Гистологические особенности дуги аорты Байкальской нерпы в возрастном аспекте

Автор: Аникиенко И.В., Рядинская Н.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Зоотехния и ветеринария
Статья в выпуске: 3, 2025 года.
Бесплатный доступ
Цель исследования - изучение гистологических особенностей стенки дуги аорты байкальской нерпы. Объект исследования - фрагменты дуги аорты в ее основании, в месте аортального расширения, перед отхождением левой сонной артерии, а также начальная часть грудной аорты сразу после отхождения левой подключичной артерии кумутканов (n = 10), неполовозрелых (n = 10) и половозрелых (n = 10) нерп. Исследование выполнено на кафедре морфологии животных и ветеринарной санитарии ФГБОУ ВО Иркутского ГАУ. Проводилось анатомическое препарирование и морфометрия различных участков дуги аорты. Гистологические срезы дуги аорты, изготовленные на микротоме, окрашивали гематоксилином и эозином, орсеином, по Маллори. Во всех возрастных группах максимальная толщина стенки отмечалась в луковице аорты, затем перед отхождением сонной артерии она достоверно снижалась, достигая минимального значения в грудной аорте. Кроме того, толщина стенки луковицы аорты у неполовозрелых нерп увеличивается в 1,46 раза, а у половозрелых - в 1,75 раза в сравнении с кумутканами. Наибольшее количество эластических мембран также обнаружено в «луковице аорты» в сравнении с остальными частями дуги аорты. Интима всех участков дуги аорты состоит из одного слоя эндотелиальных клеток с уплощенными и овальными ядрами и подэндотелиального слоя. Медиа основания аорты и луковицы аорты сформирована плотно расположенными пластинчатыми единицами, состоящими из эластических мембран, гладкомышечных клеток и расположенных между ними коллагеновых волокон. В аорте перед отхождением левой сонной артерии и грудной аорте эластические мембраны становятся более тонкими, прерывистыми, а пластинчатые более плотно расположены в участках, прилегающих к адвентиции сосуда. Наружная оболочка дуги аорты во всех возрастных группах хорошо васкуляризована, образована рыхлой соединительной тканью, состоящей главным образом из коллагеновых волокон с небольшим количеством тонких эластических волокон и фибробластов.
Байкальская нерпа, дуга аорты, интима, медиа, адвентиция, эластические мембраны
Короткий адрес: https://sciup.org/140309912
IDR: 140309912 | УДК: 611.132+619.48 | DOI: 10.36718/1819-4036-2025-3-118-130
Histological features of the aortic arch of the Baikal seal in the age aspect
The aim of the study is to investigate the histological features of the aortic arch wall in the Baikal seal. The object of the study was fragments of the aortic arch at its base, at the site of aortic dilation, before the origin of the left carotid artery, as well as the initial part of the thoracic aorta immediately after the origin of the left subclavian artery of kumutkans (n = 10), immature (n = 10) and mature (n = 10) seals. The study was carried out at the Department of Animal Morphology and Veterinary Sanitation of the Irkutsk State Agrarian University. Anatomical dissection and morphometry of various parts of the aortic arch were performed. Histological sections of the aortic arch, made on a microtome, were stained with hematoxylin and eosin, orcein, and Mallory staining. In all age groups, the maximum wall thickness was observed in the aortic bulb, then before the carotid artery originated, it significantly decreased, reaching the minimum value in the thoracic aorta. In addition, the wall thickness of the aortic bulb in immature seals increases by 1.46 times, and in mature seals - by 1.75 times compared to kumutkans. The largest number of elastic membranes was also found in the "aortic bulb" compared to other parts of the aortic arch. The intima of all areas of the aortic arch consists of a single layer of endothelial cells with flattened and oval nuclei and a subendothelial layer. The media of the aortic base and aortic bulb is formed by tightly arranged lamellar units consisting of elastic membranes, smooth muscle cells and collagen fibers located between them. In the aorta before the departure of the left carotid artery and the thoracic aorta, the elastic membranes become thinner, discontinuous, and the lamellar ones are more densely located in areas adjacent to the adventitia of the vessel. The outer shell of the aortic arch in all age groups is well vascularized, formed by loose connective tissue consisting mainly of collagen fibers with a small amount of thin elastic fibers and fibroblasts.
Текст научной статьи Гистологические особенности дуги аорты Байкальской нерпы в возрастном аспекте
Введение. Стратегия адаптации организма млекопитающих к водной среде обитания и жизни под землей (в норах) включает ряд структурных изменений. Адаптивная перестройка артериальных сосудов направлена на быструю доставку кислорода к тканям работающих органов.
