Характеристика ценофлоры Ulmus pumila L. в Западном Забайкалье

Автор: Холбоева Светлана Александровна
Журнал: Вестник Бурятского государственного университета. Биология, география @vestnik-bsu-biology-geography
Рубрика: Биология
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
В статье обсуждаются особенности ценофлоры естественных сообществ Ulmus pumila L. в Западном Забайкалье. Вид на территории региона находится на северо-западной границе ареала. Рассмотрен систематический, экологический, биоморфологический состав флоры ильмовников в сравнении с формациями ильмовников с U. pumila Восточного Забайкалья и близкой по генезису формацией Armeniaca sibirica Селенгин-ского среднегорья. Спектр преобладающих семейств и родов в формациях показывает их высокое сходство. Анализ экологического состава ценофлоры ильмовников Западного Забайкалья подтверждает ее степной характер. В географической структуре преобладают южносибирские и евразиатские элементы флоры. Влияние восточноазиатского и дауро-маньчжурского центров формирования флор значительно ослабляется, тогда как южно-сибирский центр становится определяющим. В работе использован метод расчета активности видов, который показал важнейшую фитоцено-тическую роль кустарника Spiraea aquilegifolia и травянистых видов - эдификаторов и диагностических видов центральноазиатских и южносибирских настоящих и луговых степей.
Ценофлора, западное забайкалье, активность видов
Короткий адрес: https://sciup.org/148317865
IDR: 148317865 | УДК: 581.9(571.54) | DOI: 10.18101/2587-7143-2018-2-10-23
Characteristics of Ulmus pumila L. coenoflora in Western Transbaikalia
The peculiarities of the Ulmus pumila L. coenoflora of the natural communities in the Western Transbaikalia are discussed in the article. This species is located at the northwestern border of the area. The systematic, ecological, biomorphological composition of the U. pumila community coenoflora is considered in comparison with the U. pumila formations from Eastern Transbaikalia and the similar in genesis formation of the Armeniaca sibirica of the Selenga middle mountains. The spectrums of the predominant families and genus in the formations shows their high similarity. Analysis of the ecological composition of U. pumila community coenoflora in Western Transbaikalia confirms its steppe character. In the geographical structure, the Southern Siberian and Eurasian elements of the flora are prevailed. The influence of the East Asian and Dauro-Manchurian centers of flora formation is considerably weakened, whereas the South Siberian center becomes decisive. The method of calculation of species activity was used, which showed the most important phytocoenotic role of Spiraea aquilegifolia and herbaceous species - edificators and diagnostic species of Central Asian and South Siberian true and meadow steppes.
Текст научной статьи Характеристика ценофлоры Ulmus pumila L. в Западном Забайкалье
В пределах Республики Бурятия ильм низкий произрастает в южных и центральных районах в Селенгинском среднегорье, в долинах рек, на склонах низких гор и террасах в пределах степного и лесостепного поясов. В южных районах (долина р. Джида) ильмовники приурочены к ложбинам стока в предгорьях, на южных шлейфах невысоких отрогов. В среднем сомкнутость крон деревьев, как правило, очень низка (до 0,3), местами деревья встречаются единично.
Камелин Р. В. (1987) в своей известной работе «Флороценотипы растительности Монгольской Народной Республики» обосновал выделение флороценотипа гемиксерофильных азиатских листопадных редколесий и кустарников, возникшего в плейстоцене-постплейстоцене. Среди прочих к данному флороценотипу он относил формации Armeniaca sibirica и коренные группировки Ulmus pumila, не включая его пойменные комплексы. Эти формации на территории Западного Забайкалья находятся на северо-западной границе ареала. Реликтовый характер 10
этих формаций отмечали многие исследователи растительности (Камелин, 1987, Беликович, Галанин, 2006; Намзалов, Сахьяева, 2016).
Цель работы — дать характеристику ценофлоры естественных сообществ U. pumila в Западном Забайкалье.
Материалы и методы. Нами сделаны полные геоботанические описания по стандартным методикам в районах произрастании U. pumila в пределах Республики Бурятия.
Всего учтено 153 описания с участием U. pumila , хранящихся в базе данных кафедры ботаники Бурятского государственного университета. Из них отобраны естественные сообщества, произрастающие на ненарушенных распашкой почвах, таким образом, из анализа исключены вторичные ильмовники, активно формирующиеся на залежах. Вторым критерием отбора послужило обилие U. pumila в фитоценозах от 1 и более баллов по шкале Браун-Бланке, и формирование ильмом заметного древесного яруса. Всего в анализ включено 23 описания.
Анализ ценофлоры производился с помощью системы IBIS 7.1 (Зверев, 2007). Расчет активности видов выполнен по формуле (Зверев, Королюк, 2016):
Act =
N
C
F x 100% £ '
NN
= 10 x
л / F x C^
N
%
где Act — расчетная активность таксона для ценофлоры в процентах;
N — число единичных описаний, объединенных в ценолфору;
F — встречаемость — абсолютное число описаний, в которых зарегистрирован таксон;
С i — проективное покрытие таксона в i-ом описании;
С Σ — сумма проективных покрытий таксона во всех описаниях.
