Признак-критерий отбора |
Описательная характеристика массивов оценок, M ± SEM, min-max, n = 422 ) |
||
|
МР, гол. |
ММ, шт. |
ЧОП, гол. |
|
|
МР |
-0,41±0,01* -1,77... -0,15* |
+0,01±0,00 -0,19...0,55 |
+0,01±0,01 -0,53...0,72 |
|
ММ |
+0,10±0,02 -1,27...1,57 |
-0,06±0,00* -0,20...-0,02* |
0,00±0,00 -0,53...0,72 |
|
ЧОП |
-0,02±0,02 -1,77...1,57 |
-0,01±0,00 -0,19...0,55 |
+0,10±0,00* 0,03...0,72* |
Примечание. МР — число мертворожденных поросят, ММ — число мумифицированных плодов, ЧОП — число отнятых поросят. Звездочками (*) отмечены описательные параметры целевого признака.
Для выявления генетической архитектуры репродуктивных характеристик в настоящем исследовании был проведен поиск генетических маркеров и локусов количественных признаков методом полногеномного ассоциативного исследования на свиньях, имеющих лучшие генетические оценки племенной ценности.
После фильтрации осталось 47798 SNP. Полногеномный порог значимости с использованием поправки на множественную проверку гипотез по Бонферрони составил 1,05½10 - 6 (0,05/47798), суггестивный порог — 1,05½10 - 5 (0,5/47798). Подобные уровни соответствовали стандартной практике в исследованиях на животных. Так, Q. Dong с соавт. (24) использовали эмпирическое пороговое значение 1½10 - 4.
Суггестивный порог представляет собой промежуточный уровень значимости, который используется в GWAS для выделения потенциальных ассоциаций между генетическими маркерами (например, SNP) и исследуемым признаком. Значимость, достигающая суггестивного порога, указывает на SNP, представляющие интерес для последующей проверки, но она недостаточна для утверждения достоверной ассоциации на уровне полногеномной значимости.
Конкретные значения суггестивного порога могут различаться в зависимости от размера выборки и числа проверенных гипотез. Например, в некоторых исследованиях используются пороги порядка 1½10 - 4, в других — вплоть до 1½10 - 3 (25). Такая вариация обусловлена необходимостью учета особенностей каждой отдельной выборки и различными методами коррекции при множественном тестировании.
По итогам MS-GWAS мы выявили 22 SNP-маркера, достоверно связанных с увеличением числа отнятых поросят, и 205 суггестивных. С прогнозируемым уменьшением частоты мертворождений связаны 7 (p < 0,0000001) и 186 (p < 0,000001) SNP, с учетом эффекта сокращения числа мумифицированных плодов — соответственно 14 и 275 SNP (рис.). Характерно, что обнаружены общие SNP для ЧОП и МР (4 полиморфизма), МР и ММ (2 SNP), ЧОП и ММ (1 SNP), что указывает на коррелятивные связи между признаками: с уменьшением числа мертворождений и мумификации увеличивался выход поросят при отъеме.
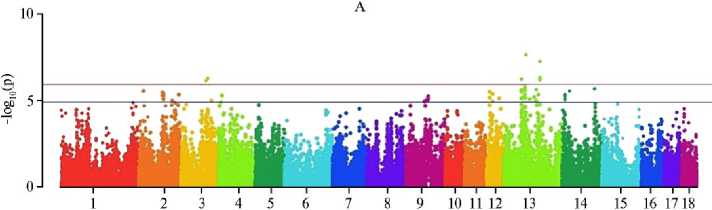
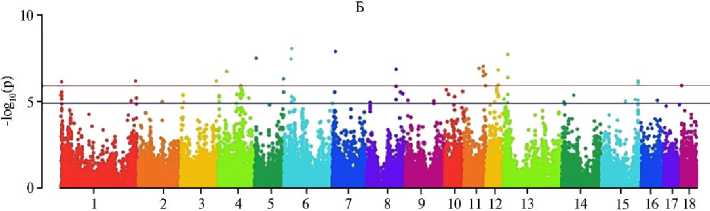
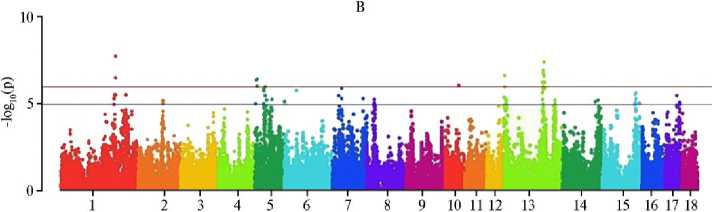
Chromosome
Графики распределения SNP, выявленных методом MS-GWAS, на хромосомах свиней ( Sus scrofa ) крупной белой породы по числу отнятых поросят (А) , числу мертворожденных поросят (Б) и «мумифицированных» плодов (В). Красная горизонтальная линия — полногеномный порог фильтрации по p-значению, синяя горизонтальная линия — суггестивный порог (ООО «СГЦ ТопГен», данные датируются 2020-2024 годами, n = 422),
Было выявлено 43 полногеномных и 666 суггестивных SNP по изучаемым признакам. Из них интерес представляет аннотация общих однонуклеотидных полиморфизмов, обнаруженных для двух или более признаков. Структурная аннотация общих полиморфизмов выявила наличие 27 протеин-кодирующих генов (табл. 3): TBP , PSMB1 , FAM120B* , DLL1 , ER-MARD , TCTE3 , PHF10 , C1H6orf120 , WDR27 , MLXIPL , VPS37D , DNAJC30 , BUD23 , STX1A , ABHD11 , CLDN3 , CLDN4 , METTL27 , TMEM270 , ELN , LIMK1 , KCNH8* , DIRC2 , HSPBAP1 , PARP14* , DTX3L , PARP9 (звездочкой помечены гены, в границах локализации которых расположены ассоциированные с изменчивостью SNP-маркеры).