У большого числа животных, способных к погружению (гренландского кита, кашалота, финвала, ларги, обыкновенного тюленя, калана и др.), описано ампулообразное расширение («луковица») восходящей части аорты [1–3]. Разница между основанием аорты и луковицей может достигать значительных величин, наиболее выражена она у настоящих тюленей. Так, у морского леопарда луковица аорты больше на 62,9 %, чем ее основание [4]. Расширение аорты, хотя и гораздо менее выраженное, чем у водных и полуводных видов, обнаружено у таких наземных животных, как бурый медведь и американский барсук. Вероятно, это связано с тем, что во время зимней спячки у данных животных, как и у млекопитающих, способных к погружению, отмечаются брадикардия, снижение температуры тела, задержка дыхания и др. [1].
Хотя микроструктура аорты, исследованная у трех видов дельфинов, ныряющих на мелково- дье или погружающихся на средней глубине, аналогична таковой у наземных млекопитающих, у дельфинов отмечается большая разница в пропорции пластинчатых единиц между восходящим и грудным сегментами аорты [5]. Аналогичные результаты были получены при исследовании луковицы аорты девяти видов глубоководных китообразных. Разница в морфометрических характеристиках различных сегментов стенки аорты, по мнению авторов, связана с адаптацией к нырянию [6]. Соотношение эластина и коллагена в восходящей части дуги аорты финвала (сельдяного кита) высокое (приблизительно 2 : 1) и значительно падает в грудной аорте (приблизительно 1 : 2). Кроме того, в восходящей части аорты волокна расположены таким образом, что она может деформироваться двухосно, выдерживая очень большие напряжения, и является основным емкостным элементом, гасящим пульсацию, в то время как нисходящая аорта, благодаря высокому содержанию коллагена, очень жесткая [7]. Исследования, проведенные позднее, показали, что жесткой является не только грудная аорта, но и ад-вентиция большинства артерий содержит особую жесткую форму коллагена, позволяющую выдерживать резкие изменения трансмурального давления, возникающие из-за ограниченной способности тканей реагировать на быстрые изменения окружающего давления океана во время погружения на глубину [8].
Гистологическое строение аорты описано у таких представителей настоящих тюленей, как хохлач, северный морской слон и обыкновенный тюлень. Луковица аорты у указанных видов содержала продольные и циркулярные эластические волокна, повторяющие их ход коллагеновые волокна, а также расположенные между ними гладкомышечные волокна. В нисходящей (грудной) аорте исследователи выявили только циркулярную ориентацию эластических волокон, как и в нисходящей аорте наземных млекопитающих. Кроме того, что восходящая аорта настоящих тюленей содержала большую концентрацию эластических волокон, чем нисходящая аорта [9, 10]. Сравнительно-гистологический анализ аорты видов семейства куньих показал, что самые мощные эластические волокна толщиной 10-12 мкм были в составе аорты калана, способного к погружению, хотя дуга аорты не имеет выраженного расширения [11].
Функциональное значение расширения аорты заключается в том, что она уменьшает нагрузку в левом желудочке сердца, возникающую вследствие повышенного периферического сопротивления и снижения упругости крупных артерий при длительном погружении. Луковица аорты является важным механизмом компенсации, обеспечивающим постоянное артериальное давление и артериальный кровоток во время удлиненного диастолического периода [12, 13]. Ультразвуковое исследование луковицы аорты у северных морских котиков и сивучей (представителей семейства Ушастые тюлени) подтвердило, что аорта значительно расширяется во время систолы и сокращается в течение всей диастолы. Таким образом, периферическая вазоконстрикция во время погружения приводит к изменению объема крови в аортальной луковице, что гарантирует непрерывный ток крови в течение всего диастолического периода [14]. Сравнение размеров аорты у семи представителей семейств Ушастые и Настоящие тюлени показало, что между диаметром луковицы аорты и массой тела существует прямая корреляция. Также была обнаружена линейная зависимость продолжительности обычного погружения от относительного диаметра луковицы
Вестник КрасГАУ. 2025. № 3 (216) аорты. В то же время максимальная продолжительность погружения не связана с диаметром аорты. Описанные связи подтверждают тот факт, что относительный диаметр луковицы аорты действительно определяет способность к погружению и частично влияет на длительность дайвинга [15].