Выделение элементов флоры (географических, поясно-зональных, биоморфо-логических) проводилось с учетом работ Пешковой Г. А. (2001); Малышева Л. И., Пешковой Г. А. (1984).
Обсуждение результатов. Систематический анализ ценофлоры. Всего в описаниях зарегистрировано 165 видов высших сосудистых растений, относящихся к 40 семействам. Из них одновидовых семейств 16.
Спектр ведущих семейств — Asteraceae, Poaceae, Fabaceae, Rosaceae, Lamiaceae — характерен для степной флоры Сибири (Рещиков, 1961, Пешкова, 2001, Намзалов, 2015) и указывает на степной генезис флоры ильмовников (табл. 1).
Таблица 1
Ведущие семейства в ценофлоре ильмовников
|
№ |
Семейство |
Число видов |
% |
|
1. |
Asteraceae |
27 |
16,4 |
|
2. |
Fabaceae |
17 |
10,3 |
|
3. |
Rosaceae |
15 |
9,1 |
|
4. |
Poaceae |
14 |
8,5 |
|
8-11 |
Alliaceae |
5 |
3.0 |
|
8-11 |
Chenopodiaceae |
5 |
3.0 |
|
8-11 |
Brassicaceae |
5 |
3.0 |
|
12-15 |
Boraginaceae |
3 |
1.8 |
|
12-15 |
Crassulaceae |
3 |
1.8 |
|
12-15 |
Cyperaceae |
3 |
1.8 |
|
16-26 |
Campanulaceae |
2 |
1.2 |
|
16-26 |
Campanulaceae |
2 |
1.2 |
|
16-26 |
Convallariaceae |
2 |
1.2 |
|
16-26 |
Ephedraceae |
2 |
1.2 |
|
16-26 |
Geraniaceae |
2 |
1.2 |
|
16-26 |
Grossulariaceae |
2 |
1.2 |
|
16-26 |
Limoniaceae |
2 |
1.2 |
|
16-26 |
Polygonaceae |
2 |
1.2 |
|
16-26 |
Primulaceae |
2 |
1.2 |
|
16-26 |
Rubiaceae |
2 |
1.2 |
|
16-26 |
Selaginellaceae |
2 |
1.2 |
|
16-26 |
Urticaceae |
2 |
1.2 |
|
Всего |
165 |
К одновидовым относятся 16 семейств: Asclepiadaceae, Asparagaceae, Betulaceae, Dipsacaceae, Gentianaceae, Iridaceae, Liliaceae, Onagraceae, Pinaceae, Plantaginaceae, Polygalaceae, Rhamnaceae, Sinopteridaceae, Ulmaceae, Valerianaceae, Woodsiaceae.
Сравнение позиций основных семейств ильмовников с U. pumila Западного Забайкалья с близкими формациями ильмовников с U. pumila Восточного Забайкалья (Бутина, 2014) и абрикосников Селенгинского среднегорья (Бухарова, Намзалов, 2016) показывает их высокое сходство, особенно позиции первых семи семейств (табл. 2), однако далее возникают различия в долях участия маловидовых семейств.
Таблица 2
Соотношение основных семейств во флоре близких формаций
|
Семейства |
Ильмовники с U. pumila Западного Забайкалья |
Ильмовники с U. pumila Восточного Забайкалья (Бутина, 2014) |
Абрикосники Селенгинского среднегорья (Бухарова, Нам-залов, 2016) |
||||
|
№ |
Семейство |
% |
место |
% |
место |
% |
место |
|
1. |
Asteraceae |
16,4 |
1 |
14,9 |
1 |
14,4 |
1 |
|
2. |
Fabaceae |
10,3 |
2 |
8,1 |
4 |
11 |
2 |
|
3. |
Rosaceae |
9.1 |
3 |
13,5 |
2 |
10,7 |
3 |
|
4. |
Poaceae |
8,5 |
4 |
9,5 |
3 |
10,3 |
4 |
|
5. |
Lamiaceae |
5,5 |
5 |
4 |
8 |
3,7 |
7-8 |
|
6. |
Caryophyllaceae |
4.2 |
6 |
4,7 |
6-7 |
3,7 |
7-8 |
|
7. |
Ranunculaceae |
3,6 |
7 |
4,7 |
6–7 |
2,9 |
9 |
|
8. |
Allyaceae |
3.0 |
8-11 |
- |
- |
- |
- |
|
9. |
Apyaceae |
3.0 |
8-11 |
3,4 |
9–10 |
2,5 |
10-11 |
|
10. |
Chenopodiaceae |
3.0 |
8-11 |
3,4 |
9–10 |
- |
* |
|
11. |
Brassicaceae |
3.0 |
8-11 |
- |
- |
4 |
6 |
Родовой спектр изучаемых ильмовников более оригинален, так как всего лишь семь родов имеют в своем составе более 3 видов, из них заметно выделяется Artemisia, включающая 10 видов (табл. 3). Далее по списку 28 двувидовых родов, 74 одновидовых рода. Последние составляют, таким образом, 44% всей це-нофлоры, что свойственно флорам аллохтонным (Толмачев, 1974) и характеризует ее как гетерогенную.