-
3. Структурная аннотация SNP, демонстрирующих ассоциации с более чем одним из исследуемых признаков свиней ( Sus scrofa ) крупной белой породы (ООО «СГЦ ТопГен», данные датируются 2020-2024 годами, n = 422)
Номер хромосомы
Ген
SNP и достоверность (p)
Признак
1
TBP
PSMB1
1_242598 (4,176e - 06), 1_242598 (3,488e - 05)
ЧОП, МР
FAM120B DLL1
1_242598* (4,176e - 06), 1_242598* (3,488e - 05), ASGA0000014 (4,176e - 06), ASGA0000014 (3,488e - 05)
ЧОП, МР
ERMARD TCTE3 PHF10 C1H6orf120 WDR27
H3GA0000048 (2,719e - 06), H3GA0000048 (7,714e - 05)
ЧОП, МР
Ïðîäîëæåíèå òàáëèöû 3
|
3 |
MLXIPL |
H3GA0008665 (1,11e - 05), H3GA0008665 (2,839e - 05) |
ЧОП, МР |
||
|
13 |
VPS37D DNAJC30 BUD23 STX1A ABHD11 CLDN3 CLDN4 METTL27 TMEM270 ELN LIMK1 KCNH8 |
ALGA0123285* |
(2,046e - 05), ALGA0123285* |
(5,737e - 06) |
ЧОП, ММ |
|
DIRC2 HSPBAP1 PARP14 DTX3L PARP9 |
MARC0042648* |
(7,545e - 05), MARC0042648* |
(1,007e - 06) |
МР, ММ |
|
Примечание. МР — число мертворожденных поросят (шт.), ММ — число мумифицированных зародышей (шт.), ЧОП — число отнятых поросят (гол.). Звездочка (*) означает, что выявленный SNP расположен в нуклеотидной последовательности гена; остальные SNP не находятся в генах, но сцеплены с участками их локализации.
Анализ источников литературы показал, что ген PSMB1 взаимодействует с неструктурными белками и разрушает их через аутофагический путь, ингибируя репликацию вируса репродуктивно-респираторного синдрома свиней (РРСС) (26). Также блокаторами абсорбции на ранних стадиях инфекции РРСС выступают в комплексе гены CLDN3 и CLDN4 , которые относятся к семейству белков клаудинов (27). В исследованиях спермы хряков пород пьетрен и дюрок были идентифицированы гены FAM120B и TCTE3 , связанные со сперматогенезом и нарушением структуры белка PPARG (peroxisome proliferator-ativated receptor gamma), поддерживающего целостность гемато-тестикулярного барьера, нарушение функции которого ведет к уменьшению возможности успешного оплодотворения (28, 29).
Ген WDR27 ассоциирован с рН (кислотностью) свинины, и ранее была обнаружена его взаимосвязь с диабетом 1 типа (17). Ключевую роль в регуляции липогенеза играет ген MLXIPL , для которого установлена взаимосвязь с интенсивностью липидного метаболизма, а также низкая экспрессия в тканях свиней (30). Ген ELN ранее был идентифицирован как положительно экспрессируемый селективный ген, обнаруженный во внеклеточном матриксе. Для этого гена идентифицированы функции обеспечения целостности, а также выработки биохимических и биомеханических сигналов для клеточных и тканевых структурных элементов в организме свиней (31).