Байкальская нерпа является глубоководным дайвером пресноводного озера Байкал. У байкальской нерпы изучены морфологические особенности артериальной системы сердца, селезенки, печени, органов желудочно-кишечного тракта и мочевыделительной системы. Исследователи отмечают высокую интенсивность кровоснабжения всех органов нерпы, но особенно селезенки и печени, участвующих в погружении и перераспределении крови [16-21]. В то же время дуга аорты описана очень обрывочно, а данные о ее топографии и микроструктуре в доступной нам литературе не были обнаружены. Морфофункциональные особенности дуги аорты байкальской нерпы необходимы не только для понимания анатомии водных млекопитающих, но и для ветеринарной практики, поскольку нерпа в условиях неволи содержится не только в нерпинариях и океанариумах России, но и в других странах.
Цель исследования - изучить гистологические особенности стенки дуги аорты байкальской нерпы в возрастном аспекте.
Объекты и методы. Исследование выполнялось на кафедре морфологии животных и ветеринарной санитарии ФГБОУ ВО Иркутского ГАУ. Объектом исследования являлась дуга аорты кумутканов (п = 10), неполовозрелых (п = 10) и половозрелых (п = 10) особей байкальской нерпы. Возраст кумутканов составлял от 1 месяца до 1 года, неполовозрелых нерп - от 1 года до 4 лет, половозрелых нерп от 4 лет и старше. Отлов нерп осуществлялся крупноячеистыми капроновыми сетями по разрешениям на добычу (вылов) в научно-исследовательских целях № 032015031172 (2015 г.), № 032021031373 (2021 г.) и № 0320220311974 (2022 г.) (оз. Байкал в административных границах Республики Бурятия). Возраст нерп определяли по спилу основания клыка [22].
Для изучения архитектоники дуги аорты применяли анатомическое препарирование. Материалом для гистологического исследования послужили фрагменты дуги аорты в ее основании, в месте максимального расширения (луковице), перед отхождением левой сонной артерии, а также начальная часть грудной аорты. Образцы фиксировали в 10 % нейтральном формалине, обезвоживали в изопропиловом спирте, пропитывали и заливали парафином LTB (ООО «Лаб-техбио», Россия). Срезы, изготовленные на микротоме «МЗП-01 ТЕХНОМ» (ООО «КБ Техном», Россия), окрашивали гематоксилином и эозином (обзорная окраска), орсеином (выявление эластических волокон), по Маллори (выявление мышечных и соединительных тканей). При окрашивании использовали как готовые наборы красителей (ООО «БиоВитрум», Россия), так и растворы, изготовленные самостоятельно (ООО «Био-Витрум», ООО «Лабтехбио», ООО «ЛабТехКом-плект»; ООО АО «РЕАХИМ»; «ИНТЕРХИМ» (Россия)). Морфометрию срезов проводили при помощи цифровой камеры Levenhuk M800 PLUS и программы «LevenhukLite» для анализа срезов. Все показатели, за исключением количества эластических мембран, приведены в мкм. Статистическую обработку проводили с помощью пакета программ Statistica 6.0. Данные в сравниваемых группах анализировали при помощи t-теста, различия считали статистически значимыми при р < 0,05.
Результаты и их обсуждение. У байкальской нерпы аорта, выходя из левого желудочка и направляясь краниально, на небольшом расстоянии (от 1,5 до 4,2 см) сохраняет свой диаметр неизменным, затем начинает постепенно увеличиваться и на уровне 4–6-го грудного позвонка аорта имеет максимальный диаметр и формирует луковицу. Между 3–5-ми грудными позвонками аорта наклоняется под углом около 90° и направляется влево. У некоторых живот- ных она может направляться краниолатераль-но. Аорта отдает у байкальской нерпы три магистральных сосуда, первой отходит плечеголовная артерия, затем левая сонная артерия и левая подключичная артерия. После отхождения последней, аорта у кумутканов на уровне 2–3-х грудных позвонков (у половозрелых – на уровне 4–5-го) резко делает поворот в каудальном направлении на 90° и идет вдоль позвоночного столба слева от него уже как грудная аорта.
Аорта (как восходящий, так и нисходящий ее участок) байкальской нерпы имеет типичные особенности сосуда эластического типа, в котором хорошо выражены три слоя: интима, медиа и адвентиция.