Таблица 3
Ведущие семейства и роды в ценофлоре ильмовников
|
№ |
Род |
Число видов |
% |
|
1 |
Artemisia |
10 |
6.1 |
|
2 |
Allium |
5 |
3.0 |
|
3-4 |
Potentilla |
4 |
2.4 |
|
3-4 |
Thalictrum |
4 |
2.4 |
|
5-8 |
Carex |
3 |
1.8 |
|
5-8 |
Oxytropis |
3 |
1.8 |
|
5-8 |
Silene |
3 |
1.8 |
|
5-8 |
Vicia |
3 |
1.8 |
Из 8 наиболее богатых родов флоры ильмовников Западного Забайкалья 4 рода ( Artemisia, Potentilla, Allium, Oxytropis ) во многих флористических работах, посвященных флоре степей (Пешкова, 2011) традиционно входят в список ведущих, что также подтверждает степной характер изучаемой ценофлоры.
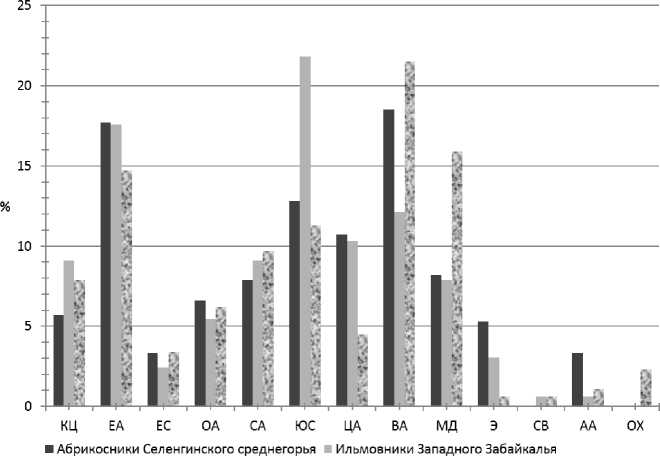
Высокую информативность при ботанико-географическом анализе флоры имеет изучение ее географической структуры. Мы сравнили группы ареалов в ценофлоре ильмовников Восточного Забайкалья (Бутина, 2009) с выделенными нами группами в ильмовниках Западного Забайкалья, а также в ценофлоре абри-косников Селенгинского среднегорья (Бухарова, Намзалов, 2016). Географическое положение, преимущественно горный рельеф изучаемого региона в современности в значительной степени влияют на ареалогическую структуру ильмовников (рис. 1). Влияние восточно-азиатского (доля участия 12,1%) и дауроманьчжурского (7,9%) центров формирования флор значительно ослабляется в сравнении с абрикосниками и ильмовниками Восточного Забайкалья, тогда как южно-сибирский центр становится определяющим (21,8%).
Сравнение этих же формаций показало достаточно близкий состав пояснозональных элементов формаций, что объясняется их общим генезисом; для них характерно сложение преимущественно степными видами (рис. 2). Широколиственно-лесное происхождение сравниваемых формаций отражается в заметном участии светлохвойных (7.3-9.5% участия) и пребореальных (2.1.-4.7%) видов.
Отличия ильмовников Западного Забайкалья состоят в высокой доле участия собственно степных видов, что объясняется более аридными условиями Западного Забайкалья наряду с широким распространением сообщества ильма низкого в Бурятии в лесостепной и степной зонах: степная группа видов включает 142 вида, что составляет 86% всей ценофлоры. Кроме того, ильмовники Западного Забайкалья имеют более широкий экологический диапазон видов, произрастающих на склонах различных экспозиций и на водоразделах, в ценофлору единично включаются монтанные, луговые, адвентивные виды.
fl Ильмовники Восточного Забайкалья
Рис. 1. Географические элементы в ценофлорах ильмовников: КЦ — циркумполярный, АА — американо-азиатский, ЕА — евразиатский, ОА — общеазиатский, СА — североазиатский, ЮС — южносибирский, ЦА — центральноазиатский, СВ — северовосточноазиатский, ВА — восточноазиатский, ЕС — евросибирский, МД — маньчжуро-даурский, Э — эндемичный, ОХ — Охотский.
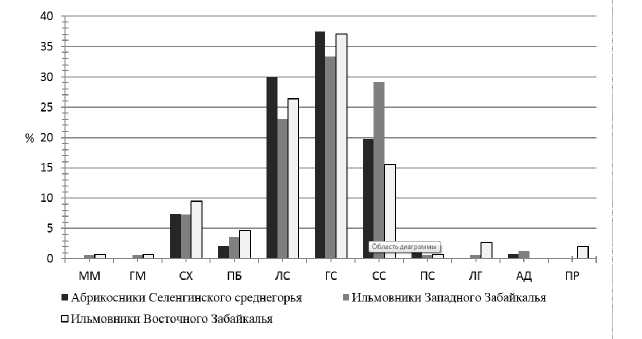
Рис. 2. Поясно-зональные элементы ценофлоры ильмовников: ММ — горная общепоясной, ГМ — гипарктомонтанный, СХ — светлохвойный, ПБ — пребореальный, ЛС — лесостепной, ГС — горностепной, СС — собственно степной, ПС — пустынностепной, ЛГ — луговой, АД — адвентивный, ПР — прибрежный.