Ген PARP14 у помесных свиней второго поколения дюрок ½ (ланд-рас ½ йокшир) ассоциирован с толщиной шпика (32). Ген DTX3L выполняет важную роль в сигнальном пути Notch в процессах развития и контролирует миогенез — образование скелетной мышечной ткани во время эмбрионального развития (33).
Гены STX1A и ABHD11 недостаточно хорошо охарактеризованы, и их функция у свиней неизвестна, но эти же гены у человека (ортологи) связывают с нарушениями нейрогенеза и ожирением, что представляет значительный интерес с точки зрения их ассоциаций у свиней (34).
С помощью функциональной аннотации мы идентифицировали 8 генов, взаимосвязанных с метаболическими путями в библиотеках Gene On-thology (GO). Согласно информации, представленной в базе, ген FAM120B , расположенный на 1-й хромосоме, служит регулятором дифференциации жировой ткани (GO:0045444), ген DLL1 взаимосвязан с регуляцией кровяного давления у Sus scrofa (GO:0008217), с развитие спинного мозга
(GO:0021510), который обеспечивает передачу сенсорных и двигательных нервных импульсов между головным мозгом и периферическими нервными тканями, и с увеличением линейных размеров или массы скелетной мышечной ткани (GO:0048630). Ген PHF10 ассоциирован с развитием нервной системы (GO:0007399).
На 3-й хромосоме были идентифицированы 5 генов. Согласно Gene Ontology, MLXIPL , отвечает за биосинтез липидов (GO:0008610); DNAJC30 — за развитие мозга (GO:0007420); CLDN4 — за формирование в коже функционального барьера, который ограничивает ее проницаемость (GO:0061436) и, как отмечалось выше, служит одним из блокаторов адсорбции патогена на ранних стадиях инфекции РРСС. Взаимосвязь изменчивости числа мертворожденных поросят и мумифицированных плодов с частотой проявления репродуктивно-респираторного синдрома подтверждается клиническими исследованиями C. Unterweger с соавт. (35). Ген ELN влияет на формирование и организацию структуры аортального клапана (GO:0003180), а также на развитие скелетной мышечной ткани (GO:0007519). Ген LIMK1 связывают с развитием мужских половых органов (GO:0001673).
Отбор животных в племенное поголовье в значительной степени зависит от цели селекции. Для количественных признаков такая цель, как правило, заключается в увеличении или снижении их значений в популяции. Как показано в нашем исследовании, отбор в том или ином направлении определяет геномную структуру той доли популяции, которая в дальнейшем будет использована для получения потомства. Один из способов, позволяющих оптимизировать процедуру отбора по комплексу количественных признаков, — построение уравнения селекционного индекса. Мы определили наличие генетических маркеров, достоверно взаимосвязанных с изменчивостью всех трех изученных признаков воспроизводства. Для определения потенциального экономического эффекта от селекции были рассчитаны индексные оценки, выражающие монетарный эквивалент племенной ценности каждой особи с учетом взаимосвязей между признаками. Отбор 10 лучших по индексной оценке особей позволил сформировать модельное племенное ядро (табл. 4).
4. Модельное племенное ядро популяции свиней ( Sus scrofa ) крупной белой породы (10 лучших по индексной оценке особей; ООО «СГЦ ТопГен», данные датируются 2020-2024 годами)
|
Номер особи |
Оценка племенной ценности признака |
Индексная оценка, руб. |
Экономический показатель |
||||
|
МР, шт. |
MM, шт. |
ЧОП, гол. |
вклад ГП в доход, руб. |
потенциальный доход от реализации потомства, руб. |
ГП относительно фактического дохода, % |
||
|
1 |
- 1,77 |
0,05 |
0,24 |
1933,51 |
78551,52 |
520000,00 |
15,11 |
|
2 |
- 1,27 |
- 0,13 |
0,45 |
1527,99 |
74127,51 |
440000,00 |
16,85 |
|
3 |
- 1,50 |
0,21 |
0,08 |
1496,02 |
54814,59 |
440000,00 |
12,46 |
|
4 |
- 1,26 |
- 0,04 |
0,36 |
1437,61 |
66369,05 |
440000,00 |
15,08 |
|
5 |
- 1,24 |
- 0,06 |
0,03 |
1432,92 |
53423,83 |
520000,00 |
10,27 |
|
6 |
- 1,29 |
0,01 |
- 0,09 |
1427,65 |
47606,29 |
520000,00 |
9,16 |
|
7 |
- 1,12 |
- 0,11 |
- 0,23 |
1341,40 |
40225,15 |
520000,00 |
7,74 |
|
8 |
- 1,16 |
- 0,05 |
0,06 |
1335,46 |
50995,18 |
560000,00 |
9,11 |
|
9 |
- 1,11 |
- 0,09 |
0,72 |
1314,92 |
76804,38 |
480000,00 |
16,00 |
|
10 |
- 1,18 |
0,04 |
- 0,04 |
1283,54 |
44049,17 |
480000,00 |
9,18 |
|
Примечание. ГП |
— генетический прогресс; МР — число мертворожденных поросят (шт.), ММ — |
||||||
|
число мумифицированных зародышей (шт.) |
, ЧОП — число |
отнятых поросят (гол.). |
|||||
Так, отобранные животные, за исключением особей ¹¹ 6, 7, 10, характеризовались положительным вкладом агрегатного генотипа в изменчивость признака «число отнятых поросят». Также генетический вклад большинства особей, за исключением ¹¹ 1, 3, 6, 10, в изменчивость признака
«число мумифицированных зародышей» оценивался как отрицательный, иными словами, давал основания прогнозировать снижение этого показателя. По признаку «число мертворожденных поросят» все особи характеризовались отрицательным значением племенной ценности, что желательно в отношении этого признака.