Во всех возрастных группах наибольшая толщина стенки отмечалась в луковице аорты, затем перед отхождением левой сонной артерии она достоверно снижалась, достигая минимального значения в грудной аорте (табл.). Тоньше всего стенка луковицы аорты была у кумутканов, у неполовозрелых нерп она увеличивалась в 1,46 раза, а у половозрелых – в 1,75 раза. При сравнении толщины стенки исследуемых участков аорты с ее основанием у кумутканов данное соотношение было представлено как 1,2 : 0,9 : 0,5; у неполовозрелых нерп – 1,2 : 1,0 : 0,4; а у половозрелых зверей – 1,2 : 0,8 : 0,5. Таким образом, стенка луковицы аорты во всех возрастных группах толще в 1,2 раза стенки основания аорты. Относительные размеры стенки аорты перед отхождением левой сонной артерии и грудной аорты больше всего отличаются у неполовозрелых нерп.
Морфометрические показатели дуги аорты байкальской нерпы в возрастном аспекте, мкм Morphometric parameters of the aortic arch of the Baikal seal in the age aspect, microns
|
Показатель |
Основание аорты |
Луковица аорты |
Дуга аорты перед отхождением левой сонной артерии |
Грудная аорта |
|
1 |
2 |
3 |
4 |
5 |
|
Кумутканы |
||||
|
Интима |
11,7±0,71 |
11,9±1,31 |
9,8±1,37 |
11,4±1,47 |
|
Медиа |
742,3±30,60 |
956,2±40,92* |
645,3±25,69 µ |
357,8±22,58*µ |
|
Адвентиция |
108,6±16,94 |
108,2±16,29 |
85,8±11,35 |
77,4±9,95 |
|
Толщина стенки |
863,1±36,35 |
1077,2±45,88* |
739,9±34,08*µ |
447,8±29,55*µ |
|
Количество эластических мембран |
76,6±3,22 |
94,7±3,71* |
61,4±3,80*µ |
43,9±1,33*µ |
Окончание табл.
|
1 |
2 |
3 |
4 |
5 |
|
Толщина мембран |
1,9±0,08 |
1,9±0,05 |
1,9±0,05 |
1,8±0,14 |
|
Неполовозрелые |
||||
|
Интима |
16,4±2,09к |
14,0±0,75 |
12,4±1,15 |
11,7±2,12 |
|
Медиа |
1078,0±67,50к |
1407,3±71,29*к |
1128,6±40,73µк |
442,5±12,71*µк |
|
Адвентиция |
172,3±17,30к |
148,2±16,93 |
111,8±5,66* |
81,0±5,71*µ |
|
Толщина стенки |
1261,3±72,91к |
1576,4±77,38*к |
1252,8±43,02µк |
531,3±16,92*µк |
|
Количество эластических мембран |
102,8±5,15к |
173,2±8,05*к |
101,0±5,83µк |
63,8±4,49*µк |
|
Толщина мембран |
2,5±0,10к |
2,2±0,06*к |
2,3±0,06к |
1,8±0,13*µ |
|
Половозрелые |
||||
|
Интима |
15,6±1,72 |
15,0±1,20 |
14,1±0,81к |
15,0±1,13 |
|
Медиа |
1318,3±70,32к |
1627,6±82,38*к |
1021,5±69,81*µк |
606,9±36,46*µкn |
|
Адвентиция |
196,1±28,31к |
246,4±28,81кn |
177,4±12,86µкn |
143,1±19,36кn |
|
Толщина стенки |
1536,2±88,05кn |
1893,2±94,06*кn |
1210,9±66,96*к |
760,4±39,19*кn |
|
Количество эластических мембран |
115,9±7,21к |
117,7±8,42кn |
69,1±4,20*µn |
60,6±1,77*к |
|
Толщина мембран |
2,1±0,05кn |
2,4±0,12к |
2,0±0,06кn |
2,0±0,11 |
Примечание :(*) – p < 0,05 – достоверность различий по сравнению c основанием аорты;(µ) – p < 0,05 – достоверность различий по сравнению c предшествующим участком дуги аорты; (к) – p < 0,05 – достоверность различий по сравнению c морфометрическими показателями дуги аорты у кумутканов; (n) – p < 0,05 – достоверность различий по сравнению c соответствующими показателями у неполовозрелых нерп.
Толщина интимы в исследуемых участках аорты достоверно не отличалась между собой. В то же время имелись отличия в ее размерах в возрастном аспекте: в основании аорты интима была больше у неполовозрелых байкальских нерп в сравнении с кумутканами, а в участке дуги перед отхождением левой сонной артерии у половозрелых нерп в сравнении с кумуткана-ми. Интима всех участков дуги аорты состоит из одного слоя эндотелиальных клеток с уплощенными и овальными ядрами и подэндотелиального слоя, состоящего из рыхлой соединительной ткани (рис. 1, 2).