Ильмовники Восточного Забайкалья проявляют более мезофитный характер с участием луговых и прибрежных видов, что объясняется более влажным климатом и близостью к центру исходного широколиственно-лиственного ареала. У абри-косников, имеющих строгую экотопическую приуроченность к южным склонам, экологический спектр беднее, что вполне закономерно, соответственно, здесь более высокая, чем в других формациях, доля горностепных и лесостепных видов.
Соотношение жизненных форм растений в ильмовниках отражает специфику сообществ (рис. 3). Из деревьев, кроме U. pumila , отмечены единично Betula pendula, Padus avium, Malus baccata. Pinus sylvestris имеет довольно высокую встречаемость — 22%, но обилие его невысокое: сосняки часто контактируют с ильмовниками, располагаясь в рельефе выше по склону. Из древесных форм высока доля кустарников (11 видов, составляющих около 7%), причем из них наиболее фитоцеотически значимы Spiraea aquilegifolia, Caragana pygmaea, Rhamnus erythroxylon, Cotoneaster melanocarpus, Ribes diacantha , встречаемость у этих видов более 30%. Остальные встречаются в специфических сообществах ( Armeniaca sibirica, Caragana buriatica, Rosa davurica, Amygdalus pedunculata, Ribes pulchellum ), и на опушках ( Spiraea media ) с небольшим обилием и встречаемостью.
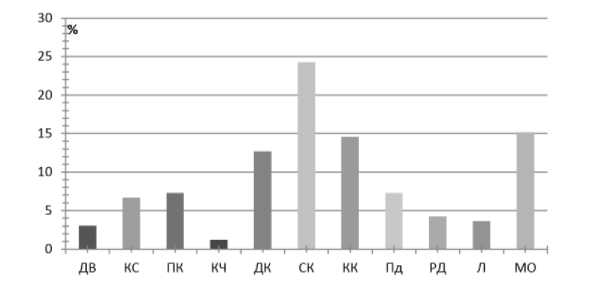
Рис. 3. Жизненные формы: ДВ — деревья, КС — кустарники, ПК — полукустарники и полукустарнички, КЧ — кустарнички и полукустарнички, ДК — длиннокорневищные, СК — стержнекорневые, КК — короткокорневищные, ПД — плотнодерновинные, РД — рыхлодерновинные, Л — луковичные и клубневые, Мо — однолетние и двулетние моно-карпики.
Ожидаемо высока доля стержнекорневых, короткокорневищных и дерновин-ных трав. Участие длиннокорневищных растений повышается в ильмовниках, произрастающих на легких песчаных и супесчаных почвах ( Leymus chinensis, Carex korshinskyi ). Интерес вызывает высокая доля монокарпических трав. Анализ их участия показал, что они чаще встречаются в сообществах, приуроченных к крутым склонам, днищам оврагов, где формируются неустойчивые экотопы, периодически нарушающиеся водным стоком или процессами сноса. Из моно-карпиков наиболее обычны Chamaerhodos erecta, Androsace septentrionalis, Dontostemon integrifolius . Большая часть монокарпических форм встречается достаточно редко, они отмечены в нескольких сообществах ( Artemisia sieversiana, Heteropappus biennis, Neopallasia pectinata, Cynoglossum divaricatum и др.).
Эффективность использования показателей активности видов во флористических исследованиях обоснована в ряде работ (Юрцев, 1968; Хитун, Зверев, 2012). Активность отражает способность той или иной группы видов господствовать в пределах флоры изучаемого ключевого участка — локальной флоры (Телятников, 2009). Различают географическую, ландшафтную и парциальную (в преде- лах того или иного типа или класса экотопов) активность (Юрцев, 1987а). В геоботанических исследованиях расчеты активности для характеристики ценофлор начали применять относительно недавно (Зверев и др., 2016).
Анализ рассчитанных в программе IBIS параметров активности видов цено-флоры U.pumila показал, что наряду с ожидаемой высокой активностью ильма также фитоценотически высокоактивен, т.е. встречается c высоким обилием, хотя более низким постоянством, кустарник Spiraea aquilegifolia (табл. 4 ). Эти данные согласуются с работой Иметхеновой О. В. (2008), где в схеме классификации сообществ спирейников выделена ильмовая формация, включающая ассоциации с доминированием спиреи водосборолистной в кустарниковом ярусе. Для Восточного Забайкалья генетические связи этих исходно неморальных видов отражены в публикациях Дулеповой Б. И. (1993) и Бутиной Н. А. (2009).