Прогнозируемый (рассчитанный на основе наблюдаемых значений признаков воспроизводства) доход варьировал от 440 тыс. руб. до 560 тыс. руб. Этот показатель учитывал как число отнятых поросят, так и упущенную выгоду (мумифицированные зародыши и мертворожденное потомство).
Наряду с этим вклад генетического прогресса в доход, рассчитанный на основе оценочных значений племенной ценности (в отличие от индексной оценки, в этом показателе не учитывались взаимосвязи между признаками), демонстрировал вариабельность от 40225,15 до 78551,52 руб., что свидетельствует о существенном разрыве между лучшими и худшими результатами даже в топ-10 по выборке.
Вклад монетарного эквивалента генетического прогресса в потенциальный фактический доход от реализации потомства особи служит очередным подтверждением того, что BLUP-оценка, учитывающая факторы, влияющие на изменчивость признака, зачастую позволяет определить генетически лучших особей, которые не характеризуются наилучшими показателями при анализе только фенотипических значений. Так, наилучшая по потенциальному доходу особь ¹ 8 (560 тыс. руб.) демонстрировала только 9,11 % совокупного генетического прогресса ( ∼ 51 тыс. руб.), в то время как особи ¹¹ 2, 3, 4 с наименьшим значением фенотипического монетарного эквивалента (440 тыс. руб.) характеризовались одними из самых высоких показателей генетического вклада в изменчивость воспроизводительных качеств в выборке (соответственно 16,85 %, 12,46 %, 15,08 %, или ∼ 74 тыс. руб, ∼ 55 тыс. руб., ∼ 66 тыс. руб.).
Итак, мы провели геномную оценку свиней крупной белой породы по трем репродуктивным признакам — числу мертворожденных поросят, числу мумифицированных плодов и числу поросят при отъеме. Селекционно-генетический анализ включал связанные между собой процедуры: GBLUP-оценку, полногеномный поиск ассоциаций и моделирование экономической эффективности селекции животных на основании индексной оценки. Проведенное полногеномное ассоциативное исследование лучших особей по названным показателям изначально идентифицировало 702 гена, из которых по 8 генам получена функциональная аннотация и по 10 имелись подтверждающие их роль исследования. Наибольший интерес представляли гены PSMB1 , CLDN3 и CLDN4 , которые служат блокаторами адсорбции и репликации вируса репродуктивно-респираторного синдрома свиней — возбудителя инфекционного заболевания, распространенного в свиноводческой отрасли во всем мире. Применение методологии построения селекционного индекса позволило выявить наиболее сбалансированных по племенной ценности признаков воспроизводства особей при потенциальном вкладе в доход хозяйства. При этом выявлен значительный разрыв между фенотипическими и генетическими оценками: особи, для которых прогнозировался наивысший потенциальный доход, не всегда характеризовались высоким генетическим вкладом в ожидаемое улучшение признаков. Это подчеркивает важность использования GBLUP-оценки, которая позволяет выявить генетически перспективных особей, даже если их фенотипические показатели не являются наилучшими.
всероссийский НИИ интегрированного рыбоводства — филиал ФГБНУ Федеральный исследовательский центр животноводства — БИЖ им. академика Л.К. Эрнста, 142460 Россия, Московская обл., Ногинский р-н,