В последней обнаружены очень тонкие эластические и коллагеновые волокна, формирующие паутинообразную структуру. Интима во всех отделах дуги аорты отделяется от медии эластичной мембраной.
Средняя оболочка имеет максимальный размер в луковице аорты и постепенно умень- шается после отхождения плечеголовной артерии, достигая минимальных значений в грудной аорте (табл., рис. 2). Толщина медии всех исследованных участков аорты у неполовозрелых и половозрелых нерп достоверно выше в сравнении с кумутканами. В то же время при сравнении толщины медии дуги аорты у неполовозрелых и половозрелых особей достоверные отличия обнаружены только в грудной аорте, которая была больше у половозрелых животных. Размер медии исследуемых участков аорты в сравнении с таковым в основании аорты у кумутканов был 1,3 : 0,9 : 0,5, у неполовозрелых нерп – 1,3 : 1,0 : 0,4, а у половозрелых – 1,2 : 0,8 : 0,5. Таким образом, у половозрелых нерп отмечено меньшее увеличение медии в расширении аорты, в то же время сужение медиальной оболочки перед отхождением левой сонной артерии, напротив, более выражено, чем у кумутканов и неполовозрелых нерп.
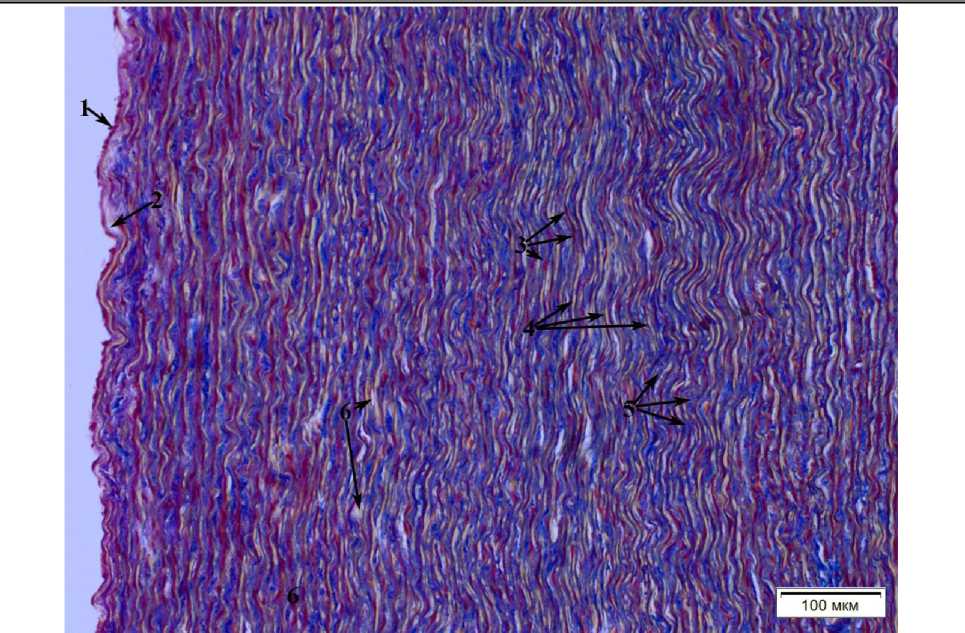
Рис. 1. Гистологическое строение основания аорты (самец, возраст 4+ года): 1 – эндотелий; 2 – подэндотелиальный слой; 3 – гладкие миоциты; 4 – эластические волокна (розовые);
5 – коллагеновые волокна (синие); 6 – капилляры (vasa vasorum)
Histological structure of the aortic base (male, age 4+ years): 1 – endothelium; 2 – subendothelial layer; 3 – smooth myocytes; 4 – elastic fibers (pink); 5 – collagen fibers (blue); 6 – capillaries (vasa vasorum)
Медиа основания аорты сформирована плотно расположенными, практически непрерывными, циркулярно расположенными эластическими волокнами, которые чередуются с гладкомышечными клетками и расположенными между ними коллагеновыми волокнами. Последние ориентированы аналогично эластическим волокнам. Такой комплекс эластических волокон, гладких мышц и коллагеновых волокон носит название пластинчатых единиц (рис. 2).
В аорте перед отхождением левой сонной артерии во внутренней части медии (примерно в одной третьей ее части) эластические мембраны становятся более тонкими, прерывистыми, пластинчатые единицы более плотно расположены в участках, прилегающих к адвентиции сосуда. Количество эластических мембран стенки аорты перед отхождением левой сонной артерии достоверно ниже в сравнении с луковицей аорты во всех возрастных группах, а у кумутка-нов и половозрелых животных также и в сравне- нии с ее основанием. Средняя толщина эластических волокон в описанном участке не отличалась от таковых в участках восходящей аорты.