В целом, в ценофлоре U. pumila высокую активность имеют степные травянистые виды — эдификаторы и диагностические виды центральноазиатских и южносибирских настоящих и луговых степей Agropyron cristatum, Potentilla acaulis и др. При этом ряд видов характеризуется высокой встречаемостью, но сравнительно небольшой активностью ( Potentilla tanacetifolia, Goniolimon speciosum), что показывает их ассектаторные свойства в фитоценозах ильмовников.
Ценофлора Ulmus pumila L. в Западном Забайкалье
Таблица 4
|
№ |
Виды |
Встречаемость, % |
Расчетная активность |
Пояснозональная группа |
Географическая группа |
Жизненная форма |
|
1. |
Ulmus pumila |
100 |
27.9 |
СС |
ЦА |
ДВ |
|
2. |
Spiraea aquilegifolia |
70 |
27.4 |
ГС |
ЦА |
КС |
|
3. |
Potentilla acaulis |
61 |
11.7 |
ГС |
ОА |
Пд |
|
4. |
Agropyron cristatum |
70 |
11.1 |
СС |
СА |
РД |
|
5. |
Achnatherum sibiricum |
48 |
10.6 |
ЛС |
СА |
Пд |
|
6. |
Artemisia gmelinii |
52 |
9.6 |
ГС |
СА |
ПК |
|
7. |
Artemisia frigida |
61 |
9.0 |
ГС |
КЦ |
ПК |
|
8. |
Galium verum |
57 |
9.0 |
ЛС |
КЦ |
ДК |
|
9. |
Potentilla tanacetifolia |
70 |
8.8 |
ЛС |
ЮС |
СК |
|
10. |
Carex korshinskyi |
35 |
7.5 |
СС |
ВА |
ДК |
|
11. |
Poa botryoides |
22 |
7.5 |
ГС |
СА |
Пд |
|
12. |
Carex pediformis |
31 |
7.3 |
ЛС |
ЕА |
Пд |
|
13. |
Lespedeza juncea |
44 |
7.0 |
СС |
ВА |
СК |
|
14. |
Carex duriuscula |
18 |
6.5 |
СС |
АА |
ДК |
|
15. |
Cleistogenes squarrosa |
48 |
6.2 |
СС |
ЕА |
РД |
|
16. |
Koeleria cristata |
44 |
5.6 |
СС |
КЦ |
Пд |
|
17. |
Lespedeza davurica |
44 |
4.8 |
СС |
ВА |
ПК |
|
18. |
Thymus baicalensis |
22 |
4.6 |
ГС |
ГЭ |
ПК |
|
19. |
Caragana pygmaea |
39 |
4.6 |
ГС |
ЮС |
КС |
|
20. |
Festuca dahurica |
9 |
4.5 |
СС |
ВА |
Пд |
|
21. |
Caragana buriatica |
18 |
4.3 |
СС |
ГЭ |
КС |
|
22. |
Goniolimon speciosum |
52 |
4.1 |
СС |
ЕА |
СК |
|
23. |
Cotoneaster melanocarpus |
31 |
3.8 |
СХ |
ЕА |
КС |
|
24. |
Pulsatilla turczaninovii |
31 |
3.8 |
ЛС |
ОА |
СК |
|
25. |
Artemisia commutata |
31 |
3.8 |
СС |
СА |
СК |
|
26. |
Schizonepeta multifida |
31 |
3.5 |
ЛС |
МД |
СК |
|
27. |
Ribes diacantha |
31 |
3.3 |
ГС |
МД |
КС |
|
28. |
Serratula centauroides |
31 |
3.3 |
СС |
ЮС |
КК |
|
29. |
Selaginella rupestris |
13 |
3.2 |
ММ |
СВ |
КК |
|
30. |
Chamaerhodos erecta |
35 |
3.2 |
СС |
СА |
МО |
|
31. |
Rhamnus erythroxylon |
35 |
3.2 |
ГС |
ЮС |
КС |
|
32. |
Stellaria dichotoma |
39 |
3.1 |
ГС |
ЮС |
СК |
|
33. |
Sedum aizoon |
31 |
3.0 |
ГС |
СА |
КК |
|
34. |
Patrinia rupestris |
31 |
2.9 |
ГС |
ЮС |
ДК |
|
35. |
Silene repens |
22 |
2.7 |
СХ |
ОА |
ДК |
|
36. |
Iris humilis |
18 |
2.7 |
ЛС |
ЕА |
РД |
|
37. |
Pinus sylvestris |
22 |
2.7 |
СХ |
ЕА |
ДВ |
|
38. |
Phlomoides tuberosa |
35 |
2.5 |
ЛС |
ЕА |
КК |
|
39. |
Androsace septentrionalis |
26 |
2.5 |
СС |
КЦ |
МО |
|
40. |
Asparagus burjaticus |
31 |
2.5 |
ГС |
ГЭ |
КК |
|
41. |
Echinops latifolius |
18 |
2.4 |
СС |
ЮС |
ДК |
|
42. |