Толщина медии грудной аорты значимо уменьшается в сравнении с участками восходящей части аорты, при этом наименьшую величину она имеет у кумутканов – (357,8 ± 22,58) мкм, наибольшую у половозрелых байкальских нерп – (606,9 ± 36,46) мкм. В средней оболочке грудной аорты эластические волокна ориентированы циркулярно, при этом в части, прилегающей к интиме, эластические волокна тонкие, в то время как слои гладкомышечных клеток, напротив, увеличиваются. Эластические волокна сильно извитые, формируют окончатые мембраны. Количество эластических волокон уменьшается в грудной аорте у кумутканов и неполовозрелых особей. Пластинчатые единицы, характерные для других участков дуги, располагаются только в наружной части медии.
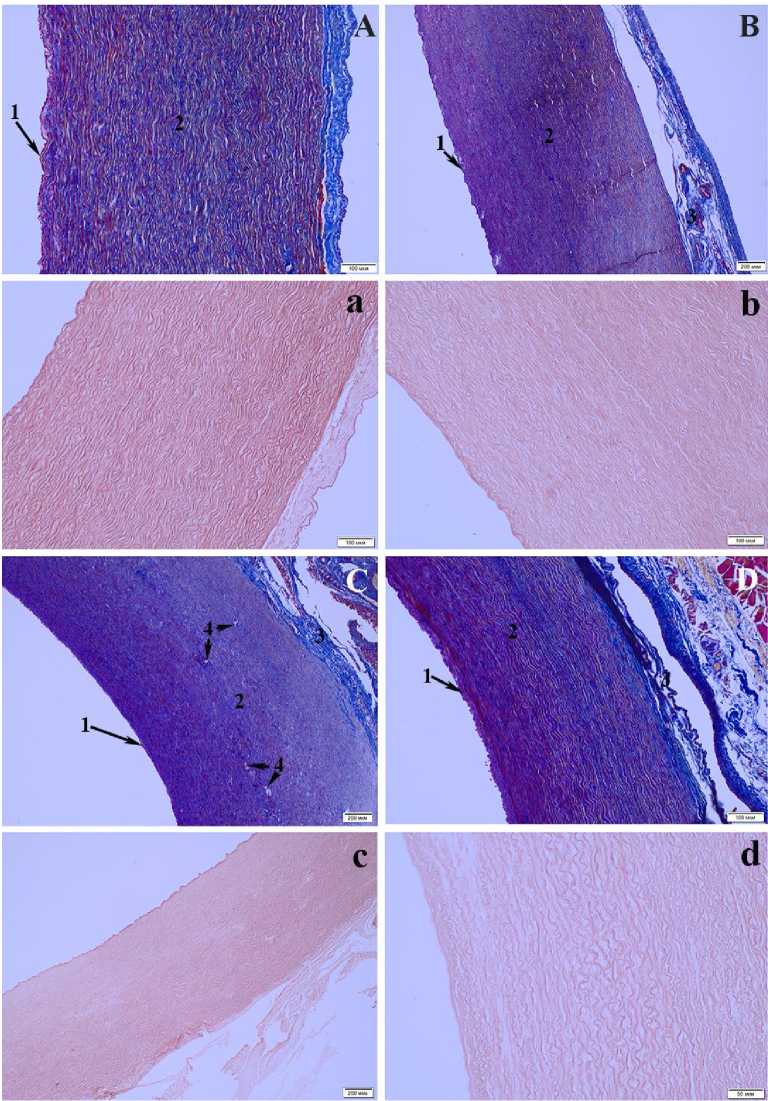
Рис. 2. Микроструктура дуги аорты байкальской нерпы (самец, возраст 4+ года):
А – основание аорты; B – расширение дуги аорты; С – дуга перед отхождением левой сонной артерии; D – грудная аорта после отхождения левой подключичной артерии, окраска по Маллори; a, b, c, d – перечисленные участки дуги аорты, окрашенные орсеином (1 – интима, 2 – медиа, 3 – адвентиция, 4 – vasa vasorum)
Microstructure of the aortic arch of the Baikal seal (male, age 4+ years): A – base of the aorta; B – expansion of the aortic arch; C – arch before the departure of the left carotid artery; D – thoracic aorta after the departure of the left subclavian artery, Mallory stain; a, b, c, d – listed areas aortic arches stained with orsein (1 – intima, 2 – media, 3 – adventitia, 4 – vasa vasorum)
Тонкостенные капилляры, доходящие практически до внутренней части медии, были обнаружены практически на всем протяжении дуги аорты, за исключением ее нисходящей части (грудной аорты).
Наружная оболочка дуги аорты во всех возрастных группах хорошо васкуляризована, образована рыхлой соединительной тканью, состоящей главным образом из коллагеновых волокон с небольшим количеством тонких эластических волокон и фибробластов. Начиная с ад-вентиции луковицы аорты и далее в нисходящей дуге отмечалось наличие жировых клеток, миелинизированных нервных волокон. У кумут-канов адвентиция аорты имеет меньший размер в сравнении с остальными возрастными группами, кроме того, в данной возрастной группе не выявлены значимые различия в исследуемых участках дуги. У неполовозрелых нерп наружная оболочка основания дуги аорты достоверно толще в сравнении с кумутканами. В данной возрастной группе адвентиция также имеет максимальную толщину в основании аорты, уменьшаясь перед отхождением левой сонной артерии и достигая минимального значения в нисходящей аорте. У половозрелых животных адвентиция имеет большую толщину по сравнению с кумутканами и неполовозрелыми особями. Максимальную толщину адвентиция половозрелых байкальских нерп имеет в луковице аорты – (246,4 ± 28,81) мкм, уменьшаясь в размере к грудной аорте.
У водных и полуводных животных исследователи отмечают три основных физиологических реакции на ныряние: апноэ (задержка дыхания), брадикардия, периферическая вазоконстрикция и гиперперфузия периферических тканей [23–25]. Снижение частоты сердечных сокращений при погружении необходимо компенсировать таким образом, чтобы жизненно важные органы не испытывали недостаток питательных веществ и кислорода, кроме того, после выныривания и восстановления дыхания следующей проблемой является реперфузия миокарда и других тканей. Важной анатомической особенностью, которая позволяет избежать последствий погружения, является луковица аорты. Во всех исследуемых возрастных группах байкальской нерпы толщина стенки луковицы аорты в сравнении с остальными участками дуги являлась максимальной. Мы связываем данную особенность со способностью нерпы к длительному, глубоководному погружению. Подтверждает данный факт и значительное увеличение толщины стенки с возрастом исследуемых животных. Луковица аорты эндемика Байкала имеет максимальный диаметр до отхождения плечеголовной артерии, в то время как у большинства настоящих тюленей луковица аорты формируется в точке после отхождения плечеголовной артерии [4, 9]. Возможно, такая особенность позволяет эндемику более равномерно распределять кровоток между магистральными сосудами правой и левой сторон.
У исследуемых животных более плотное расположение пластинчатых единиц отмечено в основании и луковице аорты – участках дуги, подвергающихся наибольшему давлению. Расположение эластических и коллагеновых волокон, а также гладких миоцитов в пластинчатых единицах обеспечивает высокую эластичность и прочность стенки аорты. Согласно A.S. Blix et al. (2016), луковица аорты обеспечивает «эффект резервуара» (нем. Windkessel effekt) и сохраняет ударный объем при каждом сокращении сердца, не позволяя чрезмерно увеличивать систолическое давление [26]. Данный факт объясняет, почему именно стенка корня и луковицы аорты у глубоководного ныряльщика – байкальской нерпы больше во всех возрастных группах, а также содержит плотно расположенные пластинчатые комплексы на протяжении всей медии в отличие от других участков дуги.
Интересно, что количество эластических волокон в луковице аорты байкальской нерпы было выше в сравнении с основанием только у неполовозрелых животных и кумутканов. Также мы обнаружили, что толщина эластических волокон в корне и перед отхождением левой сонной артерии от аорты была выше у неполовозрелых животных как в сравнении с кумутканами, так и с половозрелыми животными. Исследования физиологии дайвинга байкальской нерпы свидетельствуют о том, что длительное погружение является очень энергозатратным, что предполагает возможность существования у нерп метаболической депрессии в условиях интенсификации гликолиза. Нерпы чаще совершают короткие, не длительные погружения, чем продолжительные и глубоководные [27, 28].