Rubia cordifolia |
31 |
2.4 |
ЛС |
ВА |
КК |
|
43. |
Polygonatum odoratum |
18 |
2.3 |
ЛС |
ЕА |
ДК |
|
44. |
Dianthus versicolor |
22 |
2.2 |
ГС |
ЕА |
СК |
|
45. |
Potentilla bifurca |
31 |
2.1 |
ЛС |
ЕС |
ПК |
|
46. |
Stemmacantha uniflora |
26 |
2.0 |
СС |
ВА |
СК |
|
47. |
Allium ramosum |
18 |
1.8 |
ЛС |
ОА |
Л |
|
48. |
Lilium pumilum |
22 |
1.8 |
ГС |
ВА |
Л |
|
49. |
Armeniaca sibirica |
22 |
1.8 |
ЛС |
ВА |
КС |
|
50. |
Elytrigia repens |
5 |
1.7 |
ЛС |
КЦ |
ДК |
|
51. |
Stipa krylovii |
13 |
1.6 |
СС |
ЦА |
Пд |
|
52. |
Polygala tenuifolia |
13 |
1.6 |
ГС |
МД |
ДК |
|
53. |
Cymbaria daurica |
13 |
1.6 |
ГС |
ЦА |
СК |
|
54. |
Kochia prostrata |
22 |
1.5 |
СС |
ЕА |
ПК |
|
55. |
Alyssum obovatum |
18 |
1.5 |
ГС |
ОА |
СК |
|
56. |
Crepis bungei |
18 |
1.5 |
СХ |
ЮС |
СК |
|
57. |
Orostachys spinosa |
13 |
1.5 |
ГС |
ОА |
МО |
|
58. |
Thalictrum foetidum |
22 |
1.4 |
ГС |
ЕА |
ДК |
|
59. |
Veronica incana |
18 |
1.4 |
ГС |
ЕА |
СК |
|
60. |
Hedysarum fruticosum |
9 |
1.4 |
СС |
ЦА |
ПК |
|
61. |
Dontostemon integrifolius |
18 |
1.3 |
СС |
ЮС |
МО |
|
62. |
Urtica cannabina |
18 |
1.2 |
ЛС |
СА |
КК |
|
63. |
Vincetoxicum sibiricum |
18 |
1.2 |
ГС |
ЦА |
КК |
|
64. |
Alyssum lenense |
9 |
1.2 |
ГС |
ЕА |
ПК |
|
65. |
Adenophora stenanthina |
9 |
1.2 |
ЛС |
ВА |
СК |
|
66. |
Artemisia dracunculus |
9 |
1.2 |
ЛС |
КЦ |
ДК |
|
67. |
Thermopsis lanceolata |
18 |
1.1 |
СС |
ЕА |
ДК |
|
68. |
Chenopodium album |
13 |
0.9 |
СС |
ВА |
МО |
|
69. |
Androsace incana |
13 |
0.9 |
ГС |
ЮС |
КК |
|
70. |
Thalictrum petaloideum |
9 |
0.9 |
ГС |
ВА |
СК |
|
71. |
Saposhnikovia divaricata |
9 |
0.9 |
ГС |
ВА |
СК |
|
72. |
Thymus mongolicus |
9 |
0.9 |
ГС |
ЮС |
ПК |
|
73. |
Rosa davurica |
13 |
0.8 |
СХ |
МД |
КС |
|
74. |
Artemisia scoparia |
13 |
0.8 |
ЛС |
ЕА |
МО |
|
75. |
Ephedra monosperma |
9 |
0.8 |
ГС |
ЮС |
КЧ |
|
76. |
Thalictrum squarrosum |
9 |
0.8 |
СС |
ЮС |
КК |
|
77. |
Bupleurum scorzonerifolium |
9 |
0.8 |
СС |
ВА |
СК |
|
78. |
Ephedra dahurica |
5 |
0.8 |
СС |
ЦА |
КЧ |
|
79. |
Leymus secalinus |
5 |
0.8 |
СС |
ЦА |
ДК |
|
80. |
Allium stellerianum |
5 |
0.8 |
ГС |
ЮС |
Л |
|
81. |
Pulsatilla patens |
5 |
0.8 |
ЛС |
КЦ |
КК |
|
82. |
Amygdalus pedunculata |
5 |
0.8 |
ГС |
ГЭ |
КС |
|
83. |
Chamaenerion angustifolium |
5 |
0.8 |
СХ |
КЦ |
ДК |
|
84. |
Bupleurum bicaule |
5 |
0.8 |
ГС |
МД |
СК |
|
85. |
Artemisia rutifolia |
5 |
0.8 |
ГС |
ЦА |
ПК |
|
86. |
Selaginella sanguinolenta |
9 |
0.6 |
ГС |
ЮС |
ПК |
|
87. |
Aleuritopteris argentea |
9 |
0.6 |
ГС |
ЦА |
КК |
|
88. |
Cleistogenes kitagawae |
9 |
0.6 |
ГС |
МД |
РД |
|
89. |
Setaria viridis |
9 |
0.6 |
АД |
КЦ |
РД |
|
90. |
Allium anisopodium |
9 |
0.6 |
ГС |
ВА |
Л |
|
91. |
Allium bidentatum |
9 |
0.6 |
ГС |
ЮС |
Л |
|
92. |
Allium senescens |
9 |
0.6 |
СС |
ЕА |
Л |
|
93. |
Aconogonon divaricatum |
9 |
0.6 |
ЛС |
МД |
КК |
|
94. |
Axyris hybrida |
9 |
0.6 |
СС |
ЦА |
МО |
|
95. |