Кроме того, исследователи отмечают наличие онтогенеза дайвинга, более глубокие и длительные погружения чаще осуществляют половозрелые животные [29]. Мы предполагаем, что хотя неполовозрелые нерпы и приобрели способность к погружению на большие глубины и являются более опытными дайверами, чем ку-мутканы, гормональная регуляция сосудистого тонуса при погружении у них еще не совершенна. И такая адаптивная особенность, как более толстые и многочисленные эластические волокна в восходящей аорте, позволяет ей эффективнее реализовывать эффект Виндкесселя при погружении.
Важно отметить, что хотя нервная регуляция дайвинга изучена подробно, влияние вазоактивных гормонов на тонус и расширение сосудов до сих пор практически не исследовано. В доступной нам литературе была обнаружена только одна работа, посвященная исследованию уровней вазоактивных веществ (ангиотензина II, вазопрессина и предсердного натрийуретическиго пептида) у спящих детенышей северного морского слона и тюленя Уэдделла как во время обычного дыхания (эупноэ), так и во время апноэ. Апноэ у детенышей во время сна сопровождалось брадикардией, повышением уровня вазодилататора – натрийуретического пептида и снижением уровней вазоконстрикторов – вазопрессина и ангиотензина II. Авторы также отмечают, что факторы, оказывающие влияние на кардиореспираторную функцию, могут изменяться в процессе онтогенеза [30].
С другой стороны, более высокое содержание эластических волокон в луковице аорты ку-мутканов и неполовозрелых животных может быть связано с меньшей толщиной стенки у данных возрастных групп и является компенсаторным механизмом, позволяющим аорте выдерживать давление.
Нами также были обнаружены капилляры, которые у байкальской нерпы немного не доходили до интимы во всех участках аорты, кроме грудной. Аналогичные сосуды в медии были описаны у обыкновенного тюленя и тюленя Уэдделла [10, 13]. При исследовании дуги аорты хохлача для лучшей визуализации микрососудов (vasa vasorum interna) стенка восходящей дуги аорты промывалась сульфатом бария под обычным для нее артериальным давлением.
Оказалось, что данные сосуды начинаются в просвете аорты, а заканчиваются в адвентициальной оболочке аорты, присоединяясь к сосудам обычного артериального дерева. Сосуды имеют разный диаметр, и их стенка в основном представлена только эндотелием. По мнению авторов, несмотря на то, что поток крови через сосудистую сеть vasa vasorum interna может быть довольно небольшим, он может иметь решающее значение, обеспечивая необходимое питание и участвуя в поддержании целостности стенки аорты [26]. Мы полагаем, что при расширении аорты данные сосуды увеличиваются в диаметре, способствуя таким образом снижению давления на стенку аорты, а также обеспечивая более эффективное кровоснабжение в стенке сосуда.
Резкое уменьшение толщины грудной аорты (практически в два раза в сравнении с основанием аорты) байкальской нерпы и увеличение в ее стенке количества гладких миоцитов также играют важную роль в адаптации к нырянию. Мы полагаем, что благодаря сужению и увеличению количества гладкомышечных клеток во внутренней части медии нисходящей аорты кровенаполнение магистральных сосудов, питающих голову, шею, а также передние конечности (ласты) нерпы, будет сохранено в любых условиях.
Толщина адвентиции была максимальной у половозрелых животных как в сравнении с ку-мутканами, так и с неполовозрелыми животными. Мы связываем данный факт с увеличением размера тела и сосудов животных.
Заключение. Строение дуги аорты байкальской нерпы представляет собой морфологическую адаптацию к глубоководному погружению и имеет следующие особенности:
-
1. Луковица аорты располагается в месте поворота дуги аорты влево до отхождения плечеголовной артерии. Стенка луковицы аорты имеет наибольшую толщину в сравнении с остальными участками дуги аорты у животных всех исследуемых возрастных групп. Кроме того, при увеличении возраста увеличивается и толщина стенки луковицы аорты: у неполовозрелых нерп она увеличивалась в 1,46 раза, а у половозрелых – в 1,75 раза в сравнении с ку-мутканами.
-
2. В основании и луковице аорты эластические, коллагеновые волокна и гладкие миоциты располагаются в плотно расположенных пластинчатых единицах, в участке аорты перед отхождением левой сонной артерии и грудной аорте пластинчатые единицы располагаются только в наружной части медии.
-
3. Среднее количество эластических волокон корня и луковицы аорты выше, чем в остальных исследуемых участках аорты.
-
4. Микрососуды (vasa vasorum interna) средней оболочки восходящей части аорты практически доходят до интимы.
-
5. Стенка грудной аорты значительно уменьшается (практически в два раза) в отличие от участков восходящей части аорты, а также содержит во внутренней части медии слои гладкомышечных клеток.