Teloxys aristata |
9 |
0.6 |
СС |
КЦ |
МО |
|
96. |
Elisanthe aprica |
9 |
0.6 |
ЛС |
ЮС |
МО |
|
97. |
Camelina microcarpa |
9 |
0.6 |
ПБ |
ЮС |
МО |
|
98. |
Orostachys malacophylla |
9 |
0.6 |
ГС |
МД |
МО |
|
99. |
Spiraea media |
9 |
0.6 |
СХ |
ЕА |
КС |
|
100. |
Glycyrrhiza uralensis |
9 |
0.6 |
СС |
ОА |
КК |
|
101. |
Lupinaster pentaphyllus |
9 |
0.6 |
ЛС |
ЕА |
СК |
|
102. |
Vicia cracca |
9 |
0.6 |
ЛГ |
КЦ |
КК |
|
103. |
Amethystea caerulea |
9 |
0.6 |
ЛС |
СА |
МО |
|
104. |
Leonurus sibiricus |
9 |
0.6 |
ГС |
ЮС |
МО |
|
105. |
Campanula glomerata |
9 |
0.6 |
ЛС |
ЕА |
СК |
|
106. |
Heteropappus altaicus |
9 |
0.6 |
СС |
ЦА |
КК |
|
107. |
Scorzonera radiata |
9 |
0.6 |
СХ |
СА |
СК |
|
108. |
Chamaerhodos altaica |
5 |
0.6 |
ГС |
ЮС |
Пд |
|
109. |
Geranium sibiricum |
9 |
0.5 |
ЛС |
ЕА |
МО |
|
110. |
Dracocephalum foetidum |
9 |
0.5 |
СС |
ЦА |
МО |
|
111. |
Linaria buriatica |
9 |
0.5 |
СС |
ЮС |
СК |
|
112. |
Plantago media |
9 |
0.5 |
СХ |
ЕА |
СК |
|
113. |
Polygonatum sibiricum |
5 |
0.4 |
ГС |
МД |
КК |
|
114. |
Vicia nervata |
5 |
0.4 |
ЛС |
ЮС |
ДК |
|
115. |
Scabiosa comosa |
5 |
0.4 |
ГС |
ЮС |
СК |
|
116. |
Filifolium sibiricum |
5 |
0.4 |
ГС |
ВА |
СК |
|
117. |
Youngia tenuifolia |
5 |
0.4 |
ГС |
СА |
СК |
|
118. |
Woodsia ilvensis |
5 |
0.3 |
ГМ |
КЦ |
КК |
|
119. |
Leymus chinensis |
5 |
0.3 |
СС |
ЦА |
ДК |
|
120. |
Poa angustifolia |
5 |
0.3 |
ЛС |
ЕА |
ДК |
|
121. |
Stipa baicalensis |
5 |
0.3 |
СС |
МД |
Пд |
|
122. |
Betula pendula |
5 |
0.3 |
ПБ |
ЕС |
ДВ |
|
123. |
Urtica angustifolia |
5 |
0.3 |
ПБ |
ОА |
ДК |
|
124. |
Rheum rhabarbarum |
5 |
0.3 |
ГС |
ЮС |
КК |
|
125. |
Chenopodium acuminatum |
5 |
0.3 |
ГС |
ЦА |
МО |
|
126. |
Lychnis sibirica |
5 |
0.3 |
ГС |
ЕА |
СК |
|
127. |
Silene nutans |
5 |
0.3 |
ЛС |
ЕС |
СК |
|
128. |
Thalictrum minus |
5 |
0.3 |
СХ |
ЕА |
КК |
|
129. |
Dontostemon micranthus |
5 |
0.3 |
СС |
ЮС |
МО |
|
130. |
Ribes pulchellum |
5 |
0.3 |
ГС |
МД |
КС |
|
131. |
Fragaria orientalis |
5 |
0.3 |
ПБ |
ВА |
КК |
|
132. |
Padus avium |
5 |
0.3 |
ПБ |
ЕА |
ДВ |
|
133. |
Astragalus brevifolius |
5 |
0.3 |
СС |
ЮС |
Пд |
|
134. |
Astragalus melilotoides |
5 |
0.3 |
СС |
ЮС |
СК |
|
135. |
Oxytropis caespitosa |
5 |
0.3 |
ГС |
ГЭ |
Пд |
|
136. |
Oxytropis lanata |
5 |
0.3 |
СС |
ЮС |
КК |
|
137. |
Oxytropis oxyphylla |
5 |
0.3 |
СС |
ЮС |
РД |
|
138. |
Vicia amoena |
5 |
0.3 |
ЛС |
СА |
ДК |
|
139. |
Geranium sylvaticum |
5 |
0.3 |
ПБ |
ЕС |
КК |
|
140. |
Carum buriaticum |
5 |
0.3 |
ЛС |
МД |
СК |
|
141. |
Kitagawia baicalensis |
5 |
0.3 |
ГС |
ЮС |
СК |
|
142. |
Ciminalis squarrosa |
5 |
0.3 |
СС |
ОА |
МО |
|
143. |
Lappula squarrosa |
5 |
0.3 |
ГС |
КЦ |
МО |
|
144. |
Dracocephalum nutans |
5 |
0.3 |
ЛС |
ЕА |
СК |
|
145. |
Lophanthus chinensis |
5 |
0.3 |
ГС |
МД |
СК |
|
146. |
Achillea asiatica |
5 |
0.3 |
ЛС |
СА |
ДК |
|
147. |
Artemisia anethifolia |
5 |
0.3 |
СС |
ЮС |
МО |
|
148. |
Artemisia sericea |
5 |
0.3 |
ЛС |
ЕА |
ПК |
|
149. |
Artemisia sieversiana |
5 |
0.3 |
АД |
СА |
МО |
|
150. |
Artemisia tanacetifolia |
5 |
0.3 |
СХ |
ЮС |
ДК |
|
151. |
Heteropappus biennis |
5 |
0.3 |
СС |
ЦА |
МО |
|
152. |
Ixeris graminea |
5 |
0.3 |
СС |
ВА |
СК |
|
153. |
Leibnitzia anandria |
5 |
0.3 |
ЛС |
ВА |
КК |
|
154. |
Neopallasia pectinata |
5 |
0.3 |
ПС |
ЦА |
МО |
|
155. |
Saussurea salicifolia |
5 |
0.3 |
ГС |
ЮС |
СК |
|
156. |
Scorzonera austriaca |
5 |
0.3 |
ЛС |
ЕА |
СК |
|
157. |
Taraxacum dissectum |
5 |
0.3 |
СС |
ЮС |
СК |
|
158. |
Taraxacum officinale |
5 |
0.3 |
СХ |
КЦ |
СК |
|
159. |
Silene jeniseensis |
5 |
0.2 |
ГС |
ЮС |
Пд |
|
160. |
Malus baccata |
5 |
0.2 |
ЛС |
ВА |
ДВ |
|
161. |
Potentilla multifida |
5 |
0.2 |
ГС |
КЦ |
РД |
|
162. |
Medicago falcata |
5 |
0.2 |
ЛС |
ЕА |
СК |
|
163. |
Limonium flexuosum |
5 |
0.2 |
СС |
ЮС |
СК |
|
164. |
Cynoglossum divaricatum |
5 |
0.2 |
ЛС |
ВА |
МО |
|
165. |
Lappula redowskii |
5 |
0.2 |
СС |
СА |
МО |
Заключение. Проведенный анализ ценофлоры U. pumila в Западном Забайка- лье позволяет сделать следующие выводы.
Сравнительная характеристика ценофлоры ильмовников Западного и Восточного Забайкалья показала определяющую роль влияния южно-сибирского горностепного центра формирования флор и значительное ослабление восточноазиатского и дауро-маньчжурского центров в сравнении с абрикосниками и ильмовниками Восточного Забайкалья, что обусловлено географическим положением, рельефом и палеоклиматической историей региона.
Систематический и экологический состав ценофлоры ильмовников характерен для степной флоры Сибири и указывает на степной генезис большей части видового состава, тогда как эдификатор ильм низкий относится к реликтам разреженных широколиственных лесов центральной и восточной Азии. Данный факт позволяет предположить чрезвычайно высокую степень адаптации U. pumila к засушливым условиям региона, а также успешность вхождения этого древесного вида в степные флороценотипы и фитоценотическую коадаптацию в формируемых сообществах в процессе эволюции.
Исследования проведены при поддержке гранта БГУ № 18-14-0502
Список литературы Характеристика ценофлоры Ulmus pumila L. в Западном Забайкалье
- Бутина Н. А. Ильмовники Восточного Забайкалья: анализ флористического и фитоце-нотического разнообразия, биоэкологические особенности видов рода Ulmus L.: автореферат дис.... канд. биол. наук: 03.00.05. Улан-Удэ, 2009. 23 с
- Бутина Н. А. Систематический анализ флоры ильмовников Восточного Забайкалья // Ученые записки ЗабГУ. Серия: Естественные науки. 2014. № 1 (54). С. 13-17
- Бухарова Е. В. Намзалов Б. Б. Абрикосники Западного Забайкалья. Улан-Удэ: Изд-во БГУ, 2016. 146 с
- Дулепова Б. И. Степи горной лесостепи Даурии и их динамика. Чита: Изд-во ЧГПИ, 1993. 396 с.
- Зверев А. А. Информационные технологии в исследованиях растительного покрова. Томск: ТМЛ-Пресс, 2007. 304 с
